Variability in Ribulose-1,5-Bisphosphate Carboxylase/
Oxygenase Small Subunits and Carboxylation Activity in Fern Gametophytes Grown under Different Light Spectra
1Haviva Eilenberg, Sven Beer, Shimon Gepstein, Nurit Geva, Orly Tadmor, and Aviah Zilberstein*
Department of Botany, Tel Aviv University, RamatAviv, 69978 TelAviv, Israel (H.E., S.B., N.G., 0.T., A.Z.); and Department of Biology, Technion, Haifa32000, Israel (S.G.)
ABSTRACT
Two distinct ribulose-1,5-bisphosphate carboxylase/oxygen- ase(Rubisco)small subunit (SSU) populations were observed in Pteris vittata gametophytes grown under different illumination conditions. Exposure of the fem gametophytes to continuous red light (R) resulted in Rubisco SSUs that were not recognized by polyclonal antibodies raised against SSUs from spinach. Unlike the R-inducedSSUs, blue light (B) induced SSUs were well rec- ognized. This difference in SSU composition also reflected in Rubisco activity. In vitro, B-induced Rubisco exhibits a signifi- cantly higher carboxylation activity as compared to the R-induced Rubisco. Approximately atwo-tothreefold increase in theVm,x value of theB-induced carboxylase as compared to the R-induced one wasmeasured. It thus seems very likely that certain domains inthe SSU molecule affect enzyme activity.
Rubisco2 (EC 4.1.1.39) isanabundantenzymein all pho- tosyntheticorganisms. It catalyses both photosynthetic CO2 fixation via the carboxylation of RuBP in the Calvin cycle and oxygenation of the same substrate in photorespiration (3). In C3 plants, which lack a CO2 concentrating system, photosynthesis doesnotreach maximalefficiency duetothe relatively low ratio ofCO2/02 in the atmosphere. Possible modification of theholoenzymestructure, leadingtoa pref- erential increaseincarboxylation activity,could thus increase plant growth.
The Rubiscoholoenzyme iscomposedofeight chloroplast- encoded LSUs and eight nuclear-encoded SSUs. The rbcL, encodingtheLSU, ispresentas asinglecopy perchloroplast
genome. Due to reiteration ofgenomes perchloroplast and chloroplastspercell, therearethousands ofcopiesofrbcLin everyleaf cell (18). Thesegenesare posttranslationally pho- toregulatedandveryconservedthroughoutevolution(8, 16).
Only small differences have been found in the amino acid sequenceof the LSUs from differentplants (3).
'Researchsupported bythe IsraeliMinistryof ScienceandTech- nology, National Council for Research and Development, under Grant 2694-1-87.
2Abbreviations: Rubisco, ribulose-1,5-bisphosphate carboxylase/
oxygenase; RuBP, ribulose- 1,5-bisphosphate; LSU, large subunit;
SSU,smallsubunit; R, redlight; B,bluelight; W,whitelight;rbcL andrbcS,genesencodingLSUsand SSUsrespectively;AC02,activator C02-
Incontrast tothe LSUs,theRubiscoSSUs are encoded by anucleargene-family, whosemembers(rbcSs)are usually all expressed (7, 28). However, individual gene members can showmaximalexpression in differentorgansandatdifferent developmentalstages(7, 14). Although thereis divergence in the nucleotide sequence ofrbcS coding regions, the amino acid composition of the various mature subunits is very conserved and only slight differences amongst the mature proteins have been identified (7). Transcription of rbcS is light-regulatedthrough phytochromeand ablue-lightphoto- receptor, which act in concert (12, 14, 25). This regulation has been shown to be linked to light-responsive elements located upstream to the consensus sequences of the rbcS promoter(12).
Plant Rubisco requiresthe presenceofboth the LSUs and theSSUs forcatalysis (1). The activation site ofthe enzyme by CO2 and Mg2+, as well as the RuBP binding site, are located on the LSU, which also determines the degree of partitioning between carboxylation and oxygenation (2).
Thereis relatively littleknown about the SSU role.However, based on crystallography studies of L2 and L8S8 Rubisco, Schneideretal. (24) recently suggestedthat the SSU modu- lates substratebindingtothe LSUbyintroducing conforma- tional changesinLSUactive site.
Thepresentworkfurthersupportsthe ideathat SSUsare functionally involved in governingcarboxylation activity of Rubisco. Usingauniquesystemoffern gametophytesgrown under blue or red illumination it was possibleto show the occurrence oftwo distinct light-induced SSU populations.
Thediversityin SSUswasfoundtobecorrelated withdifferent catalyticpropertiesof Rubiscoas acarboxylase.
MATERIALS AND METHODS Materials
['4C]NaHCO3(50mCi/mmol)and[125I]donkeyanti-rabbit IgGwerepurchased from the RadiochemicalCenter, Amer- sham. SpinachRuBPcarboxylase and mol wtmarkerswere
purchased from Sigma. All chemicalswereofanalytical grade.
Germinationand Illumination of Ferns
FreshsporesofPteris vittata L. werecollected inthe Botan- ical Gardens of Tel-Aviv University, sterilized, and grown axenicallyasdescribedpreviously (30).Following48 himbi- 298
VARIABILITY IN RUBISCO SMALL SUBUNITS AND C02 FIXATION
bitionin the dark,synchronizedgerminationwasinduced by exposing the cultures for 2 h to red light. The spores were keptforanadditional48 h in darkness and thentransferred to continuous R (600-660 nm), B (400-550 nm), or W as described before (30). All cultures were illuminated at an equalenergyof1 Wm-2.
Anti-LSU andAnti-SSU Antibodies
Preparation ofantibodies was carried out essentially ac- cording to Zemel and Gepstein (29). Spinach Rubisco sub- units were separated on SDS-PAGE gels and the LSU and SSU bandswereeluted. Rabbit anti-LSU and anti-SSU pol- yclonal antibodieswerepreparedaccordingtoNelson(23).
Preparation ofGametophyte Extracts
Axenicallygrownfern gametophyteswerecollected,frozen andground inliquidN2, and thengroundandhomogenized inafreshlypreparedbuffercontaining200mmTris-HCl(pH 8.0), 0.2 mm EDTA, 10 mM MgCl2, 10 mM ascorbate, 2%
PVP(Mr 360,000), and 5 mm DTTat4°C. Since Rubisco is astromal enzyme, its relative concentration ingametophyte extracts could be increased by using an extraction buffer devoid ofdetergent, thusavoiding the apparent destruction ofthylacoidmembranes.Following centrifugationat12,000g for 1 min thesupernatant ("Rubiscoenriched" extract)was collected and usedfor the differentmeasurements.Extraction oftotalproteinswasdoneasaboveexceptthat thehomoge- nizingbuffer contained 60 mM Tris-HCl(pH6.8), 3% SDS, 10% glycerol and 0.7Mfl-mercaptoethanol.
Evaluation of RubiscoContentin Extracts
Total protein content asdetermined accordingto Marder etal. (21) was found to varybetween 2 and 7 mgprotein/
mL. Evaluation of Rubisco amounts in each extract was
performedby dotblotting ofincreasingamounts of Rubisco extracts (upto3 ,ug)and thenprobing firstwith rabbit anti- LSU and then with '251-donkey anti-rabbit IgG. Following overnightexposuretoRP-2(Agfa) film,the dotswereexcised from the nitrocellulose sheet and counted. Purified spinach Rubisco protein was used as standard. Linear correlation between Rubisco concentration and 1251-counting enabled accurateestimation of Rubiscocontentin theenzyme assays.
Separation ofProteins byPolyacrylamide Gels
The total proteinpattern was analyzed on SDS-polyacryl- amidegradient gels (5-15% or 10-20%) (4). Rubisco holo- enzyme wasfirst separatedon5 to25%nondenaturing poly- acrylamide gels. Theholoenzymebandwas eluted in 25 mM Tris,0.2 Mglycine, 12.5% glycerol, and 2 mm PMSF, at 4°C overnight, and then electrophoresedon SDS-PAGE. Purified spinach Rubisco and/or proteins of known mol wt were includedasmarkers.
RubiscoCarboxylation Activity
Rubiscocarboxylaseactivity was assayed in closed vials, at pH 8.0and25°C,bymeasuringRuBP-dependent incorpora-
tion of"'CO2intoacid-stable products. The totalassay volume was 0.55 mL andcontained0.3 mL assaybuffer(50mM Tris- HCI [pH 8.0], 1 mm EDTA, 10 mmMgCl2,and 5 mm DTT), 0.1 mL activated enzyme extract and various amounts of ["'C]NaHCO3 or RuBP togive final concentrations asindi- cated in the legends ofFigures 6 and7.FullRubisco activation wasachievedbypreincubatingthe enzyme extract at 4°Cfor 10 min with 10 mM Mg2+ (present in the extraction buffer) and 10 mm NaHCO3. Catalysiswas initiated by addition of the activated enzyme to the reaction mixture. The reaction wasterminatedafter0.5minof incubationby theadditionof 0.1 mL 6 NHCI. Sampleswere driedfor 2 h at60°C under anair stream, and then countedforradioactivity in Hydrol- uma.Exact[CO2] of theassaysolutionswerecalculated, given the total[NaHCO3] injected (including that added withreac- tants and buffers as well as CO2 present in the gas phase before closingthe vials, altogether0.46 ,mol), asthe distri- bution of various carbon forms in the assay mixtureat the given pH (=0.7%C02), temperatureandionicstrength, and taking into account the distribution of CO2 between the aqueousandgasphase of the closed vial (the molar fraction- ation ratio being0.88). Preexperiments had shown thatCO2 equilibria with thegasphasewerecompleted well within the activationperiod.
RESULTS
Growth ofGametophytesunder BlueorRed Illumination The free-living haploid gametophytes of the fern Pteris vittata are easy tomanipulate and their growth can becon- trolled andsynchronized byred and bluelight (30). Figure 1 shows the differentpattern ofdevelopment ofP. vittata ga- metophytes as aresultofgrowth undercontinuous R, B, or W. The Rgametophytes are filamentous whilethe B game- tophytes exhibit the usual two-dimensional growth pattern similarto thatobserved underW. Furthermore, the chloro- plastsof the Rgametophytesare mainly concentratedatthe tip ofthe filament around the nucleus while in B and W gametophytes thereis a relativelyevendistribution of chlo- roplasts in the cells.
Protein PatternsinR, B, and WGametophytes
The morphological differences observed in gametophytes grownunderdifferent lightspectraled us to question whether any alterations in total protein pattern between these three groupscould alsobe detected. Crude extracts prepared from eachilluminationgroup were electrophoresed on a 10 to 20%
SDS-polyacrylamide gel (Fig. 2). No significant differences were detected inthepattern of protein distribution in any of theextracts. Similarly, no apparentdifference in the relative intensities of the bands could be observed. But when fern Rubisco subunits were compared to spinach Rubisco sub- units,thefern SSU is 1.3kDsmaller than that of the spinach.
Interestingly, incyanobacterial SSUs a 12 amino acid portion ismissing (3)and thisaccountsfora peptide that is 1.4 kD smaller than thehigherplantSSUs.
299
A
%'
I
% 4
w.4
'4.
\ t
4..
B
C10
Cw I
C D
c1..
,
-y
I II
E
'4'
'V
.4
... ...
.,
.. k
.1
#.:-3
F
w .
50pm
h--
V.'I.W
.A,
-II..lk
I1) i, u,. / '.
..4
.-rF-:
".--el"
..r,
VARIABILITY IN RUBISCO SMALL SUBUNITS AND C02 FIXATION
1 2 3 4 5
LSU
.. . . .
further separated on SDS-PAGE gels, the SSUs could be visualized clearly and evenly in all light treatments (Fig. 5, kD panel B). Thus, our results suggest that, in contrast to the homogenous appearance of LSUs in every light treatment,
-66.0 theSSUs of Rubisco from the differentphotoinduced types of gametophytes (R and B) probablyrepresent twodifferent
45.0 typesof antigens. Since whitespectrumconsists of both blue
* and red lights, the SSU population ofthe Wtreatmentappears tobeamixture ofboth SSUtypesand itsantigenicresponse
reflects the blue induced SSUs.
-24.0
Comparison of Rubisco Carboxylase Activity in Extracts
-18.4 of R- and B-Gametophytes
SSU -14.3
s w B R
Figure 2. Total protein pattern of extracts preparedfrom W-(lane 2),B-(lane 3), and R-gametophytes (lane 4).Totalproteinsextracted from 19 d old gametophytesin thepresenceof SDSwereseparated
on a 10 to 20% SDS-PAGE gel. Each lane (2-4) contains 51 ,g protein. Purified spinach(S)Rubisco(12jg)wasusedas amarker toidentify the fernLSUs and SSUs(lane 1).
Identification of RubiscoHoloenzymeandSubunitsin FernGametophytes
Tospecifically identifyRubisco LSUs and SSUs in theR-, B-, and W-gametophytes, equal amountsofprotein extracts wereseparated on SDS-PAGEgels. Thereafter the gelswere
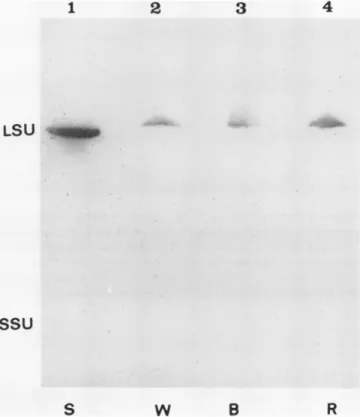
blottedonto nitrocellulose filters andprobedwithpolyclonal antibodiesraised in rabbitagainsteither SDS-treatedspinach LSUorSSU.Figures3 and 4 showtypicalimmunoblotsusing anti-LSU or anti-SSU, respectively, asprobes. LSUs in ex- tracts from each light treatment are equally recognized by anti-LSU sera (Fig. 3). However, R-induced SSUs are not recognized by antibodies raised against SSU from spinach, while B- and W-induced SSUs are well recognized (Fig. 4).
Application ofa double amount ofprotein extract did not improve detection of R-induced SSUs(10).Because R and B LSUsare equally well recognized by spinach anti-LSU sera,
the addition ofa small amount ofanti-LSU, as marker, to the anti-SSU servedtoverifythatequalamountsofRubisco, from each treatment, were indeed loaded on gels. The sur-
prising lack of reactivity of the anti-SSU antibodies with the SSUs oftheR-gametophytesraised thepossibilitythatperhaps the SSUs were entirely missing in the R-induced gameto- phytes.Therefore, the threetypesofproteinextracts,isolated under nondenaturing conditions, were electrophoresed on
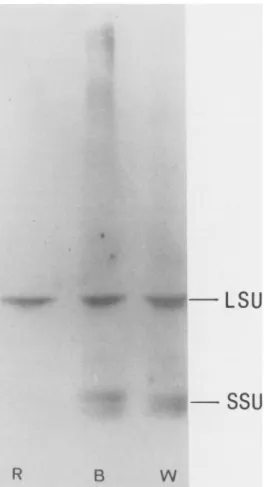
native PAGE gels (Fig. 5, panel A). No difference could be detectedinthe migrationpattern of the holoenzymes in the R, B, or W extracts. Furthermore, when the Rubisco holo-
enzyme bands were excised and eluted from the gel and
Thepresenceof SSUs have been showntobe essential for
propercatalysis. However, theirexactfunction remainsques-
tionable. Thus, the natural occurrence oftwo distinct SSU populations in gametophytic cells while LSUs are identical enabledustoexamine thepossible effectof different SSUson
carboxylation activity of naturally produced holoenzymes.
1 2 3 4
LSU __
SSu
S W B R
Figure 3. Immunodetection of R, B,andWLSUs. Rubiscoenriched extracts (50 ,ug) from 6 d old R, B, and W grown cultures were
separatedon a10 to 20%SDS-PAGE gel similarlytothatin Figure 3. Following blottingontoanitrocellulosefilter,the blotwasprobed withpolyclonal antibodies raised in rabbitagainst SDS-treated spin- ach LSUand then with 1251-donkey anti-rabbit IgG. Purified spinach (S)Rubisco(10 Mig)wasusedas amarkerfor the LSU oftheenzyme.
Figure 1. Morphology of fern gametophytes grown under different light spectra. Fern gametophytes were germinated and synchronized as describedin"Materials and Methods."Afteradditional48 h in dark, the gametophytes were exposed to continuous R (panel A and B), B (panel CandD),orW(panelE andF)light for18d.
301
Plant Physiol.Vol.95, 1991
12 r
~~~~~
10.i
L
E
N
0 6
E
- 4.
i- 2 !
> 2-2
0.00
_-a ~
U
0.75 1.50
RuBP (mM)
Figure 6. Rubisco carboxylase activityas afunction of RuBPcon-
centration.Fern gametophytesweregrownundercontinuous RorB for 18 d. Rubisco enriched extractswereprepared from both illumi- nationgroups asdescribed in "Materials and Methods." Carboxylation activitywasassayed using 27ugRand 39 AgB Rubisco protein in thepresenceof18.2mmNaHCO3 (22.5 !LMC02). Dataareaverage
of threereplicates; coefficient of variationwasless than 15%.
Sssu
B vv
Figure 4. Immunodetection of R, B, and W SSUs. Total soluble proteins (45 ,ug)extracted from 6d old R, B, orWgrowncultures
were separated on a 5 to 20% SDS-PAGE gel. Following blotting ontoanitrocellulosefilter,the blotwasprobedwithrabbitanti-spinach SSUIgGthatalso containedasmall amount ofanti-spinachLSUIgG.
The blotRLibiscowas.#LAthenprobed with_ 440669232
~~~~~~~~~~~~3
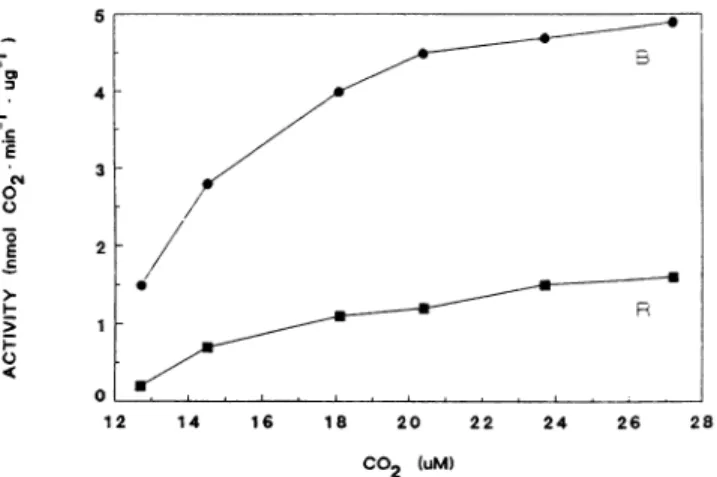
1251-donkey;ILSUSSu anti-rabbit0....t IgG.P0 B We chose to comparethe activity in the R- and B-gameto- phytes only, since these represent homogenouspopulations of SSUs. Rubiscoenrichedextractswereprepared from both R-and B-gametophytes. Themaximalrateofcarboxylation of Rubisco was determined in the presence of increasing concentrations of either ofthetwosubstrates (RuBPorCO2).Figure 6 depicts carboxylation activity as a function of in- creasing concentrations of RuBP. It shows clearly that the carboxylation activity of B-gametophytes is significantly higher than that ofR-gametophytes.Similarly, when Rubisco carboxylation activity was determined in the presence of increasing concentrations of NaHCO3 (CO2) (Fig. 7), again
anapproximately threefold higher maximalrateofcarboxyl- ation is observed inthe B-gametophyteextract. Evidently, a
certain correlation exists between differencesinSSUprimary structureas revealedby its antigenicity and the holoenzyme carboxylation activity.
Figure 5. Characterization of Rubisco holoen-
kD zyme and subunits in fern gametophytes. A,
97 4 Rubisco enriched extracts(18Ag)from 21 dold R,B,and Wgrowncultureswereloadedonto5 66.0 to 25%nondenaturatingacrylamide gelandelec-
trophoresedtoequilibrium parallelyto sizemar- kers. Following electrophoresis, the gel was 45.0 stained with Coomassie brilliantblue. B, Sepa-
290 rationon 10 to 20%SDS-PAGEgelofRubisco bandsexcisedfrom thenondenaturating gel (A) and eluted overnight in SDS-PAGE loading buffer. Afterelectrophoresis,thegelwasstained 14.4 asabove.The left lanerepresentsthepatternof 60 ugtotal proteinsextracted from R-gameto-
w phytesinthepresenceof SDS.
2.25
va
302 EILENBERG ET AL.
I I:
!,f-oi ilia" lw:l---- L SU
VARIABILITY IN RUBISCO SMALL SUBUNITS AND C02 FIXATION
-
E
('4
00 -a E 0
CO2 (uM)
Figure 7. Rubisco carboxylase activityas afunction ofC02concen-
trationintheassay.FerngametophytesweregrownincontinuousR orBfor 24 d. Rubisco enriched extracts wereprepared from both illuminationgroupsasdescribed in "Materials and Methods." Carbox- ylation activitywas assayed using 25 Ag R and 46 qg B Rubisco protein inthe presence of 2 mm RuBP. Data areaverageof three replicates;coefficient of variationwasless than 13%.
DISCUSSION
Growth of fern gametophytes under continuous B or R results in theappearanceoftwodistincttypesof SSUs which
can be distinguished bytheir antigenicity. Although the in- volvement of R and B in the regulation oftranscription of differentrbcSs has beenreported (14),weknow ofnoreports indicating the presence ofsignificantly different translation products in thesame species(22). The naturaloccurrenceof twoSSUpopulationsinthesameplantgrownunderdifferent illumination conditionsenabledustostudythepossiblecor-
relation betweenspecificSSU type andcatalytic propertiesof Rubisco.Indeed, itwasfoundthat Rubiscocarboxylaseactiv- ity in vitro issignificantly higherin extractsfrom ferngame-
tophytes grown under B as compared to R. The enhanced activity could not be accounted for by differential Aco2 *
Mg2+ activation kinetics ofthe enzyme from both sources,
since bothtypesofcarboxylaseswerefound to be fullyacti- vated after 10minpreincubation with HC03-andMg2+ (data notshown).Inaddition,the influence ofapossibleRinduced inhibitor, analogoustothe nocturnalone(17),wasruledout bymeasuring carboxylation activityofenzymeextracted from Rgrowngametophytes thatweretransferredto W24 hprior totheexperiment. No increaseincarboxylation activity could be detected in these extracts (data not shown). It seems,
therefore, that the enhanced Rubisco activity in B-gameto- phytes comparedtoR-gametophytes is dictated by theenzyme
molecule. The presence oftwo distinct SSU populations in the R-andB-gametophytes suggeststhata correlation exists between theSSUprimarystructurerevealedby its antigenicity and theholoenzyme activity.
The functionof SSUhas beenasubject ofdisputeoverthe pastyears. It iswellagreed that thecarbamateformation site (active site) aswell assubstratebinding sites of theenzyme arelocatedontheLSU(3). However,recentworkby Schnei- deretal. (24)suggeststhat the SSU can modulate substrate
bindingby inducing conformational changes of the activesite.
Thedegreeofpartitioningbetweencarboxylationandoxygen- ation(Tvalue)ofRubisco from higherplants as compared to that of less evolved photosynthetic organisms (r = 80 for higherplants, r=60foralgae,andT=50for cyanobacteria) on one hand and the conservation of LSU amino acid se- quencesthroughout evolutiononthe otherhandsuggestthat differences in SSUsmayplayanimportant role in increasing Rubisco net carboxylation activity (3, 19). Nevertheless, no changein theTvalue was revealedby Andrews andLorimer (2) when comparing catalytic properties ofa hybrid between cyanobacterial LSUs andhigher plant SSUs of Rubisco with those ofahomologously reassembled cyanobacterialenzyme.
Similar studies usinghigher plant LSUs have not been per- formed yet due tothe irreversible denaturation of LSUs in theabsence of SSUs and the need forachaperon foracorrect assembly of the LSU octamer(1 1, 15, 26).Usingadifferent approach, the contribution of SSUs to Rubiscoactivity has also been studied inAnacystis nidulans by Voordouwet al.
(27). They performed site-directed mutagenesis oftwo Trp residues in one of the conserved regions of cyanobacterial rbcS. The mutatedenzymehadasimilarTvalue and therefore inthis respectitconfirmsthe resultsofAndrews and Lorimer (2). However, whereas the Km(C02)wasunchanged,the Vmax (CO2) was significantly lower for the mutated enzyme as compared to the wild-type one, demonstrating that single amino acid replacements in thesocalled"noncatalytic"SSU influencethecatalyticrateoftheenzyme. Theseresultssug- gest anactivatingrather thanregulatingrole for the SSU.
The fern gametophytesare aunique system forthestudy of the contribution of SSUstoRubiscoactivity. Thissystem enablesus toexaminenaturallymodifiedsourcesof rbcSsby using the special photoinduced occurrence of two distinct SSU populations. Although we have not determined the T
values of the R- and B-gametophytes, our results clearly suggest afunctional role for the SSUs.Hence,comparisonof SSU aminoacidsequencesin R-andB-gametophytes might indicate specificamino acid differences affectingcarboxyla- tionactivity.Acomputerprediction (9)of theantigenicindex of tobacco SSU shows that theevolutionaryconservedregion, composedof amino acids40to70 inthematurepolypeptide, is expectedtobe the mostantigenic. Interestingly, the point mutations in AnacystisrbcS affecting carboxylationactivity, introduced by Voordouw etal. (27), are alsolocatedin this region. Thus, it is plausiblethat the differentialimmunore- cognition observed in the fern gametophyte SSUs could be due to changes in amino acid sequences in this particular region.
Incontrast tothe rbcL,transcription ofrbcSs is known to belight-regulated,both through thephytochrome system and via blue light receptor (12, 13). Quantitative regulation of rbcShas beenattributed mainlyto sequences 5' and/or 3' to the codingregion (5, 6, 20). Differencesbetweenexpression ofrbcSgenes doexist betweendifferentorgansand atvarious developmental stages (7). Thus, it is possible that under variouslightspectradifferent rbcSsfrom themultigene family are expressed. Currently the fern rbcS gene-family is being studied inorder topointoutpossibledifferences in rbcSexons thatcouldaffect carboxylaseactivity.
4-
3-
2-
0
1 1 20 2 2 2 28.
12 14 16 18 20 22 24 26 28
B
303
LITERATURE CITED
1. Andrews TJ, Ballment B (1983) The function of the small subunits of ribulose bisphosphate carboxylase-oxygenase. J Biol Chem 258:7514-7518
2. Andrews TJ, Lorimer GH (1985) Catalyticpropertiesofahybrid betweencyanobacteriallarge subunits andhigher plantsmall subunits of ribulose bisphosphate carboxylase-oxygenase. J Biol Chem260: 4632-4636
3. Andrews TJ,Lorimer GH (1987) Rubisco:structure,mechanism andprospectsforimprovement.In MDHatch,NKBoardman, eds, TheBiochemistryofPlants, Vol 10.AcademicPress, New York,pp 131-217
4. Avni A,Edelman M,Rachailovich I,Aviv D,Fluhr R(1989)A point mutationinthegenefor thelargesubunit of ribulose1,5 bisphosphate carboxylase/oxygenase affects holoenzyme as- semblyinNicotianatabacum.EMBOJ8: 1915-1918 5. DeanC, Favreau M,Bond-Nutter D, Bedbrook J, Dunsmuir P
(1989) Sequences downstream of translation start regulate quantitive expression oftwopetuniarbcSgenes. PlantCell 1:
201-208
6. DeanC,FavreauM, BedbrookJ,Dunsmuir P(1989)Sequences 5' tothetranslationstartregulateexpression of petunia rbcS
genes.PlantCell 1: 209-215
7. DeanC, Pichersky E, DunsmuirP (1989) Structure, evolution andregulation of rbcSgenesinhigher plants.AnnuRevPlant PhysiolPlantMol Biol 40:415-439
8. Deng X-W, GruissemW(1987)Control ofplastidgeneexpres-
sion during development: the limited role oftranscriptional regulation. Cell49:379-387
9. DevereuxJ,Haeberli P,Smithies0(1984)Acomprehensiveset ofsequenceanalysisprogramsfor the VAX.NucleicAcids Res 12:387-395
10. EilenbergH,BeerS, GepsteinS,Geva N, Zilberstein A (1990) Redorblue lightinduceddifferent rubisco small subunits in ferngametophytes affecting carboxylation rates. In M Balts- cheffsky, ed,Current ResearchinPhotosynthesis,Vol3, Klu-
werAcademic Publishers, Dordrecht,pp 11.411-11.414 11. EllisRJ,vanderViesSM (1988)The rubisco subunit binding
protein. Photosynth Res16: 101-115
12. FluhrR,Chua NH(1986) Developmentalregulationoftwogenes
encodingribulose-bisphosphate carboxylase small subunitin peaandtransgenicpetunia plants: phytochromeresponseand bluelightinduction. Proc Natl Acad Sci USA 83: 2358-2362 13. FluhrR,KuhlemeierC,NagyF,Chua N-H(1986) Organ-specific
andlight-induced expressionofplantgenes.Science 232:1106- 1112
14. Fluhr R(1989)Structure andregulationoflight-induciblegenes:
genesinvolved inphotosynthesis. In J Schell, IK Vasil, eds, Cell Culture and Somatic Cell Genetics of Plants, Vol 6.
AcademicPress,NewYork,pp 133-153
15. Goloubinoff P, Gatenby AA, Lorimer GH (1989) GroE heat- shockproteinspromoteassemblyofforeignprocaryotic ribu-
lose bisphosphate carboxylase oligomers in Escherichia coli.
Nature 337: 44-47
16. GruissemW(1989)Chloroplast gene expression: how plants turn theirplastidson.Cell 56: 161-170
17. Gutteridge S, Parry MAJ, Burton S, Keys AJ, Feeney J, Ser- vaitesJC, Pierce J (1986)Anocturnalinhibitor of carboxyla- tion in leaves. Nature 324: 274-276
18. Gutteridge S,GatenbyAA(1987) The molecularanalysisof the assembly,structureandfunctionofRubisco. In BJMiflin,ed, Oxford Surveys of Plant Molecular & Cell Biology, Vol 4.
Oxford University Press, New York, pp 95-135
19. Jordan DB, Ogren WL(1981)Speciesvariation in the specificity of ribulose bisphosphate carboxylase/oxygenase. Nature 291:
513-515
20. KuhlemeierC, FluhrR,Green PJ,ChuaN-H(1987) Sequences in the pearbcS-3A gene have homology to constitutive mam- malian enhancers but function as negative regulators. Genes Dev 1:247-255
21. Marder JB, Mattoo AK,EdelmanM(1986) Identification and characterization of the psbA gene product: the 32-kDa chlo- roplast membraneprotein. Methods Enzymol 118: 384-396 22. Nagy F, Fluhr R,Morelli G, Kuhlemeier C, Poulsen C, Keith B,
Boutry M,ChuaN-H(1986) The rubisco small subunit gene as aparadigmfor studies on differential gene expression during plant development. Phil Trans R Soc Lond B Biol Sci 313:
409-417
23. NelsonN(1983) Structure andsynthesis of chloroplast ATPase.
Methods Enzymol 97: 510-523
24. Schneider G, Knight S, Andersson I, BrandenC-I,Lindquist Y, Lundquist T(1990)Comparisonof the crystal structures of L2 and L8S8 Rubisco suggests a functional role for the small subunit. EMBO J 8: 2045-2050
25. Tobin EM, Silverthorne J (1985)Light regulation of gene expres- sion inhigher plants.AnnuRevPlantPhysiol 36: 569-593 26. VoordouwG, van der Vies SM, Bouwmeister PJ (1984) Disso-
ciation of ribulose-1,5-bisphosphate carboxylase/oxygenase from spinach by urea. Eur J Biochem 141: 313-318
27. VoordouwG, De Vries PA,vandenBerg WAM, De Clerck EPJ (1987)Site-directed mutagenesis of the small subunit of ribu- lose-1,5-bisphosphate carboxylase/oxygenase from Anacystis nidulans. EurJBiochem 163: 591-598
28. Wolter FP, Fritz CC, Willmitzer L,SchellJ,SchreierPH(1988) rbcS genes in Solanum tuberosum: conservation of transit peptideandexon shuffling duringevolution. Proc Natl Acad Sci USA 85: 846-850
29. Zemel E, Gepstein S (1985) Immunological evidence for the presence of ribulose bisphosphate carboxylase in guard cell chloroplasts. PlantPhysiol78:586-590
30. Zilberstein A, ArzeeT,Gressel J (1984) Early morphogenetic changesduringphytochrome-inducedfern sporegermination.
I. Theexistenceofapre-photoinductionphase and theaccu- mulation of chlorophyll.ZPlanzenphysiol114:97-107