Assessing the reliability of pRIA for identifying ancient proteins from
archaeological contexts
Ben A. Potter
a,*, Joshua D. Reuther
b,c, Jerold M. Lowenstein
d, Gary Scheuenstuhl
d aDepartment of Anthropology, University of Alaska, Fairbanks, AK 99775-7720, USAbDepartment of Anthropology, University of Arizona, Tuscon, AZ 85721-0030, USA cNorthern Land Use Research, Inc., 600 University Avenue, Suite 6, Fairbanks, AK 99709, USA
dDepartment of Nuclear Medicine, California Pacific Medical Center, 2333 Buchanan Street, San Francisco, CA 94115, USA
a r t i c l e
i n f o
Article history:
Received 22 September 2009 Received in revised form 12 November 2009 Accepted 17 November 2009
Keywords:
Protein radioimmunoassay Protein residues Gerstle River site Croxton site Alaska
a b s t r a c t
The use of immunological techniques for identifying the origin of proteins and inferring foodstuffs exploited by prehistoric occupations has been conducted for several decades. Cases of experimental laboratory and archaeological studies have shown the potential of these techniques for reliable results. However, very few of these case studies employ archaeological sites that have excellent preservation and high-resolution spatial contexts between identifiable faunal remains, features, and stone tools. We present an assessment of the identification reliability of one immunological technique, protein radio-immunoassay (pRIA), using faunal remains and stone tools from two sites from arctic and subarctic contexts. The results of this research indicate that, even in contexts with excellent preservation, the identifications produced by the pRIA technique are subject to misidentification and cross-reactions due to diagenetic alteration of proteins. We propose a higher minimum reaction value (percent binding of labeled antibody) that mitigates these effects, and renders the pRIA results more reliable for ancient, poorly preserved organic remains.
Ó2009 Elsevier Ltd. All rights reserved.
1. Introduction
Archaeological residue studies over the past 30 years have illustrated the potential utility of immunological techniques for identifying ancient proteins and the benefits these identifications can provide on several archaeological problems. Archaeological questions that immunochemistry techniques can contribute to our knowledge include: potential direct evidence on the role of humans in the extinction of Pleistocene species (Loy and Dixon, 1998; Kooyman et al., 2006); the ability to provide relative minimum age estimates on tools with adhering residues of extinct animals such as mammoth, mastodon, and North American Equids (Loy and Dixon, 1998); the relationship between artifacts and faunal remains with less than desirable associative contexts (e.g., disturbed and/or surface finds; Dixon and Marlar, 1997; Kooyman et al., 2001; Reuther and Gerlach, 2006); the function of different artifact types and the nature of paleo-subsistence (Gerlach et al., 1996; Seeman et al., 2008); and human sacrifice rituals and cannibalism (Bourget and Newman, 1998; Marlar et al., 2000).Child and Pollard (1992),
Marlar et al. (1995), andSmith and Wilson (2001)provide excellent reviews of the use of these techniques in archaeological studies.
While many immunologically based techniques have been used in forensic cases, the complexity of interactions of environmental agents that act to degrade proteins is typically minimized. Forensic specimens tend to be acquired less than 50 years, perhaps within days, after being deposited into the lithosphere, lessening the length of time proteins are subjected to diagenetic agents such as microbes. The major difference between forensic applications of immunological techniques and their use in archaeology is the fact that the denaturing process is obviously enhanced in ancient contexts. Experiments designed to assess the reliability of these techniques to identify ancient proteins require that the complex-ities of environmental processes and time that effects diagenesis of organic materials be accounted for.
Several studies have attempted to assess the reliability of these techniques in identifying ancient proteins with varying success. These reliability assessments include: the use of known modern residues and bone in controlled laboratory and outdoor experiments (Barnard et al., 2007; Cattaneo et al., 1993; Eisele et al., 1995; Gurfinkel and Franklin, 1988; Hyland et al., 1990; Mauldin et al., 1995; Leach and Mauldin, 1995; Smith and Wilson, 1992; Tuross et al., 1996); comparisons of the results on archaeological materials between *Corresponding author. Fax:þ1 907 474 7453.
E-mail address:[email protected](B.A. Potter).
Contents lists available atScienceDirect
Journal of Archaeological Science
j o u r n a l h o m e p a g e : h t t p : / / w w w . e l s e v i e r . c o m / l o c a t e / j a s
different immunological techniques (Downs and Lowenstein, 1995); comparisons of results on archaeological materials between immu-nological and non-immuimmu-nological techniques (e.g., DNA;Newman et al., 1998); and the use of a site with exceptional spatial and strati-graphic integrity and excellent preservation to compare immuno-logical results derived from proteins adhering to stone tools to a list of species based on associated and identifiable faunal remains ( Kooy-man et al., 1992; NewKooy-man et al., 1996; Shanks et al., 1999). Many of these studies have provided equivocal results on the reliability of these techniques to accurately identify ancient residues.
Our studies on protein radioimmunoassay (pRIA) have concen-trated on assessing the reliability of the identifications of modern and ancient proteins (Lowenstein et al., 2006; Reuther et al., 2006; Ubelaker et al., 2004). Essential to assessing this technique’s reli-ability is to understand the nature of potential cross-reactions that can lead to false-positive identifications. Experiments using modern blood samples and bones indicated that the technique has the potential to reliably identify the origin of proteins, in many instances, to a Genus level of the Linnaean taxonomy (Lowenstein et al., 2006; Reuther et al., 2006). In this paper, we present an assessment of pRIA in the identification of ancient proteins using faunal remains and stone tools from two Alaskan prehistoric sites, the Gerstle River and Croxton sites, both with excellent archaeo-logical contextual control and preservation.
2. Methods
2.1. Site and sample selection
Identified faunal remains and stone tools were selected from two buried, prehistoric sites in Alaska: Gerstle River, located in the Tanana River Lowlands currently in the boreal forest, and Croxton, located in the Brooks Range. Both sites are described below.
Gerstle River is situated on a bedrock outcrop overlain with deep (>4 m), stratified aeolian deposits (Potter, 2005, 2007). Each of the
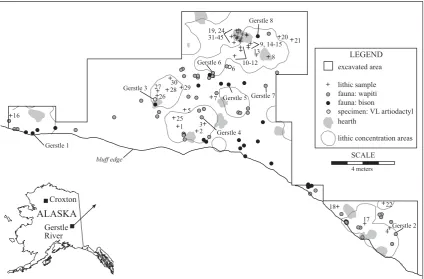
five components is well separated by sterile sediments, and the radiocarbon chronology is very consistent (Potter, 2008). Compo-nent 3 occurred during loess deposition between 10,300 and 9600 cal BP. Component 3 consists of over 7000 lithic artifacts directly associated with 13 unlined hearths and abundant faunal material in a ‘‘living floor’’ that forms a spatially integrated depo-sitional set (Fig. 1;Potter, 2005: 156–174). Evidence for contem-poraneity include: a tight unimodal vertical distribution of all artifacts and ecofacts, rapid burial after deposition, lack of post-depositional disturbance such as cryoturbation, refits between areas, and contemporaneous radiocarbon dates (averaging w10,100 cal BP) (Potter, 2005: 697–747).
Sediment analyses indicate well-drained sediments with little potential for leaching of residual proteins from the faunal remains. The collagen yields for the faunal remains are relatively high, and the radiocarbon results from the site (using ultrafiltration pretreatment) show a tight distribution (identical with the hearth charcoal dates) (Potter et al., n.d.). Results for pH indicate a basic environment (8.0) in Component 3 stratum (pedogenic calcium carbonates are present in lower strata at the site). Loss-on-ignition results indicate low percent carbon (1.64% by weight), calcium carbonate (5.45%), and water (3.03%) in the Component 3 stratum (Potter, 2005). In sum, organic preservation is high, and we expect little chance of contamination of faunal samples or lithic residues with respect to groundwater leaching, humic acids, or other soil-related chemical processes.
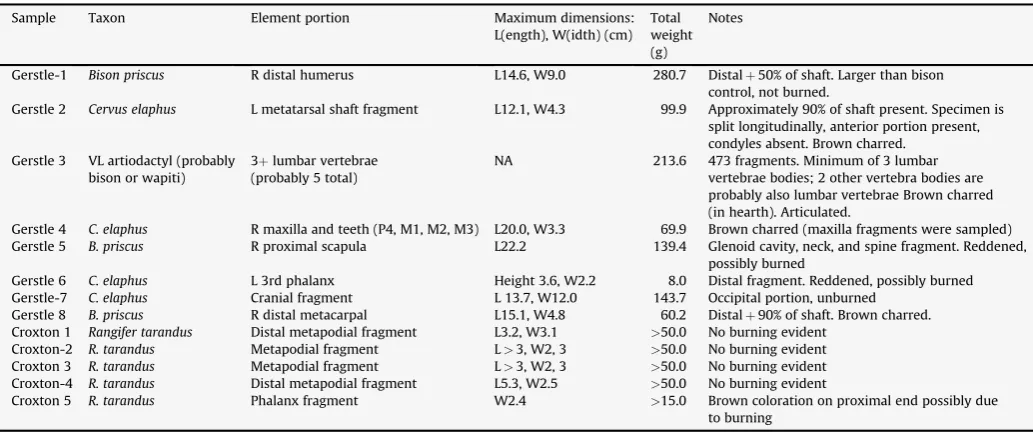
Faunal specimens from Gerstle River, Component 3, are rela-tively well preserved for their age (Potter, 2007). Identifications of the 8 specimens sampled for this research were made based on morphology, and summary information is provided in Table 1. Component 3 faunal remains compriseCervus elaphus (wapiti or elk), Bison sp. (bison), or very large (VL) artiodactyls, which are probably wapiti or bison, but are too fragmented to definitively identify. All cervid remains from this component were wapiti-sized (not moose or caribou-sized), and elements with distinctive wapiti
or moose identification landmarks (e.g., distal metapodials) indi-cate wapiti was the only cervid in Component 3. Bison from the site dating tow11,000 cal BP were sampled for mtDNA analyses, and results indicate they areBison priscus, steppe bison (Shapiro et al., 2004). Thus, we consider the taxonomic identifications to be robust for Gerstle River.Fig. 1illustrates the spatial locations of the iden-tified specimens and the samples included in this research.
The 45 stone tools that were sampled from Component 3 are in close proximity to the faunal remains in inferred butchering activity areas, and thus have potential to contain organic residue (Fig. 1). Lithic artifacts include boulder spall scrapers (n¼12), burin (n¼1), end scrapers (n¼2), microblades and microblade frag-ments (n¼17), and modified flakes (n¼13). These tools are inter-preted to be associated with hunting implements (microblades used as insets in organic weapon tips), fabricators (burins used to incise organic weapon tips), and butchery/processing implements (modified flakes, boulder spall scrapers, the latter possibly used in hide processing). End scrapers probably had multiple functions, but could have been used in working soft material (e.g., hides), and medium or hard materials (e.g., wood, bone, antler). Two toolsets are inferred at Gerstle River: (1) a curated toolset dominated by hunting implements (personal hunting gear), deposited near the hearths (microblades, burins), and (2) expedient faunal processing toolset (boulder spall scrapers, retouched flakes) of primarily local materials spatially associated with the interface between faunal and lithic concentrations.
The first set of lithic samples (#1–30) were selected in 2005, many years after they were first recovered from the site. Multiple individuals handled these artifacts during excavation and analysis, and many were cleaned using a dry brush during accessioning. These lithic samples are labeledGroup A. Because handling may affect the pRIA results, a portion of the site was excavated in 2006, with specific safeguards against contamination or removal of residual proteins. Excavators used latex gloves and artifacts were minimally handled and were not brushed. Fifteen artifacts (#31–45) were placed in archival (non-reactive) plastic bags immediately upon excavation and sent in for pRIA analysis, labeled Group B. Two analytical runs were made on several specimens (bone samples Gerstle #1–3 and lithic samples #1–5; 10–12; 15–17; 19–20) in order to analyze additional taxa.
The Croxton site is situated on the southeastern shore of Tukuto Lake in the northern foothills of the central Brooks Range, over 200 miles north of the Arctic Circle (Gerlach, 1989). The archaeological deposits at Locality J of Croxton are within fluvial and colluvial sediments. The majority of archaeological remains at Locality J are associated with caribou procurement and processing activities that date between 1650 and 790 BP (Gerlach, 1989; Reuther, 2003). Sediments are perennially frozen due to the site’s high latitude location [68300 N]. Organic preservation is excellent at the site with immense amount of wood, antler and bone being recovered during intermittent excavation for the past 25 years (1981–1983, 2000). Range of species identified from the faunal remains include avifauna, bear, beaver, canids, caribou, fish, fox, ground squirrel, sheep, and other less identifiable remains of large, medium and small mammals (Gerlach, 1989). Five bone fragments from Croxton identified as caribou remains were analyzed (Table 1). The samples are extremely well preserved, consistent with remains recovered from permafrost contexts at high latitudes.
These data allow for control of certain variables to establish accuracy and precision for pRIA in archaeological contexts. This dataset derived from Gerstle River and Croxton is used to analyze (1) bone specimens morphologically identified to taxa, (2) bone specimens relative to taphonomy (different ages and geological contexts), (3) residues on associated lithics, (4) potential for human contamination using different handling procedures, and (5) multiple analyses on the same specimens.
Table 1
Faunal sample information.
Sample Taxon Element portion Maximum dimensions:
L(ength), W(idth) (cm) Total weight (g)
Notes
Gerstle-1 Bison priscus R distal humerus L14.6, W9.0 280.7 Distalþ50% of shaft. Larger than bison
control, not burned.
Gerstle 2 Cervus elaphus L metatarsal shaft fragment L12.1, W4.3 99.9 Approximately 90% of shaft present. Specimen is split longitudinally, anterior portion present, condyles absent. Brown charred.
Gerstle 3 VL artiodactyl (probably bison or wapiti)
3þlumbar vertebrae (probably 5 total)
NA 213.6 473 fragments. Minimum of 3 lumbar
vertebrae bodies; 2 other vertebra bodies are probably also lumbar vertebrae Brown charred (in hearth). Articulated.
Gerstle 4 C. elaphus R maxilla and teeth (P4, M1, M2, M3) L20.0, W3.3 69.9 Brown charred (maxilla fragments were sampled)
Gerstle 5 B. priscus R proximal scapula L22.2 139.4 Glenoid cavity, neck, and spine fragment. Reddened,
possibly burned
Gerstle 6 C. elaphus L 3rd phalanx Height 3.6, W2.2 8.0 Distal fragment. Reddened, possibly burned
Gerstle-7 C. elaphus Cranial fragment L 13.7, W12.0 143.7 Occipital portion, unburned
Gerstle 8 B. priscus R distal metacarpal L15.1, W4.8 60.2 Distalþ90% of shaft. Brown charred.
Croxton 1 Rangifer tarandus Distal metapodial fragment L3.2, W3.1 >50.0 No burning evident
Croxton-2 R. tarandus Metapodial fragment L>3, W2, 3 >50.0 No burning evident
Croxton 3 R. tarandus Metapodial fragment L>3, W2, 3 >50.0 No burning evident
Croxton-4 R. tarandus Distal metapodial fragment L5.3, W2.5 >50.0 No burning evident
Croxton 5 R. tarandus Phalanx fragment W2.4 >15.0 Brown coloration on proximal end possibly due
to burning
Table 2
List of reagents used in pRIA analysis.
Control sera Anitsera to Antigen
Bear Bear Albumin
Bison Bison Albumin
Chicken Chicken Albumin
Dog Dog Albumin
Elephant Elephant Albumin
Tule Elk Tule Elk Albumin
Human Human Albumin
Musk-ox Musk-ox Albumin
Sheep Sheep Albumin
2.2. pRIA technique
Lowenstein and Scheuenstuhl carried out solid-phase double-antibody radioimmunoassay (pRIA) on each bone extract and stone tool. pRIA is a highly sensitive immunological technique able to detect to 0.001 mg of antigen with a similar quantity of antibody. This high sensitivity of pRIA provides a means for detecting resi-dues that other immunological techniques (CIEP, ELISA) can miss (Downs and Lowenstein, 1995). The exterior of bone samples were cleaned with a 1 mm stainless steel Dremel drill bit to remove and discard the outerw2 mm surfaces. The remaining material was considered free of external protein contamination and powderized using a second drill bit. Both drill bits were sterilized. Proteins were brought into solution by placing powdered samples in a 10 ml Vacutainer and 1 M of EDTA was added through the septum of the cap by a syringe. Bone particles were dissolved in the EDTA solution through gentle agitation of the container for 2–5 days.
pRIA was conducted on the bone extracts by placing 20 micro-liters EDTA extracts into polystyrene microtiter plate wells. After remaining in the microtiter for 1 h at room temperature, protein binds to the plastic walls of the wells. The liquid extracts were washed out with soy protein solution that coats any portions of the well walls that do not have bound proteins present. The soy coating prevents further binding of proteins to the walls.
Stone tools that were small enough in size were placed in test tubes and soaked in distilled water to place any adhering proteins into solution. Stone tools that were larger than the test tubes had drops of distilled water randomly placed across their surfaces and stirred with a pipette tip to bring potential proteins into solution. Approximately, 20
m
l of solution extracts from surface material from tools were placed in cups of a microtiter plate for 24 h at room temperature. Again, any proteins present in the solution bind to walls of the wells.Polyclonal antisera raised in rabbits were added to wells for 24 h, allowing them to bind with the matching albumins.Table 2lists the reagents used in the analyses. The antibodies were produced by Low-enstein and Scheuenstuhl and University of California colleagues in their laboratories. The I-125 labeled DARGG was obtained from the Amersham Corporation. The plate was washed with bovine serum albumin and radioactive (I-125 labeled) donkey anti-rabbit gamma globulin (DARGG) was then added to the cups for another 24 h. DARGG binds to any rabbit gamma globulins adhering to wells with the excess being washed out with sterile water. Radioactivity of DARGG is measured by a scintillation counter and converted to percentage uptake of the radioactive antibody. The amount of radioactivity reflects specific antibody to specific antigen binding; however, a minor amount (0.5– 2.0%) of nonspecific DARGG binding occurs in wells in which no extracts and rabbit antisera were added. This is because proteins in solid-phase RIA have a strong affinity for binding to the polystyrene plate wells. To minimize nonspecific DARGG binding, the DARGG solution is made up with a much stronger solution of bovine serum, so that the bovine serum proteins fill nearly all the nonspecific binding sites on the plate well and the DARGG affinity is nearly entirely limited to the rabbit globulin specifically bound to the extracted protein. However, the nonspecific binding is not completely suppressed, so control wells with no rabbit globulin are used so that this nonspecific binding can be subtracted to give the specific binding. Thus, nonspecific binding was subtracted from the total % radioactivity measured in each well.
Each sample was analyzed three times with values from all three test runs averaged. As reagents get older, the ratio of nonspecific to specific binding tends to increase. For the reaction between anti-sera and albumin to be significant, the amount of radioactivity in a well should be at least twice the amount of nonspecific binding. The reaction values represent percent binding of labeled antibody. Highest percentage of binding reflects a significant reaction and is interpreted as an identification of a species. Non-identification (‘‘no
reaction’’) occurs when values are extremely low; we consider significant reactions to be3 (following earlier studies, e.g., Low-enstein et al., 2006). Weak reactions can occur when one of the three test runs yields significant high binding values and subse-quent testing yields little to no reaction values. Total values may be skewed toward the test run with the high binding values and meet significant value criteria though subsequent testing yielded no reactions.
2.3. Amino acid racemization. C:N ratios and
d
13C valuesAmino Acid Racemization results were obtained from Micro-Analytica, Inc. in Miami, Florida. Materials extracted for amino acid analysis were obtained by first grinding off the outer surface from a 5–10 mm spot on the bone fragment surface. The surface grindings were discarded. About 2–4 mg of underlying bone mineral was then extracted and collected for each sample. Each aliquot was hydro-lyzed at 100C for 24 h to peptize proteins and isolate amino acids. Amino acid racemization ratios were then obtained using a 5 micron Hypersil BDS C18 column in a Hewlett Packard 1100 HPLC.
Amino acid racemization ratios, stable
d
13C values, and stable carbon and nitrogen ratios (13C:15N) of selected bone samples were measured to evaluate diagenesis and potential protein loss or denaturation. Diagenetic alteration of protein structure may lead to cross-reactions and false identifications (Tuross et al., 1996). Aspartic (Asp) acid racemization ratios have generally been used to assess the potential for aDNA and protein preservation in an archaeological assemblage (Poinar et al., 1996; for a critique on the usage of racemization to assess aDNA preservation seeFerna´ndez et al., 2009). Ratio values over 0.1 are interpreted as reflecting samples that have been subjected to diagenetic processes and likely will not produce well-preserved proteins.Bone collagen extractions were conducted at the NSF-Arizona AMS by use of a continuous flow system that administers acid– alkali–acid washes (Bronk Ramsey et al., 2004). The final collagen solution was heated at 70C for 24 h to gelatinize, and freeze dried. Collagen extract carbon–nitrogen ratios were measured by a CHN elemental analyzer at the National Science Foundation-Arizona Accelerator Mass Spectrometry (NSF-Arizona AMS) Laboratory and the Environmental Isotope Laboratory at the Department of Geo-sciences, University of Arizona.
d
13C values were calculated at the NSF-Arizona AMS and Beta Analytic, Inc.C:N atomic ratios for well-preserved modern collagen range between 2.8 and 3.5 (Ambrose, 1990).
d
13C values for bison and wapiti (elk) should range between23.0 and19.0&(Matheus et al., 2003). Cariboud
13C values can be expected to fall within the range of arctic terrestrial herbivores (19.0 and17.0&;Coltrain et al., 2004).d
13C values that fall outside of these ranges may indicate contributions of exogenous carbon and/or fractionation during the process of diagenesis and reflects the potential for poor protein preservation (Hedges and Van Klinken, 1992; van Klinken, 1999).3. Results
3.1. Gerstle river site, component 3
within the range of wapiti and bison (23.0 to19.0&) and do not reflect any contributions of exogenous carbon that would be reflected in samples that have undergone higher degree of diagenesis. Amino acid racemization values (D/L Asp) for the Gerstle River samples range between 0.135 and 0.253 (Table 3). In general, these values have been interpreted as reflecting samples where proteins were been denatured due to diagenesis. However, several bison bones approximately the same age (w11,000 cal BP) recovered from Gerstle River yielded sufficiently preserved DNA for analysis (Shapiro et al., 2004), and several bone specimens from Component 3 (n¼12) had high enough collagen yields for normal radiocarbon analysis (Potter et al., n.d.).
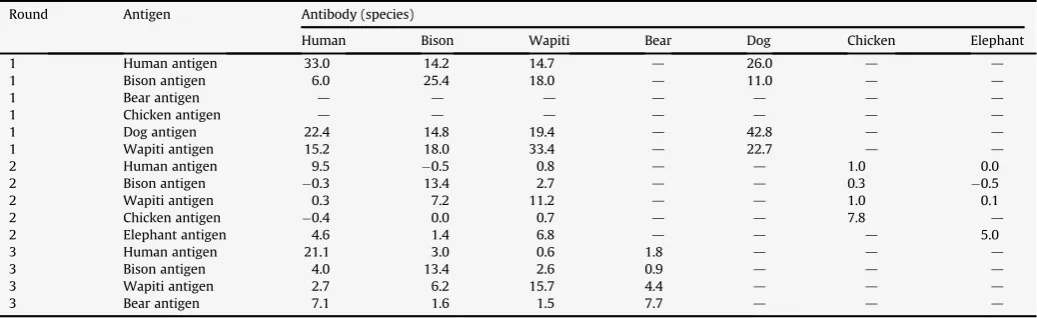
Five of the eight Gerstle River bone samples reacted strongly to antisera (Gerstle #1, 3, 4, and 5) (Table 4). Four of the five reacted to bison antisera, while three of the five reacted to wapiti antisera. Two reacted strongly to both wapiti and bison antisera. The pRIA results were mixed for Gerstle River specimens. Of the bison specimens (#1, 5, 8), one was identified through pRIA as bison (#1, though it also yielded a significant wapiti reaction), one was misidentified as wapiti (#5), and one yielded no significant reaction (#8). Of the wapiti specimens (#2, 4, 6, and 7), one was identified through pRIA as wapiti (#4), one was misidentified as bison (#2), and two yielded no significant reactions (#6 and 7). Two specimens yielded signifi-cant reactions to both wapiti and bison (#1 and 3) suggesting that factors leading to interspecific cross-reactions have yet to be isolated and controlled. The relatively low reaction values (3–6) is signifi-cantly lower than cross-check values (antigen–antibody) which averaged much higher (up to 43) (Table 5). These results suggest that for ancient samples with concomitantly poorer preservation, the minimum values deemed sufficient for significant reactions should be elevated, perhaps to 6. Repeatability of the reaction values is difficult to gauge with only three specimens run twice. These three specimens were analyzed again in order to include species not tested in the first run (e.g., wapiti). The reaction values did vary in Gerstle #1–3, increasing for most human and bison antibody reaction values from the first to the second run, while chicken antibody reaction values remained unchanged. This suggests that subsequent analyt-ical runs may offer new significant reactions.
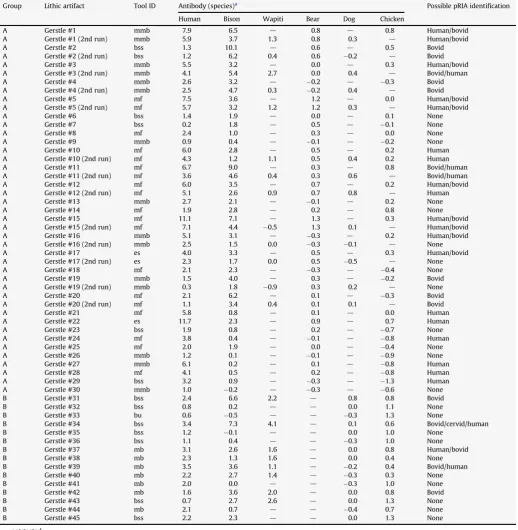
pRIA analysis results for the lithic artifacts are provided inTable 6. Repeatability is evaluated through the 13 lithic samples run twice against human, bison, and bear. All human reactions decreased on the second run (averaging 1.7 less, ranging from 0.1 to 4.0). Eleven of the 13 bison reactions decreased on the second run (averaging 1.9 less). Bear results were more mixed, with generally low reaction values (<1.0). There were shifts from significant reactions to non-significant reactions (1 human reaction, and 4 bear reactions were not replicated during the second run), but there was no instance of a shift from non-significant to significant reactions. This may
indicate fewer organic residues remain after the first analytical run, an opposite situation than the bone results (above). We cannot account for this difference in additional significant reactions in subsequent analytical runs, but more research is necessary to evaluate effects of multiple pRIA analyses on a single specimen.
In order to quantify the potential for human handling to contaminate the lithic specimens, we analyzed the 30 lithics recovered between 1999 and 2003 using no safeguards to prevent human contamination (via sweat, skin cells, etc.) (Group A), and the 15 lithics recovered in 2006 with safeguards in place (Group B). Antibody reaction values were averaged for each artifact. The average human reaction values were significantly different between these groups (averaging 3.62.7 for Group A and 1.90.9 for Group B;t-test,p¼0.018). The average bison reaction values were not significantly different between these groups (averaging 2.52.1 for Group A and 2.22.2 for Group B;t-test,p¼0.530). Wapiti was not compared as only 47% of the artifacts were tested against wapiti antibodies. These results clearly indicate a reduction by almost half of the significant human reactions (27% of Group B compared with 57% of Group A) with the anti-contamination protocols. Several human positive reactions for Group A exceeded 6.0, whereas none of the Group B human positive reactions did, adding further support for the elevation of minimum significant reactions to 6 instead of 3.
We analyzed the lithics to see if there were differences in significant reactions based on artifact type and presumed function. Modified flakes did have larger reactions to bison antibodies (aver-aging 2.72.0) than microblades (averaging 2.11.4), consistent with the hypothesis that modified flakes were used directly in butchery on-site, and discarded expediently, whereas microblades were removed from weapon tips, possibly some distance and time removed from contact with animal tissue (i.e., heavily curated). Boulder spall scrapers were the most variable (averaging 2.82.9) along with the highest reactions to wapiti (averaging 2.31.5), possibly due to the larger surface area of the artifacts, more coarse-grained texture, with more concavities to trap residues (relative to finer grained silicates used for other tool types), and the closer inferred relationship to animal butchery/hide processing, their expedient nature (i.e., they were likely created quickly, used on-site, and quickly discarded on-site without further transport).
There was no difference in bison reaction values between unmodified or modified microblades on average, which may suggest that either (1) microblades were used without retouch or obvious damage, thus yielding similar levels of bison and wapiti residues, or (2) the low levels of reactions (generally <3.0) are artifacts of the pRIA analysis and do not meet the threshold of significant reactions.
There are clear spatial clusters of wapiti and bison at Gerstle River, as well as areas where both taxa overlap (Fig. 1). Gerstle 2,
Table 3
C:N ratios,d13C values, and amino acid racemization results.
Sample Site D/L Asp D/L Ala C:N ratio d13C&
Gerstle-1 Gerstle River 0.154 0.021 3.39 21.5
Gerstle-2 Gerstle River 0.178 0.023 Insufficient collagen 21.7
Gerstle-3 Gerstle River 0.153 0.021 3.51 21.3
Gerstle-4 Gerstle River 0.253 0.078 3.90 Lost
Gerstle-5 Gerstle River 0.141 0.021 4.00 22.0
Gerstle-6 Gerstle River 0.194 0.067 Not run Not run
Gerstle-7 Gerstle River 0.212 0.040 Not run Not run
Gerstle-8 Gerstle River 0.135 0.015 Not run Not run
Croxton-1 Croxton, Loc. J 0.050 0.001 3.20 18.4
Croxton-2 Croxton, Loc. J 0.114 0.005 Not run Not run
Croxton-3 Croxton, Loc. J 0.046 0.001 Not run Not run
Croxton-4 Croxton, Loc. J 0.045 0.001 3.20 18.9
Croxton-5 Croxton, Loc. J 0.045 0.001 3.20 18.4
Ref-1 n/a 0.026 0.010 n/a n/a
a wapiti metatarsal, and one end scraper (#22) are located in an area with exclusively wapiti remains. The end scraper yielded no significant reactions to wapiti antibodies. The remaining lithics are from areas with both bison and wapiti, but many more wapiti elements are present (wapiti NISP¼73, MNI¼5; bison NISP¼33, MNI¼3) (Potter, 2007). The predominance of lithics with bison reactions (n¼18) over wapiti (n¼3) is difficult to reconcile with the spatial patterning of artifacts (Fig. 1) and the overall faunal predominance of wapiti. All three artifacts with significant wapiti reactions had higher reactions to bison. We speculate that the potential for cross-reactions between species and the general low levels of percent uptake of the radioactive antibodies (probably due to diagenetic factors given the pRIA results on bone samples from this site) render precise taxonomic identifications unfeasible.
Cross-reactions appear to be present, given the multiple signifi-cant reactions to human, bison, and wapiti antibodies in the Gerstle bone samples. We explored the possibility that these are related to genetic distance (i.e., if cross-reaction potential is related to genetics, then bison and musk-ox (Family Bovidae)) should have a higher correlation of significant reactions than bison and human (different Orders). This is not the case, the Pearsonrvalues of bison:wapiti (same Order, Artiodactyla) are the highest (r¼0.68), whereas bison:sheep and bison:musk-ox are lower (r¼0.35, 0.39, respec-tively), with bison:human at similar low levels (r¼0.35). Artiodac-tyls, trout, and chicken all yielded similar low levels of correlation.
We believe further inquiry on inter-taxon cross-reactions will be needed to further evaluate the pRIA analysis methods used here.
3.2. Croxton site, loc. J
Five bone samples recovered from Croxton and identified as caribou remains were analyzed. Amino acid racemization ratios for all five samples are 0.1 or less. Stable carbon and nitrogen ratios and stable13C values were measured for three samples. C:N ratios for each sample is 3.2 and
d
13C values that range between18.4 and 18.9&(Table 3). C:N and amino acid racemization ratios generally indicate samples that have excellent preservation for proteins.d
13C values fall within expected values (17& to 19&) of arctic terrestrial herbivores (Coltrain et al., 2004), again, indicating that contributions of exogenous carbon is minimal and/or diagenesis of the organic fractions of the samples is minor.Bone extracts from all five of the Croxton samples strongly reacted to wapiti antisera indicating an origin of proteins from Cervidae family, of which caribou (Rangifer tarandus) belong to. However, strong bovid and bear reactions were present in all samples, and Croxton-4 yielded higher bear reactions than cervid. One extract (Croxton-2), had the highest reaction with bison anti-sera. This sample also had the highest aspartic acid racemization ratio and may indicate some degree of diagenetic alteration leading to a cross-reaction. The remaining Croxton samples had low
Table 4
pRIA results for faunal specimens.
Round Archaeological bone specimen Antibody (species) Morphological identification Possible pRIA identification
Human Bison Wapiti Bear Dog Chicken Elephant
1 Gerstle-1 2.0 5.4 d 1.0 d 1.0 d Bison Bovid
1 Gerstle-1 (2nd run) 3.2 5.4 3.2 0.2 0.0 1.0 d Bison Bovid
1 Gerstle 2 0.7 2.0 d 0.7 d 1.9 d Wapiti None
1 Gerstle 2 (2nd run) 3.0 3.2 2.2 0.7 0.5 1.9 d Wapiti Bovid/human
1 Gerstle 3 0.8 2.3 d 0.3 d 0.6 d VL artiodactyl None
1 Gerstle 3 (2nd run) 2.8 5.7 3.3 0.3 0.6 0.6 d VL artiodactyl Bovid/cervid
2 Gerstle 4 1.6 2.2 2.9 d d 1.3 0.5 Wapiti None
2 Gerstle 5 0.9 0.8 2.7 d d 1.0 0.5 Bison None
2 Gerstle 6 0.1 0.2 0.0 d d 1.0 0.0 Wapiti None
2 Gerstle-7 0.4 0.3 1.6 d d 1.0 0.3 Wapiti None
2 Gerstle 8 1.0 1.1 0.9 d d 1.0 0.5 Bison None
3 Croxton-1 1.2 7.1 9.6 4.5 d d d Caribou Cervid, bovid, bear
3 Croxton-2 1.7 9.4 9.5 1.7 d d d Caribou Cervid, bovid
3 Croxton-3 1.6 5.0 12.1 4.0 d d d Caribou Cervid, bear
3 Croxton-4 1.3 6.4 9.0 9.8 d d d Caribou Bear, cervid, bovid
3 Croxton-5 1.6 4.6 9.8 5.0 d d d Caribou Cervid, bear, bovid
d¼not tested.
Table 5
Cross-check results for antigen and antibodies.
Round Antigen Antibody (species)
Human Bison Wapiti Bear Dog Chicken Elephant
1 Human antigen 33.0 14.2 14.7 d 26.0 d d
1 Bison antigen 6.0 25.4 18.0 d 11.0 d d
1 Bear antigen d d d d d d d
1 Chicken antigen d d d d d d d
1 Dog antigen 22.4 14.8 19.4 d 42.8 d d
1 Wapiti antigen 15.2 18.0 33.4 d 22.7 d d
2 Human antigen 9.5 0.5 0.8 d d 1.0 0.0
2 Bison antigen 0.3 13.4 2.7 d d 0.3 0.5
2 Wapiti antigen 0.3 7.2 11.2 d d 1.0 0.1
2 Chicken antigen 0.4 0.0 0.7 d d 7.8 d
2 Elephant antigen 4.6 1.4 6.8 d d d 5.0
3 Human antigen 21.1 3.0 0.6 1.8 d d d
3 Bison antigen 4.0 13.4 2.6 0.9 d d d
3 Wapiti antigen 2.7 6.2 15.7 4.4 d d d
3 Bear antigen 7.1 1.6 1.5 7.7 d d d
aspartic acid racemization ratios, perhaps indicating weaker diagenetic alterations.
4. Discussion
Preservation of organic portions of animal bones can be affected by a variety of taphonomic and diagenetic factors (for a review see Millard, 2001), including burning, burial, and interaction with soil
chemistry and microbes. Burning did not seem to have significant effects on pRIA results or measures of diagenesis (C:N ratios, amino acid racemization) (Table 3). One of the two unburned Gerstle River bone samples (Gerstle-1) and one exhibiting charring had the among the highest pRIA reaction values, whereas the other unburned Gerstle River bone sample (Gerstle-7) had among the lowest pRIA reaction values. As a whole, however, the amino acid racemization ratios are elevated for the Gersle River samples
Table 6
pRIA results for lithic artifacts (all from Gerstle River).
Group Lithic artifact Tool ID Antibody (species)a Possible pRIA identification
Human Bison Wapiti Bear Dog Chicken
A Gerstle #1 mmb 7.9 6.5 d 0.8 d 0.8 Human/bovid
A Gerstle #1 (2nd run) mmb 5.9 3.7 1.3 0.8 0.3 d Human/bovid
A Gerstle #2 bss 1.3 10.1 d 0.6 d 0.5 Bovid
A Gerstle #2 (2nd run) bss 1.2 6.2 0.4 0.6 0.2 d Bovid
A Gerstle #3 mmb 5.5 3.2 d 0.0 d 0.3 Human/bovid
A Gerstle #3 (2nd run) mmb 4.1 5.4 2.7 0.0 0.4 d Bovid/human
A Gerstle #4 mmb 2.6 3.2 d 0.2 d 0.3 Bovid
A Gerstle #4 (2nd run) mmb 2.5 4.7 0.3 0.2 0.4 d Bovid
A Gerstle #5 mf 7.5 3.6 d 1.2 d 0.0 Human/bovid
A Gerstle #5 (2nd run) mf 5.7 3.2 1.2 1.2 0.3 d Human/bovid
A Gerstle #6 bss 1.4 1.9 d 0.0 d 0.1 None
A Gerstle #7 bss 0.2 1.8 d 0.5 d 0.1 None
A Gerstle #8 mf 2.4 1.0 d 0.3 d 0.0 None
A Gerstle #9 mmb 0.9 0.4 d 0.1 d 0.2 None
A Gerstle #10 mf 6.0 2.8 d 0.5 d 0.2 Human
A Gerstle #10 (2nd run) mf 4.3 1.2 1.1 0.5 0.4 0.2 Human
A Gerstle #11 mf 6.7 9.0 d 0.3 d 0.8 Bovid/human
A Gerstle #11 (2nd run) mf 3.6 4.6 0.4 0.3 0.6 d Bovid/human
A Gerstle #12 mf 6.0 3.5 d 0.7 d 0.2 Human/bovid
A Gerstle #12 (2nd run) mf 5.1 2.6 0.9 0.7 0.8 d Human
A Gerstle #13 mmb 2.7 2.1 d 0.1 d 0.2 None
A Gerstle #14 mf 1.9 2.8 d 0.2 d 0.8 None
A Gerstle #15 mf 11.1 7.1 d 1.3 d 0.3 Human/bovid
A Gerstle #15 (2nd run) mf 7.1 4.4 0.5 1.3 0.1 d Human/bovid
A Gerstle #16 mmb 5.1 3.1 d 0.3 d 0.2 Human/bovid
A Gerstle #16 (2nd run) mmb 2.5 1.5 0.0 0.3 0.1 d None
A Gerstle #17 es 4.0 3.3 d 0.5 d 0.3 Human/bovid
A Gerstle #17 (2nd run) es 2.3 1.7 0.0 0.5 0.5 d None
A Gerstle #18 mf 2.1 2.3 d 0.3 d 0.4 None
A Gerstle #19 mmb 1.5 4.0 d 0.3 d 0.2 Bovid
A Gerstle #19 (2nd run) mmb 0.3 1.8 0.9 0.3 0.2 d None
A Gerstle #20 mf 2.1 6.2 d 0.1 d 0.3 Bovid
A Gerstle #20 (2nd run) mf 1.1 3.4 0.4 0.1 0.1 d Bovid
A Gerstle #21 mf 5.8 0.8 d 0.1 d 0.0 Human
A Gerstle #22 es 11.7 2.3 d 0.9 d 0.7 Human
A Gerstle #23 bss 1.9 0.8 d 0.2 d 0.7 None
A Gerstle #24 mf 3.8 0.4 d 0.1 d 0.8 Human
A Gerstle #25 mf 2.0 1.9 d 0.0 d 0.4 None
A Gerstle #26 mmb 1.2 0.1 d 0.1 d 0.9 None
A Gerstle #27 mmb 6.1 0.2 d 0.1 d 0.8 Human
A Gerstle #28 mf 4.1 0.5 d 0.2 d 0.8 Human
A Gerstle #29 bss 3.2 0.9 d 0.3 d 1.3 Human
A Gerstle #30 mmb 1.0 0.2 d 0.3 d 0.6 None
B Gerstle #31 bss 2.4 6.6 2.2 d 0.8 0.8 Bovid
B Gerstle #32 bss 0.8 0.2 d d 0.0 1.1 None
B Gerstle #33 bu 0.6 0.5 d d 0.3 1.3 None
B Gerstle #34 bss 3.4 7.3 4.1 d 0.1 0.6 Bovid/cervid/human
B Gerstle #35 bss 1.2 0.1 d d 0.0 1.0 None
B Gerstle #36 bss 1.1 0.4 d d 0.3 1.0 None
B Gerstle #37 mb 3.1 2.6 1.6 d 0.0 0.8 Human/bovid
B Gerstle #38 mb 2.3 1.3 1.6 d 0.0 0.4 None
B Gerstle #39 mb 3.5 3.6 1.1 d 0.2 0.4 Bovid/human
B Gerstle #40 mb 2.2 2.7 1.4 d 0.3 0.3 None
B Gerstle #41 mb 2.0 0.0 d d 0.3 1.0 None
B Gerstle #42 mb 1.6 3.6 2.0 d 0.0 0.8 Bovid
B Gerstle #43 bss 0.7 2.7 2.6 d 0.0 1.3 None
B Gerstle #44 mb 2.1 0.7 d d 0.4 0.7 None
B Gerstle #45 bss 2.2 2.3 d d 0.0 1.3 None
d¼not tested.
(averaging 0.1780.040 for D/L Asp and 0.0360.024 for D/L Ala), several times higher than for the Croxton bones (Table 7). This, along with the depleted
d
13C/12C values and relatively higher C:N ratios indicates that the Gerstle River specimens were affected more by diagenetic alteration of collagen. Using the Hemstix and dot blot assay methods,Manning (1994) suggests that trace amounts of foreign substances could be detected resulting in false-positive identifications that do not relate to an archaeological event. This contamination would be amplified when comparing the results of the detection of foreign substances and those derived from ancient residues that have already low detection values. pRIA is a highly sensitive technique that is able to detect to the 0.001m
g of antigen and may be more susceptible to false-positive identifications as the indigenous protein has degraded and sample size has decreased to a very low detection level. The Gerstle River samples appear to have this potential contamination (with human reactions in 3 of the 8 samples, 38%). In bone material from more recent contexts (Croxton) with less inferred diagenetic alteration, this contamination is not present (i.e., no significant human reaction values,Table 4). In an earlier study (Lowenstein et al., 2006: 183), the pRIA reaction values for modern bone ranged from 10 to 23, generally>5 to 10% greater than the second greatest reaction values. The fossil human bone in that study yielded a reaction of 3 for human and 2 for bison; these are lower absolute values, similar to those reported for ancient bones (Gerstle River samples) in this study.Diagenetic factors leading to the loss of collagen include dete-rioration of the microstructural elements of bone by microbial attack or chemical dissolution leading to the increased exposure of organic components, accelerating hydrolysis of organic compo-nents, and subjection to extreme high temperatures such as partial combustion in a hearth. To explore the effects of these diagenetic factors on the pRIA analysis’ efficacy, we summarize data from Tables 4 and 5, providing averages of various measures of diage-netic alteration as well as the highest reaction values and the difference between the highest and next highest reaction values for each taphonomic group (Early Holocene, Late Holocene, recent, and modern cross-checks) (Table 7). We discuss the pRIA bone results in these contexts.
Of the 8 early Holocene bone specimens (w10,100 cal BP), the pRIA analyses yielded correct identifications for 1 (13%), a misidentification for 1 (13%), a mixed identification of bison/ wapiti for 1 (13%) and no significant reactions for the remaining specimens (63%). While the sample size is small, these results collectively contrast with those of the late Holocene samples. The pRIA analyses yielded correct identifications (based on highest reactions) for four of the five late Holocene bone specimens (1650-790 cal BP), though two of the results had nearly equally high cross-reactions (Croxton-2, 4), and all had cross-cross-reactions (>3) with multiple non-human taxa.
The cross-checks summarized from Table 5 quantify the effects of diagenesis on the reaction values (Table 7). The highest values for modern samples are similar to the late Holocene samples (around 10), but the difference between the highest and next highest reaction values are much larger for modern samples
compared with late Holocene bone samples. The early Holocene bones show even more decreases in the average highest reaction values (around 3) and slight differences between the highest and next highest reaction values. While Gerstle River and Croxton sites are in many ways unique, the diagenetic effects identified in these specimens can be useful for evaluating the efficacy of pRIA when dealing with ancient organic remains in archaeological contexts.
An earlier study using modern experimental samples yielded generally good results (Lowenstein et al., 2006), and the results from the late Holocene Croxton bone samples reported here are consistent with that study, showing that the pRIA analysis method can be effective for modern and recent bone with little diagenetic alteration, though there is still potential for non-human cross-reactions. We recommend multiple samples from the same archaeological contexts be used to assess variability in the cross-reactions and mitigate false-positive identifications.
The results from Gerstle River reported here indicate that the method appears ineffective when dealing with ancient bones with some diagenetic alteration. The potential for inter-taxonomic cross-reactions appears elevated when the overall highest reaction values are very low. For this reason, quantified above, we recommend that the minimum sufficient reaction value of significance be raised from 3 to 6. This appears to mitigate the problems encountered with diagenetically altered bone, and also provides a more appro-priate sensitivity level for the test.
In conclusion, we consider the pRIA analysis described here and elsewhere (Lowenstein et al., 2006) to be a valuable forensic tool for modern and recent samples. However, our results indicate that the test is at present ineffective when dealing with more ancient samples, in the form of both false positives and false negatives. This is largely due to the high sensitivity of the pRIA technique. More studies are needed to control for various levels of diagenetic alteration, to calibrate the analysis and identify its limitations. The high levels of inter-taxonomic cross-reactions should be more fully examined in future studies to explore their causes and develop strategies to effectively mitigate their effects.
Acknowledgments
We thank the following individuals for their contributions to this project: Darden Hood, MicroAnalytica, LLC, analyzed the amino acids, Carol Gelvin-Reymiller identified the caribou bone samples from Croxton, and the NSF-Arizona AMS Facility and Environmental Isotope Laboratory at the Department of Geosciences, University of Arizona, provided stable carbon and nitrogen ratio measurements. Anonymous reviewers provided useful comments. We also thank researchers and institutions that have supported this research, particularly S. Craig Gerlach of the Cross-Cultural Studies Depart-ment at the University of Alaska Fairbanks for initiating earlier blind studies that in part stimulated this research and Mike Kunz of the Bureau of Land Management for supporting the 2000 Croxton excavations.
Table 7
Summary data on bone samples (all runs averaged for each sample, then total averages taken for each age group).
Age N D/L Asp D/L Ala C:N d13C Highest reaction
value
Difference between highest and next highest reaction value
w10,100 cal BP 8 (n¼4 for C:N andd13C) 0.178 0.036 3.7 21.6 2.7 0.9
1650-790 cal BP 5 (n¼3 for C:N andd13C) 0.060 0.002 3.2 18.6 10.2 3.0
Recent bone 2 0.029 0.050 n/a n/a n/a n/a
Modern cross-checks (antigen and antibodies)a 13 n/a n/a n/a n/a 9.5 8.7
References
Ambrose, S.H., 1990. Preparation and characterization of bone and tooth collagen for isotopic analysis. Journal of Archaeological Science 17, 431–451.
Barnard, H., Ambrose, S.H., Beehr, D.E., Forster, M.D., Lanehart, R.E., Malainey, M.E., Parr, R.E., Rider, M., Solazzo, C., Yohe II, R.M., 2007. Mixed results of seven methods for organic residue analysis applied to one vessel with the residue of a known foodstuff. Journal of Archaeological Science 34, 28–37.
Bourget, S., Newman, M.E., 1998. A Toast to the ancestors: ritual warfare and sacrificial blood in Moche culture. Baessler-Archiv 46, 85–106.
Bronk Ramsey, C., Higham, T., Bowles, A., Hedges, R., 2004. Improvements to the pretreatment of bone at Oxford. Radiocarbon 46 (1), 155–163.
Cattaneo, C., Gelsthorpe, K., Phillips, R., Sokal, R.J., 1993. Blood residues on stone tools: indoor and outdoor experiments. World Archaeology 25, 29–43. Child, A.M., Pollard, A.M., 1992. A review of the applications of immunochemistry to
archaeological bone. Journal of Archaeological Science 19, 39–47.
Coltrain, J.B., Hayes, M.G., O’Rourke, D.H., 2004. Sealing, whaling and caribou: the skeletal isotope chemistry of Eastern Arctic foragers. Journal of Archaeological Science 31 (1), 39–57.
Dixon, E.J., Marlar, R., 1997. A new discovery at the Folsom type site. Plains Anthropologist 42, 371–374.
Downs, E.F., Lowenstein, J.M., 1995. Identification of archaeological proteins: a cautionary note. Journal of Archaeological Science 22, 11–16.
Eisele, J.A., Fowler, D.D., Haynes, G., Lewis, R.A., 1995. Survival and detection of blood residues on stone tools. Journal of Archaeological Science 69, 36–46. Ferna´ndez, E., Ortiz, J.E., Pe´rez- Pe´rez, A., Prats, E., Turbo´n, D., Torres, T.,
Arroyo-Pardo, E., 2009. Aspartic acid racemization variability in ancient human remains: implications in the prediction of ancient DNA recovery. Journal of Archaeological Science 36, 965–972.
Gerlach, S.C., 1989. Models exploitation, butchery and processing at the Croxton site, Tukuto Lake, Alaska. Ph.D. dissertation, Department of Anthropology, Brown University, Providence.
Gerlach, S.C., Newman, M.E., Knell, E.J., Hall Jr., E.S., 1996. Blood protein residues on lithic artifacts from two archaeological sites in the De Long Mountains, Northwestern Alaska. Arctic 49, 1–10.
Gurfinkel, D.M., Franklin, U.M., 1988. A study of the feasibility of detecting blood residue on artifacts. Journal of Archaeological Science 15, 83–97.
Hedges, R.E.M., Van Klinken, G.J., 1992. A review of current approaches in the pretreatment of bone for radiocarbon dating by AMS. Radiocarbon 34 (3), 279–291.
Hyland, D.C., Tersak, J.M., Adovasio, J.M., Siegel, M.I., 1990. Identification of the species of origin of residual blood on lithic material. American Antiquity 55, 104–112.
van Klinken, G.J., 1999. Bone collagen quality indicators for paleodietary and radiocarbon measurements. Journal of Archaeological Science 26, 687–695. Kooyman, B., Newman, M.E., Ceri, H., 1992. Verifying reliability of blood residue
analysis on archaeological tools. Journal of Archaeological Science 19, 265–270. Kooyman, B., Newman, M.E., Cluney, C., Lobb, M., Tolman, S., McNeil, P., Hills, L.V., 2001. Identification of horse exploitation by Clovis hunters based on protein analysis. American Antiquity 66, 686–691.
Kooyman, B., Hills, L.V., Tolman, S., 2006. Late Pleistocene horse hunting at the Wally’s Beach site (DhPg-8), Canada. American Antiquity 71, 101–121. Leach, J.D., Mauldin, R.P., 1995. Additional comments on blood residue analysis in
archaeology. Antiquity 69, 1020–1022.
Lowenstein, J.M., Reuther, J.D., Hood, D.G., Scheuenstuhl, G., Gerlach, S.C., Ubelaker, D.H., 2006. Identification of animal species by protein radioimmu-noassay of bone fragments and bloodstained stone tools. Forensic Science International 159, 182–188.
Loy, T.H., Dixon, E.J., 1998. Blood residues on fluted points from eastern Beringia. American Antiquity 63, 21–46.
Manning, A.P., 1994. A cautionary note on the use of Hemastix and Dot-blot assays for the detection and confirmation of archaeological blood residues. Journal of Archaeological Science 21, 159–162.
Marlar, R.A., Puseman, K., Cummings, L.S., 1995. Protein residue analysis of archaeological materials: comments on criticisms and methods. Southwestern Lore 61 (2), 27–37.
Marlar, R.A., Banks, L.L., Billman, B.R., Lambert, P.M., Marlar, J.E., 2000. Biochemical evidence of cannibalism at a prehistoric Puebloan site in southwestern Colo-rado. Nature 407, 74–78.
Matheus, P., Guthrie, R. D., Kunz, M. L., 2003. Isotope ecology of late Quaternary megafauna in Eastern Beringia. Storer, J. (Ed.), Abstracts for the 3rd Interna-tional Mammoth Conference, Yukon Territory, Canada, May 2003. Abstracts Available from:<http://www.yukonmuseums.ca/mammoth/>.
Mauldin, R.P., Leach, J.D., Amick, D.S., 1995. On the identification of blood residues on Paleoindian artifacts. Current Research in the Pleistocene 12, 85–87. Millard, A., 2001. The deterioration of bone. In: Brothwell, D.R., Pollard, A.M. (Eds.),
Handbook of Archaeological Sciences. John Wiley & Sons, Ltd., Chichester, England, pp. 637–647.
Newman, M.E., Ceri, H., Kooyman, B., 1996. The use of immunological techniques in the analysis of archaeological materials – a response to Eisele; with report of studies at head-smashed-in buffalo jump. Antiquity 70, 677–682.
Newman, M.E., Byrne, G., Ceri, H., Dimnik, L.S., Bridge, P.J., 1998. Immunological and DNA analysis of blood residues from a surgeon’s kit used in the American Civil War. Journal of Archaeological Science 25, 553–557.
Poinar, H.N., Ho¨ss, M., Bada, J.L., Pa¨a¨bo, S., 1996. Amino acid racemization and the preservation of ancient DNA. Science 272, 864–866.
Potter, B. A., 2005. Site structure and organization in central Alaska: archaeological investigations at the Gerstle River site. PhD dissertation, University of Alaska Fairbanks.
Potter, B.A., 2007. Models of faunal processing and economy in early Holocene Interior Alaska. Environmental Archaeology 12 (1), 3–23.
Potter, B.A., 2008. Radiocarbon chronology of central Alaska: technological conti-nuity and economic change. Radiocarbon 50 (2), 181–204.
Potter, B.A., J. D. Reuther, G. Hodgens, S. LaMotta, n.d. Radiocarbon analysis of bone and charcoal in high-resolution archaeological contexts. Manuscript in possession of the authors, University of Alaska, Fairbanks.
Reuther, J. D., 2003. An analysis of radiocarbon assays produced by three labora-tories: a case study on the Croxton site, locality J, northern Alaska. Master’s thesis, Department of Anthropology, University of Alaska, Fairbanks. Reuther, J.D., Gerlach, S.C., 2006. CIEP analysis of an Alaskan archaeological
conundrum. Current Research in the Pleistocene 23, 138–140.
Reuther, J.D., Lowenstein, J.M., Gerlach, S.C., Hood, D., Scheuenstuhl, G., Ubelaker, D.H., 2006. The use of an improved pRIA technique in the identifi-cation of protein residues. Journal of Archaeological Science 33, 531–537. Seeman, M.F., Nilsson, N.E., Summers, G.L., Morris, L.L., Barans, P.J., Dowd, E.,
Newman, M.E., 2008. Evaluating protein residues on Gainey phase Paleoindian stone tools. Journal of Archaeological Science 35, 2742–2750.
Shanks, O.C., Kornfeld, M., Hawk, D.D., 1999. Protein analysis of Bugas-Holding tools: new trends in immunological studies. Journal of Archaeological Science 26, 1183–1191.
Shapiro, B., Drummond, A.J., Rambaut, A., Wilson, M.C., Matheus, P.E., Sher, A.V., Pybus, O.G., Gilbert, M.T.P., Barnes, I., BinLaden, J., Willerslev, E., Hansen, A.J., Baryshnikov, G.F., Burns, J.A., Davydov, S., Driver, J.C., Froese, D.G., Harington, C.R., Keddie, G., Kosintsev, P., Kunz, M.L., Martin, L.D., Stephenson, R.O., Storer, J., Tedford, R., ZimovCooper, A., 2004. Rise and fall of the Beringian Steppe Bison. Science 306, 1561–1565.
Smith, P.R., Wilson, M.T., 1992. Blood residues on ancient tool surfaces: a cautionary note. Journal of Archaeological Science 19, 237–241.
Smith, P.R., Wilson, M.T., 2001. Blood residues in archaeology. In: Brothwell, D.R., Pollard, A.M. (Eds.), Handbook of Archaeological Sciences. John Wiley & Sons, Ltd., Chichester, England, pp. 313–322.
Tuross, N., Barnes, I., Potts, R., 1996. Protein identification of blood residues on experimental stone tools. Journal of Archaeological Science 23, 289–296. Ubelaker, D.H., Lowenstein, J.M., Hood, D.G., 2004. Use of solid-phase