Original Article
Elevated expression of cyclophilin A
in human periodontitis
Li Zhao
1,2, Xijiao Yu
3, Lu Zhao
1,2, Fenghe Zhang
11Department of Oral and Maxillofacial Surgery, School of Stomatology Shandong University, Shandong Provincial Key Laboratory of Oral Tissue Regeneration, Jinan 250012, Shandong, PR China; 2Department of Periodontology, Binzhou People’s Hospital, Binzhou, People’s Republic of China; 3Department of Endodontics, Jinan Stomatologi-cal Hospital, Jinan, Shandong, China
Received October 6, 2017; Accepted February 2, 2018; Epub March 15, 2018; Published March 30, 2018
Abstract: This study elucidated the correlation between cyclophilin A (CypA) and inflammatory infiltrating cells in human periodontitis. Western blotting, immunohistochemistry, and immunofluorescence were performed to detect the expression level of CypA in gingival tissues of human periodontitis. Healthy gingival tissues were chosen as a control. The distribution of CD3+, CD4+, CD22+, and CD68+ infiltrating cells in the gingival tissues were also detected
by immunohistochemistry. Western blotting analysis and immunohistochemistry revealed significantly increased ex-pression of CypA in human periodontitis. NF-κB p-p65 and p-IκBα exex-pression was detected to investigate NF-κB acti-vation. Immunohistochemistry and immunofluorescence identified that the positive-expressed CypA was localized in the infiltrating cells. CD3+, CD4+, CD22+, and CD68+ cells all could be observed in the CypA-positive infiltrating cells.
The NF-κB pathway was activated in human periodontitis. In conclusion, CypA is involved in leukocyte attraction in the periodontal inflammatory response. These efforts not only highlight the pathogenesis of human periodontitis, but also signify CypA as a potential target for anti-inflammatory therapeutics.
Keywords: Cyclophilin A, periodontitis, inflammatory cells
Introduction
Cyclophilin A (CypA), a member of the
immu-nophilin family, has peptidyl prolylcis-trans
isomerase (PPIase) activity, which regulates
immune-modulation, protein folding, trafficking
assembly, and cell signaling [1, 2]. CypA is a
highly conserved protein that is expressed in
a wide range of tissues. Evidence supporting
important functions for CypA in rheumatoid
arthritis [3, 4], cancer [5], cardiovascular
dis-eases [6], sepsis [7], periodontitis [8, 9] and
aging [10], are gradually increasing. Recent
research shows that CypA can be secreted by
the infiltrating cells in response to inflammatory
stimuli [11, 12]. CypA is a potent chemokine,
which can directly induce leukocyte chemotaxis
and contribute to the pathogenesis of
inflam-mation-mediated diseases [13]. We presumed
that elevated CypA induces more infiltrating
cells to diseased sites, and then the infiltrating
cells further secrete CypA to aggravate
peri-odontal inflammatory destruction. The
correla-tion of CypA and inflammatory infiltrating cells
in human periodontitis not only elucidates the
pathogenesis of human periodontitis, but also
signifies CypA as a target for anti-inflammatory
therapeutics [14]. Therefore, this work aimed
to elucidate the distribution of CypA and its
cor-relation with inflammatory infiltrating cells in
human periodontitis.
Material and methods
Study participants and sample collection
during orthodontic extractions from 10 heal-
thy donors without systematic diseases. Each
specimen was divided into two parts of
approxi-mately equal size. One part was immediately
fixed in 4% paraformaldehyde and then 5 μm
serial sections were made for
immunohisto-chemistry and immunofluorescence analyses.
The other part was stored in liquid nitrogen for
Western blotting.
Immunohistochemistry
Immunohistochemical study was performed by
[image:2.612.94.376.71.260.2]protein quantitative analysis assay kit (BOSHE,
China); Proteins were separated on 10% SDS
gels and then transferred onto polyvinylidene
difluoride membranes (Millipore, USA). After
blocking in 0.1% Tween 20 in Tris-buffered
saline (TBST) containing 5% nonfat dried milk
for 1 h at room temperature, the membranes
were incubated with antibodies against CypA
(diluted 1:1000, Abcam), NF-κB p-p65 (1:1000,
Cell Signaling Technology, USA) or p-IκBα
(1:1000, Cell Signaling Technology, USA)
over-night at 4°C. Before incubation with
horserad-ish peroxidase (HRP)-labeled second antibody
Figure 1. Expression and distribution of CypA in human inflamed and healthygingiva. The positive staining (red arrows) was distributed in the gingival epi-thelium of the healthy gingiva. The positive staining (red arrows) was princi-pally distributed in the infiltrating cells in human periodontitis.
Beijing, China) as previously
described [15]. Polyclonal an-
tibody CypA (dilution 1:100,
Abcam, UK), CD3 (dilution
1:100, Abcam, UK), CD4
(dilu-tion 1:100, Abcam, UK), CD22
(dilution 1:100, Abcam, UK),
and CD68 (dilution 1:100, Ab-
cam, UK) were applied. PBS
was obtained as control.
Immunofluorescence
Sections were deparaffinized
in xylene and rehydrated. After
washing, the sections were in-
cubated with polyclonal
anti-body CypA (dilution 1:100, Ab-
cam, UK) at 4°C overnight.
The sections were then in-
cubated with rhodamine (TRI-
TC)-conjugated goat
anti-rab-bit IgG (Sigma, USA) for 1 h
at room temperature. Nucl-
ei were stained with DAPI
solution (Sigma, USA) for 5
min. The sections were
photo-graphed with
immunofluores-cence microscopy (OLYMPUS
BX-60, Japan).
Western blotting analysis
Samples were washed by cold
PBS three times respectively,
and then homogenized in RIPA
buffer for 30 min. Phenylme-
thane sulfonyl-fluoride (1 mM)
was added into the buffer in
advance. Protein
concentra-tions were measured using a
bicinchoninic acid assay (BCA)
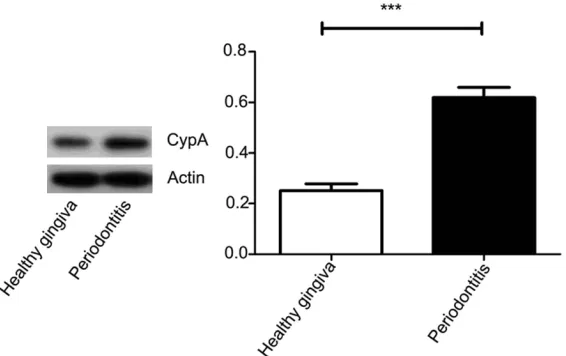
Figure 2. Expression of CypA protein in human healthy and inflamed gingiva. [image:2.612.93.376.330.508.2]TBST for 10 min 3 times. The bands were
visi-ble on the Canon films using ECL substrate
solution (Millipore). Actin (1:10000) was used
as internal control.
Statistical analysis
All of the images of Western blots assays were
representative of at least three independent
experiments. All measurement results are
pre-sented as the mean ± SD. All values were
CD68
+in the CypA-positive infiltrating cells in
human inflamed gingiva
[image:3.612.90.376.73.178.2]CD3
+, CD4
+, CD22
+, and CD68
+cells could all
be observed in the CypA-positive infiltrating
cells in the inflamed gingival tissues. CD4
+(T-helper/inducer) cells, CD68
+cells
(macro-phages), and CD3
+cells were predominant in
the lamina propria and the bottom parts of
the gingival epithelium. CD22
+B cells were
arranged in clusters in the lamina propria
Figure 3. Histological observation of infiltrating cells in human inflamedgin-giva. Many inflammatory infiltrating cells (red arrows) could be observed in the inflamed gingival of human periodontitis.
calculated using Student’s
t
-
test. The differences were co-
nsidered to be statistically
sig-nificant at
P
< 0.05.
Results
Expression of CypA in human
gingival tissues
Expression and distribution
of CypA was detected in both
the periondontitis and heal-
thy groups. Expression of Cy-
pA in the periondontitis group
was higher than that of the
healthy group. Positive
expres-sion was localized in the
gingi-val epithelium of the healthy
gingiva. In human
periodonti-tis, positive staining was
prin-cipally distributed in the
infil-trating cells, besides gingival
epithelium (
Figure 1
). Western
blotting results also showed
that expression of CypA in
inflamed gingiva was higher
than that of healthy donors (
P
< 0.05,
Figure 2
). According to
histological observation, many
inflammatory infiltrating cells
could be observed in the in-
flamed gingival of human pe-
riodontitis (
Figure 3
). Expres-
sion of CypA in the
inflamma-tory infiltrating cells was also
observed by
immunofluores-cence. Positive expression of
CypA mainly exists in the
cyto-plasm of the inflammatory in-
filtrating cells (
Figure 4
).
Expression and distribution
of CD3
+, CD4
+, CD22
+, and
[image:3.612.94.376.237.530.2]Figure 5. Expression and distribution of CD3+, CD4+, CD22+, and CD68+
in-flammatory infiltrating cells in human periodontitis. CD3+, CD4+, CD22+, and
CD68+ infiltrating cells all could be observed in the inflamed gingival tissues.
Figure 6. Quantitation of CD3+, CD4+, CD22+, and CD68+ in the
inflamma-tory infiltrating cells in human periodontitis. Positive cells were quantified by imaging five fields of view under 100-fold magnification and directly counting the number of CD3+, CD4+, CD22+ and CD68+ positive cells.
CD3+, CD4+, CD22+, and CD68+ positive cells all could be observed in the
(
Figure 5
). Positive cells were
quantified by imaging five fi-
elds of view under 100-fold
magnification and directly co-
unting the number of CD3
+,
CD4
+, CD22
+, and CD68
+posi-tive cells. CD3
+cells (T cells)
comprised the major
popula-tion of lymphocytes (
Figure 6
).
NF-κB activation
Western blotting results show
NF-κB 65 and IκBα
phosphor-ylation was upregulated in the
inflamed gingiva to higher
lev-els than in the healthy gingiva.
Therefore, the NF-κB pathway
was activated in human
peri-odontitis (
Figure 7
).
Discussion
CypA is believed to have
criti-cal roles in regulating
inflam-matory responses,
immune-modulation, trafficking asse-
mbly and MMPs production,
and it can be secreted in re-
sponse to inflammatory sti-
muli such as hypoxia,
infec-tion, and oxidative stress
[16-18]. CypA is associated with
inflammatory infiltration and
alveolar bone destruction in
rat experimental periodontitis.
CypA can induce migration of
monocyte/macrophages,
lym-phocytes, and neutrophils into
tissues [19], and contributes
to inflammatory responses th-
rough its chemotactic activity
[20-22].
[image:4.612.92.365.426.666.2]Periodontitis affects a great number of patients
all over the world, which leads to gingival
inflam-mation, alveolar bone loss, and even loose
teeth [23]. Different subsets of leukocytes
such as monocyte/macrophages, lymphocytes,
and neutrophils, are involved in the
histopatho-genesis of human periodontitis [24, 25]. CypA
levels increase in periodontitis, but cell types
expressing CypA and the function of CypA
in the pathogenesis of periodontitis are not
known yet. This study further elucidates the
correlation of CypA and the inflammatory
infil-trating cells in human periodontitis.
Inflammatory infiltrating cells are a key step in
the inflammatory response. A large amount of
inflammatory infiltrating cells could be observed
in the inflamed gingival of human periodontitis
[26].
T
he inflammatory infiltrating cells mig-
rate from blood vessels into sites of inflam-
mation, and are recognized as macrophages,
lymphocytes, and neutrophils according to th-
eir shape, size, and location [27-30]. In this
study, we found significant positive expression
of CD3
+(T cells), CD4
+(T helper cells), CD22
+(B
cells), and CD68
+(macrophages) in the
infiltra-tion of the human periodontitis. CD3
+, CD4
+,
CD22
+, and CD68
+cells all could be observed
in the CypA-positive infiltrating cells in human
periodontitis. Elevated CypA induced more
infil-trating cells to the diseased sites, and then
the infiltrating cells further secrete CypA to
aggravate periodontal inflammatory
destruc-tion, and promote NF-κB pathway activation.
The results showed that CypA could be
target-ed for anti-inflammatory therapeutics.
Although different subsets of leukocytes are
recruited during the inflammatory response
[31-33], synergism of CypA and different
sub-sets of the infiltrating cells in human
periodon-titis still remains to be determined.
Acknowledgements
This work was supported by Natural Science
Foundation of Shandong Province (ZR2017-
QH007) and Science Development Program
Project of Jinan (201121043) awarded to Xijiao
Yu.
Disclosure of conflict of interest
None.
Address correspondence to: Fenghe Zhang, Depart- ment of Oral and Maxillofacial Surgery, School of Stomatology Shandong University, Shandong Pro- vincial Key Laboratory of Oral Tissue Regeneration, 44-1 West Wenhua Road, Jinan 250012, Shandong, PR China. Tel: +86 531 88382504; E-mail: zfengh@ sdu.edu.cn
References
[1] Hou X, Liu R, Huang C, Jiang L, Zhou Y and Chen Q. Cyclophilin A was revealed as a candi-date marker for human oral submucous fibro-sis by proteomic analyfibro-sis. Cancer Biomark 2017; 20: 345-356.
[image:5.612.94.523.73.237.2][2] Piechota-Polanczyk A, Wlodarczyk M, Sobo-lewska-Wlodarczyk A, Jonakowski M, Pilarczyk A, Stec-Michalska K, Wisniewska-Jarosinska M and Fichna J. Serum cyclophilin A correlates with increased tissue MMP-9 in patients with
ulcerative colitis, but not with Crohn’s disease. Dig Dis Sci 2017; 62: 1511-1517.
[3] Wang L, Wang CH, Jia JF, Ma XK, Li Y, Zhu HB, Tang H, Chen ZN and Zhu P. Contribution of cyclophilin A to the regulation of inflammatory processes in rheumatoid arthritis. J Clin Immu-nol 2010; 30: 24-33.
[4] Lv M, Miao J, Zhao P, Luo X, Han Q, Wu Z, Zhang K and Zhu P. CD147-mediated chemo-taxis of CD4+CD161+ T cells may contribute to local inflammation in rheumatoid arthritis. Clin Rheumatol 2018; 37: 59-66.
[5] Howard BA, Furumai R, Campa MJ, Rabbani ZN, Vujaskovic Z, Wang XF, Patz EF Jr. Stable RNA interference-mediated suppression of cy-clophilin A diminishes non-small-cell lung tu-mor growth in vivo. Cancer Res 2005; 65: 8853-8860.
[6] Xue C, Sowden M and Berk BC. Extracellular cyclophilin A, especially acetylated, causes pulmonary hypertension by stimulating endo-thelial apoptosis, redox stress, and inflamma-tion. Arterioscler Thromb Vasc Biol 2017; 37: 1138-1146.
[7] Song T, Yang M, Chen J, Huang L, Yin H, He T, Huang H and Hu X. Prognosis of sepsis in-duced by cecal ligation and puncture in mice improved by anti-Clonorchis Sinensis cyclopho-lin a antibodies. Parasit Vectors 2015; 8: 502. [8] de Oliveira Nobrega FJ, de Oliveira D, Vascon-celos RG, Nonaka CFW and Queiroz LMG. Study of the participation of MMP-7, EMMPRIN and cyclophilin A in the pathogenesis of peri-odontal disease. Arch Oral Biol 2016; 72: 172-178.
[9] Liu L, Li C, Cai C, Xiang J and Cao Z. Cyclophilin A (CypA) is associated with the inflammatory infiltration and alveolar bone destruction in an experimental periodontitis. Biochem Biophys Res Commun 2010; 391: 1000-1006. [10] Nigro P, Pompilio G and Capogrossi MC.
Cy-clophilin A: a key player for human disease. Cell Death Dis 2013; 4: e888.
[11] Hoffmann H and Schiene-Fischer C. Functional aspects of extracellular cyclophilins. Biol Chem 2014; 395: 721-735.
[12] Bukrinsky M. Extracellular cyclophilins in heal- th and disease. Biochim Biophys Acta 2015; 1850: 2087-2095.
[13] Arora K, Gwinn WM, Bower MA, Watson A, Ok-wumabua I, MacDonald HR, Bukrinsky MI and Constant SL. Extracellular cyclophilins contrib-ute to the regulation of inflammatory respons-es. J Immunol 2005; 175: 517-522.
[14] Yurchenko V, Constant S, Eisenmesser E and Bukrinsky M. Cyclophilin-CD147 interactions: a new target for anti-inflammatory therapeutics. Clin Exp Immunol 2010; 160: 305-317. [15] Yu X, Lv L, Zhang J, Zhang T, Xiao C and Li S.
Expression of neuropeptides and bone
remod-eling-related factors during periodontal tissue regeneration in denervated rats. J Mol Histol 2015; 46: 195-203.
[16] Suzuki J, Jin ZG, Meoli DF, Matoba T and Berk BC. Cyclophilin A is secreted by a vesicular pathway in vascular smooth muscle cells. Circ Res 2006; 98: 811-817.
[17] Seko Y, Fujimura T, Taka H, Mineki R, Muraya-ma K and Nagai R. Hypoxia followed by reoxy-genation induces secretion of cyclophilin A from cultured rat cardiac myocytes. Biochem Biophys Res Commun 2004; 317: 162-168. [18] Jin ZG, Melaragno MG, Liao DF, Yan C,
Haendel-er J, Suh YA, Lambeth JD and BHaendel-erk BC. Cy-clophilin A is a secreted growth factor induced by oxidative stress. Circ Res 2000; 87: 789-796.
[19] Ren Y, Yang H, Zhu P, Fan CM, Wang YH, Li J and Liu H. [Effect of CsA bleomycinduced in-terstitial pulmonary disease in mice]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2012; 28: 232-236. [20] Sherry B, Yarlett N, Strupp A and Cerami A.
Identification of cyclophilin as a proinflamma-tory secreproinflamma-tory product of lipopolysaccharide-activated macrophages. Proc Natl Acad Sci U S A 1992; 89: 3511-3515.
[21] Dawar FU, Tu J, Xiong Y, Lan J, Dong XX, Liu X, Khattak MN, Mei J and Lin L. Chemotactic ac-tivity of cyclophilin A in the skin mucus of yel-low catfish (Pelteobagrus fulvidraco) and its active site for chemotaxis. Int J Mol Sci 2016; 17.
[22] Takanashi S, Nochi T, Abe M, Itaya N, Urakawa M, Sato K, Zhuang T, Umemura S, Hayashi T, Kiku Y, Kitazawa H, Rose MT, Watanabe K and Aso H. Extracellular cyclophilin A possesses chemotaxic activity in cattle. Vet Res 2015; 46: 80.
[23] Yu X, Gong Z, Lin Q, Wang W, Liu S and Li S. Denervation effectively aggravates rat experi-mental periodontitis. J Periodontal Res 2017; 52: 1011-1020.
[24] Gonzales JR. T- and B-cell subsets in periodon-titis. Periodontol 2000 2015; 69: 181-200. [25] Thorbert-Mros S, Larsson L and Berglundh T.
Cellular composition of long-standing gingivitis and periodontitis lesions. J Periodontal Res 2015; 50: 535-543.
[26] Liu L, Li C, Xiang J, Dong W and Cao Z. Over-expression and potential role of cyclophilin A in human periodontitis. J Periodontal Res 2013; 48: 615-622.
[27] Zhou GX and Liu ZJ. Potential roles of neutro-phils in regulating intestinal mucosal inflam-mation of inflammatory bowel disease. J Dig Dis 2017; 18: 495-503.
and Tacke F. Therapeutic Inhibition of Inflam-matory Monocyte Recruitment Reduces Ste-atohepatitis and Liver Fibrosis. 2017.
[29] Tacke F. Targeting hepatic macrophages to treat liver diseases. J Hepatol 2017; 66: 1300-1312.
[30] Chandrashekara S, Mukhtar Ahmad M, Renu-ka P, Anupama KR, RenuRenu-ka K. Characterization of neutrophil-to-lymphocyte ratio as a measure of inflammation in rheumatoid arthritis. Int J Rheum Dis 2017; 20: 1457-1467.
[31] Pawlik I, Mackiewicz U, Lacki JK, Wiktorowicz K and Konys J. The differences in the expression of CD45 isoforms on peripheral blood lym- phocytes derived from patients with seasonal or perennial atopic allergy. Tohoku J Exp Med 1997; 182: 1-8.
[32] Madej JA, Madej JP, Dzimira S and Nowak M. An immunohistochemical analysis of lympho-cytic infiltrations in canine skin cancers. Pol J Vet Sci 2017; 20: 141-147.