With 2 text-figures Printed in Great Britain
WEIGHT DISCRIMINATION BY OCTOPUS
BY M. J. WELLS
Department of Zoology, University of Cambridge
{Received 28 July i960)
INTRODUCTION
Blinded octopuses can readily be taught to discriminate between objects touched provided that these differ in 'roughness'; objects with equally irregular surfaces, differing only in the distribution of irregularities, and objects of similar texture but different shape, are not distinguished. It seems that the animals are incapable of learning to recognize specific patterns of stimulation in the tactile sensory field, or to recognize how the arms must bend around objects grasped; they evidently distinguish things that they touch simply from the proportion of sense organs excited in the area of contact (Wells & Wells, 1957). The failure to take into account the relative positions of the sense organs excited seems best explained by supposing that Octopus is incapable of integrating proprioceptive with surface tactile informa-tion in learning to recognize objects by touch. This hypothesis is interesting because, if true, it would mean that the tactile learning system of Octopus is peculiarly simple, a mechanism limited to making only a quantitative distinction between surfaces touched. A direct test of the matter is to train octopuses to dis-criminate between objects of similar texture but different weight; such objects cannot be identified on a basis of surface tactile information alone, but can readily be distinguished by any animal capable of learning to recognize tension or pressure differences within the parts of its own body handling the objects.
The experiments reported here show that octopuses cannot be taught to dis-criminate between objects differing only in weight, although they very readily learn to make textural discriminations under the same conditions.
MATERIAL AND METHODS
Octopus vulgaris Lamarck of between 250 and 850 g. were used in these experiments.
The animals were collected in the Bay of Naples and kept separately in large asbestos tanks. A few days before the beginning of training the octopuses were blinded by section of the optic nerves, as described in Wells & Wells (1956).
presented, and rejecting t h e 'negative' b y thrusting it away t o arm's length. Errors at t h e start of a n experiment are characteristically b y failure to reject the negative object, since all unfamiliar objects are at first passed to the mouth.
F o r t h e present series of experiments two training routines were used. I n one ( ' l o n g - t e r m ' conditions) there were 8 trials per day at intervals of not less than i h r . systematized t h u s : -\ 1 M first day, —| M h second day, and so on. I n the second ('short-term' conditions) there were five times as m a n y trials per day, arranged in two groups of twenty trials n o t less than 6 hr. apart. W i t h i n each group t h e trials, at intervals of 5 min., were systematized t h u s :
-\ 1 hH 1 1 1 h H 1 — . N o gross differences in
perfor-m a n c e occurred under t h e two sets of conditions.
Table 1. Details of the test objects used
(All were cylindrical, 3-0 cm. long, cut from Perspex rod of 2-5 cm diameter.)
P i
P4 P8
P4H1 P4H2 P8H
Surface texture Vertical grooves Smooth
Circumferential grooves Smooth
Smooth
Circumferential grooves
Proportion of total surface
cut away as grooves (%)
3° 5° — 5°
Weight in sea water to
nearest g.
5
s s
45
IS 25
Pictures of Pi, P4, P8, P4H1 and P8H are included in fig. 2.
Characteristics of the test objects used
Six different cylindrical Perspex objects were used in these experiments. Each object was 3-0 cm. long and 2-5 cm. in diameter. Three of the cylinders were left smooth (P4, P4H1 and P4H2), and three (Pi, P8 and P8H) had deep, 1 mm. wide, grooves cut into their otherwise smooth surfaces. Two of the smooth objects (P4H1 and P4H2) and one of the grooved cylinders (P8H) were hollowed out, filled with lead and sealed with a Perspex plug. A more detailed specification of each object is given in Table 1, and pictures of most of them included in Fig. 2.
Behaviour of octopuses in handling the heavy objects
Weight discrimination by Octopus 129
tone of the animal resting on the side of its tank was sufficient to support the weight. In the case of the weighted objects, and particularly of the very heavy P4H1 (45 g.), the situation was different. Here slacking the line frequently resulted in con-siderable passive extension of the arm grasping the object; the effect was transient, the animal rapidly contracting its arm and supporting the weight, but was nearly always clearly visible, so that an observer could identify the object presented by the octopus's reaction to it. It will be shown below that despite the obvious muscular adjustments needed to support the heavy cylinders, octopuses were unable to distinguish these from much lighter cylinders of similar surface texture.
EXPERIMENTAL RESULTS Long-term experiments
Octopuses were trained to discriminate between P8 and P4, Pi and P4, and Pi and P8, all pairs of objects differing in texture, and between P4H2 and P4, which were alike in texture but different in weight. The animals readily learned to make the distinctions based on textural differences, but failed to learn to discriminate between P4, weighing 5 g., and P4H2, weighing three times as much (Fig. 1). In these experiments nearly all the errors were due to acceptance of the 'negative' object, which the animals were punished for taking, and in the weight discrimination tests five out of the six octopuses continued to take very nearly all of the objects presented throughout the 128 trials (16 days) of their training; these five animals together re-jected only three objects in the whole course of training (2 negatives, 1 positive). The remaining animal concerned in the weight-discrimination experiment did reject a proportion of the test objects in the last 9 days of its training, but its per-formance (it rejected the negative object three times, and the positive eight times) was erratic, and gave no indication of learning to make the discrimination.
In the interpretation of these results it must be borne in mind that octopuses con-tinuing to take both members of a pair of objects are not necessarily unable to dis-criminate between them; in retention tests, for instance, a reduction in the over-riding tendency to react positively often reveals a capacity to discriminate that is otherwise masked (Wells & Wells, 1958). Although this was thought unlikely to account for the failure to discriminate between P4H2 and P4, further experiments were made under the short-term conditions, where the tendency to err only by acceptance of the negative object is normally less marked.
Short-term experiments
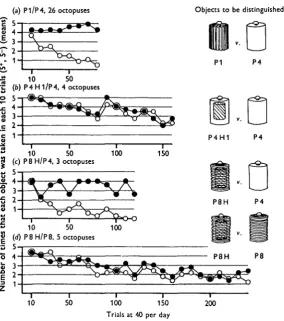
Four octopuses were trained to discriminate between P4 and P4H1 (a second smooth object, nine times as heavy as P4) at a rate of 40 trials per day. In Fig. 2 the performance of these animals is compared with that of 26 other octopuses trained to discriminate between the grooved Pi and the smooth P4. Once again the textural discrimination was learned rapidly, while the octopuses failed to make the weight discrimination. Over the period of training the proportion of positive responses by the four animals trained on P4H1/P4 declined until by the end of the
160 training trials less than half of the objects presented were accepted; under these conditions any tendency to discriminate should have been manifest were the octopuses at all capable of distinguishing between the objects. In fact the four animals together took the positive object 240 times and the negative 234 times in 320 presentations of each; even in the last 80 trials the scores were respectively 112 and 103 positive responses to the objects to be discriminated. Clearly these are chance scores and show no evidence of discrimination.
(a) P8/P4,12 octopuses
8 40 (b) P1/P4,13 octopuses
I
80
_ 8 40 £ (d) P4H2/P4, 6 octopuses
° 4 -| ©®(§®®(D 9-8
%--• 3
Z 1
-40 80 Trials at 8 per day
120
Fig. I. A summary of training experiments carried out under the 'long-term' conditions. The plots show the number of times that each of the objects to be discriminated was taken on each day of 8 trials (4 + , 4 — ); • = positive object (which the octopus was rewarded for taking), O = negative object (octopus given a 6-10 V. a.c. shock for taking); where two points coincide the two are plotted ©. In (a), (6) and (c) octopuses were trained to discriminate between Perspex cylinders of the same weight differing in surface texture; the three discriminations are arranged in order of difficulty for Octopus. In (d) an attempt was made to train octopuses to distinguish between P4 and P4H2, an object three times as heavy as P4, but similarly smooth in texture. In each group approximately half of the animals were trained with one object as the positive and half with the other.
Weight discrimination by Octopus 131
The performance of the three animals trained to discriminate between P8H and P4 may be compared with that of the 26 animals trained to distinguish between Pi and P4 (Fig. 2). The two discriminations were learned about equally readily. Since P8H/P4 is on texture alone somewhat the easier discrimination of the two (see P8/P4 and P1/P4 in Fig. 1) this result again suggests that weight is not a property of objects that Octopus can learn to recognize.
(a) P1/P4, 26 octopuses Objects to be distinguished
P1 P4
10 50 P4H1/P4. 4 octopuses
P 4 H 1 P4
10 50 100 P8H/P4, 3 octopuses
150
O-O 10 50 100 P8H/P8, 5 octopuses
P8H P4
P8H P8
50 100 150
Trials at 40 per day
[image:5.451.85.370.169.491.2]200
Fig. 2. The results of training experiments made under the 'short-term' conditions, with 40 trials per day in two groups of 20. In (a) twenty-six octopuses were trained to discriminate between the smooth P4 and the grooved P1, both objects being of the same weight (5 g.). In (6) an attempt was made to train four octopuses to distinguish between P4 and P4Hi, a second smooth object nine times as heavy as the first. In (c) three animals were trained to discriminate between P8H and P4, which differed in both weight and texture. In (d) an attempt was made to train five animals, two of them already trained successfully under (c) to discriminate between P8H (weighing 25 g.) and P8 (5 g.). Experimental results plotted as in fig. 1. Some further details of individual scores are given in Table 2.
The attempt to train five octopuses to discriminate between P8H and P8 (P8H being five times as heavy as P8) was not successful (Fig. 2 d). The performance of the animals was erratic, and although, over the 240 trials of training to which each was
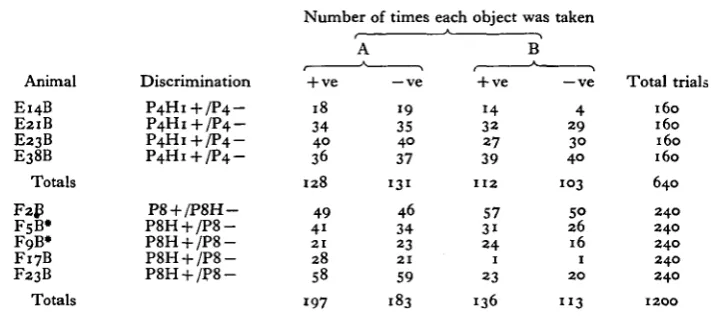
Table 2. Details of individual performances from the experiments
summarized in Fig. 2
(Columns A and B show scores in the first and second halves of the period of training) Number of times each object was taken
Animal E14B E21B E23B E38B Totals F2B F5B* FoB* F17B F23B Totals Discrimination P 4 H 1 + / P 4 P 4 H 1 + / P 4 P 4 H 1 + / P 4 P 4 H 1 + / P 4
-P8 + /-P8H- /P8HP8H + / P 8 P8H + / P 8 P8H + / P 8 P8H + / P 8
-+ ve 18 34 40 36 128 49 4i 21 28 58 197 A — ve 19 35 40 37 131 46 34 23 21 59 183 + ve 14 32 27 39
1 1 2
57 3i 24 1 23 136 B — ve 4 29 30 40 103 50 26 16 1 20 " 3 Total trial 160 160 160 160
6 4 0
240 240 240 240 240 1200
P4H1 was nine times the weight of P4, and P8H five times the weight of P8. * F5B and F9B were pretrained to discriminate between P8H+ and P4 (see text).
subjected, every animal took the positive object slightly more often than the negative (regardless of which this was, P8H or P8), the difference in total number of takes was small; the positive object was taken 333 times out of a possible 600 and the negative 296 times (x2 = i-o6, P = 0-3); details of individual scores are given in Table 2. When training was continued for a further 80 trials with three of the animals, the positive object was taken 91 times and the negative 65 times out of a possible 120 (x2 = 4 7 , P = 0-03). It is worth noting that this bias towards the positive object was found only in the P8H/P8 experiments, where there was a possibility of small textural differences between the objects due to inaccuracies in cutting the grooves; it has already been shown elsewhere that octopuses can detect quite minor differences of this sort (Wells & Wells, 1957). In the P4H1/P4 experi-ments, where the weight difference was greater than in P8H/P8, two out of the four animals took the negative object more often than the positive.
DISCUSSION
Weight discrimination by Octopus 133
This is interesting because of its relation to facts that have already been collected about the organization of the central nervous system of Octopus. It has been shown that octopuses fail to make tactile discriminations that would depend upon their taking into account the relative positions of their tactile sense organs. They cannot, for example, be taught to discriminate between cylinders (of the P1/P8 type) that differ only in the pattern or orientation of grooves cut into them, and it seems that they cannot learn to discriminate between objects of different shape but similar texture (Wells & Wells, 1956, 1957). The failures are in discriminations that could in principle be made readily were the animals able either to scan the objects by moving their arms over them and integrate information about these movements, or to take into account the relative positions in space of the suckers in contact. Evidently octopuses cannot do either of these things in learning to recognize objects by touch, perhaps because there is no proprioceptive input to the inferior frontal system in the supraoesophageal part of the brain; for it is here that tactile learning takes place (Wells, 1959). The failure to learn to recognize the weight of objects supported by the arms is susceptible to the same explanation; proprioceptive information must enter the central nervous system—it is difficult to see how the animal could co-ordinate movements of the eight arms without it—but is evidently not available for integration with other sensory information in learned processes. A recent confirma-tion of this has come from the demonstraconfirma-tion that learned visual discriminaconfirma-tion of orientation in Octopus depends upon the constant orientation of the retina and not upon central integration of positional information with the visual input (Wells, i960).
SUMMARY
1. Blinded octopuses can readily be trained to discriminate by touch between objects differing in texture.
2. They seem unable to discriminate between similar objects differing only in weight.
3. The relevance of this to what is already known about the use of proprioceptive information in learning by Octopus is discussed.
The author would like to thank the Director and staff of the Stazione zoologica di Napoli for their hospitality and for the facilities that they provided during the summers of 1958 and 1959, when these experiments were being made.
REFERENCES
WELLS, M. J. (1959). A touch learning centre in Octopus. J. Exp. Biol. 36, 590-612.
WELLS, M. J. (i960). Proprioception and visual discrimination of orientation in Octopus. J. Exp.
Biol. 37, 489-99.
WELLS, M. J. & WELLS, J. (1956). Tactile discrimination and the behaviour of blind Octopus.
Pubbl. Staz. zool. Napoli, 28, 94-126.
WELLS, M. J. & WELLS, J. (1957). The function of the brain of Octopus in tactile discrimination.
J. Exp. Biol. 34,