R E V I E W
Open Access
Microbial modulation of plant ethylene
signaling: ecological and evolutionary
consequences
Mohammadhossein Ravanbakhsh
1, Rashmi Sasidharan
2, Laurentius A. C. J. Voesenek
2, George A. Kowalchuk
1and Alexandre Jousset
1*Abstract
The plant hormone ethylene is one of the central regulators of plant development and stress resistance. Optimal
ethylene signaling is essential for plant fitness and is under strong selection pressure. Plants upregulate ethylene
production in response to stress, and this hormone triggers defense mechanisms. Due to the pleiotropic effects of
ethylene, adjusting stress responses to maximize resistance, while minimizing costs, is a central determinant of plant
fitness. Ethylene signaling is influenced by the plant-associated microbiome. We therefore argue that the regulation,
physiology, and evolution of the ethylene signaling can best be viewed as the interactive result of plant genotype
and associated microbiota. In this article, we summarize the current knowledge on ethylene signaling and
recapitulate the multiple ways microorganisms interfere with it. We present ethylene signaling as a model system
for holobiont-level evolution of plant phenotype: this cascade is tractable, extremely well studied from both a plant
and a microbial perspective, and regulates fundamental components of plant life history. We finally discuss the
potential impacts of ethylene modulation microorganisms on plant ecology and evolution. We assert that ethylene
signaling cannot be fully appreciated without considering microbiota as integral regulatory actors, and we more
generally suggest that plant ecophysiology and evolution can only be fully understood in the light of
plant-microbiome interactions.
Keywords:
Evolution, Holobiont, Plant, Phenotype, Physiology, Ethylene, Microbiota, Microbiome, ACC deaminase
Background
Environmental stress and plant fitness
Plants are constantly facing a range of different
environ-mental stressors linked for instance to temperature,
water availability, presence of toxic minerals, or
patho-gens. Stress can be permanent, for instance, when a
plant lives outside its ecological optimum, or acute
dur-ing climatic extremes such as drought and flooddur-ing
waves. Environmental stress has an important effect on
plant fitness and elicits specific adaptations [
1
]. Plants
have evolved a range of physiological and morphological
responses to stressors, allowing them to cope with the
prevailing environmental conditions. Although these
re-sponses vary widely, they all share one characteristic:
they all come at a cost to the plant, diverting resources
from growth and reproduction, and causing negative
side effects that may have consequences on other traits
that can result in indirect fitness costs. Optimizing the
relative investment into stress response and other life
history traits is thus essential to maximize fitness [
2
].
Due to the variability of stressors and their interactive
effects with plant genotype, regulating stress response is
a complex task with several possible optima. Further,
adaptation to one specific set of environmental
condi-tions may negatively affect plant fitness under other
con-ditions [
3
]. Plant transpiration illustrates this dilemma
well: stomatal closure, a typical plant response to
drought, reduces water loss, but comes at a cost of lower
photosynthesis, gas exchange and sap flow. Given that
stomata are also an entry point for several pathogens [
4
],
the optimal aperture will be a function of several
* Correspondence:[email protected]
1Ecology and Biodiversity, Institute of Environmental Biology, Utrecht
University, 3584 CH Utrecht, The Netherlands
Full list of author information is available at the end of the article
parameters including water availability, plant sensitivity
to desiccation, and presence of pathogens [
5
].
To optimize fitness, stress responses must be therefore
carefully adjusted. This implies that plants need to
per-ceive different stressors, process the signals, and trigger
an optimal stress response, thereby maximizing
resist-ance while minimizing costs and side effects. Signal
inte-gration in plants is generally achieved by alterations in
hormonal balance. Hormones such as ethylene, auxin, or
jasmonic acid interactively shape the relative investment
of plants into growth, reproduction, and stress defense
[
6
]. Hormone concentrations are dictated by the
com-bined action of the plant
’
s own regulatory pathways as
well as the activities of its associated microbiota.
Hormonal regulation therefore offers an excellent model
to approach plant physiology, ecology, and evolution
from a holobiont perspective in which plants and
mi-crobes form a coherent unit of selection [
7
].
In this review, we approach ethylene, a central plant
hormone regulating the balance between growth and
stress tolerance, from a holobiont perspective. We first
briefly, summarize the importance of ethylene for stress
tolerance and other life history traits. We then go on to
provide an overview of how plants and their associated
microbiota jointly shape hormonal balances, thereby
shifting plant response toward or away from adaptation
to specific situations.
Regulation of stress response by plants
Role of ethylene
Ethylene is a central plant hormone regulating several
aspects of plant growth and development, throughout
the whole plant life cycle, from germination to
senes-cence [
8
]. In addition, this hormone is essential to
regu-late stress responses and confer stress tolerance [
9
,
10
].
Stress results in increased levels of ethylene in plants.
Stress-derived ethylene is a signal triggering adaptive
re-sponses and influences other hormonal signaling
path-ways [
11
]. Due to the multiple effects of ethylene on
plant phenotype, increased ethylene levels will induce a
range of pleiotropic effects, such as growth inhibition
and late flowering [
12
], in addition to the target response
[
13
]. Precisely controlling cellular ethylene levels is thus
a key aspect of plant physiology [
13
].
Ethylene production, signal transduction, and response
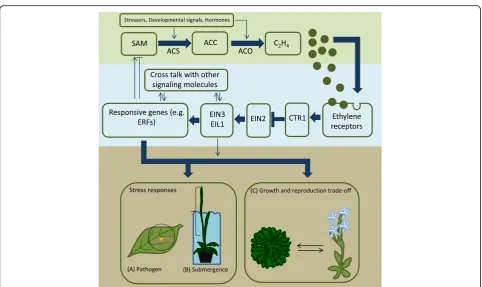
An important step in ethylene production is the
synthe-sis of its precursor ACC (1-aminocyclopropane-1
car-boxylic acid) by ACC synthase (ACS) enzyme (Fig.
1
).
Upon stress detection, ACS mediates the synthesis of
the ethylene precursor ACC, which is transformed to
ethylene by the enzyme ACC oxidase (ACO; Fig.
1
).
Both
ACS
and
ACO
form large multigene families in
plants and different members can be regulated by
different internal and external stimuli [
14
–
16
]. Ethylene
binding to its receptors triggers the expression of
down-stream response genes. In the absence of ethylene, the
ethylene receptors activate CTR1 which is a negative
regulator of ethylene signalling. Ethylene binding
inacti-vates the receptors and therefore CTR1. This
conse-quently relieves the inhibition of EIN2, a positive
regulator of ethylene signaling. Downstream of EIN2 are
the transcription factors EIN3 (ethylene-insensitive 3)
and its homolog EIL1 (ethylene-insensitive 3-like 1) that
are primary mediators of the transcriptional responses
to ethylene [
17
,
18
]. These transcription factors will
in-crease the expression of ethylene responsive
transcrip-tion factors (ERFs) [
19
], resulting in ethylene-mediated
stress responses in plants [
20
]. ERF-regulated traits
in-clude activation of plant immunity [
21
,
22
], metabolic
and morphological adaptations to flooding [
9
,
23
],
ex-pression of systems for scavenging reactive oxygen
spe-cies and modification of enzymatic activity under heavy
metal and salinity stress conditions [
24
–
26
]. The type of
ethylene-mediated response is highly variable, as
dis-cussed below in
“
ethylene variation in plants.
”
Ethylene varies as a function of stress type and intensity
Ethylene production depends on the intensity and
dur-ation of stress periods. For instance, different levels of
heavy metal [
25
,
27
] or different dehydration rates [
28
]
differentially modulate ethylene biosynthesis and
signal-ing. In another example, low-levels of stress stimulate
ethylene production, while high levels may decrease it
[
28
,
29
], either as part of a targeted stress response or as
the result of impaired plant metabolism.
Ethylene response from ecological and agricultural
perspective
Plants are continually confronted with variable
environ-mental conditions, to which they must respond and
adapt. Ethylene has a central role in plant survival and
adaptation
in
dynamic
environments.
life history traits to reach the best possible phenotype
for survival in prevailing ambient conditions, thereby
maximizing reproductive fitness. Variation in ethylene
levels might be due to selection on the plant genetic
material or, as we will discuss in the next section, on
the microorganisms that co-regulate ethylene (Fig.
2
).
Ethylene variation in plants
Variability in ethylene-based stress responses across plant
species
The core ethylene transduction cascade is highly
con-served in plants, and a wide range of plants use ethylene
as a regulator of their stress responses. However, plants
vary greatly in how ethylene impacts stress perception,
transduction, and the final response. Plants evolved in a
certain environment have adapted ethylene signaling to
the stresses typical for that environment or even
dropped it, if not useful. For instance, plants living in
flood-prone or riparian areas, including rice and
Rumex
palustris
, use flooding-induced accumulation of ethylene
to trigger important adaptive responses [
31
]. In contrast,
some aquatic plants adapted to a permanently
sub-merged lifestyle and lost many genes involved in
ethyl-ene signaling [
32
,
33
]. Contrasting ethylene-mediated
responses are even seen in closely related plant species.
For instance, different species of
Rumex
sp. or different
varieties of rice all use ethylene as a flooding signal to
trigger adaptive responses, yet the responses itself are
highly variable, ranging from compensatory growth to
complete quiescence [
31
,
34
].
Variability in stress perception and ethylene signal
transduction
Both
ACS
and
ACO
ethylene biosynthetic genes are
encoded by large multigene families. These genes are
organ-specific and are differentially regulated by
differ-ent environmdiffer-ental signals [
15
,
35
,
36
]. The size of these
multigene families can vary between plant species and
could link to the variation in ethylene-mediated stress
perception. For instance, apple harbors 19
ACS
genes as
compared to 12
ACS
genes in
Arabidopsis thaliana
and
9 in tomato [
36
–
38
]. Deletion of one single gene,
ACS6
,
Fig. 1Overview of the pathways linked to ethylene production (top panel), signal transduction (central panel), and response (bottom panel). Ethylene concentration determines plant resource allocation into growth, reproduction, and stress response [13]. The thick arrows show the main ethylene cascade, and the thin ones point to possible interaction with external and internal stimuli. We illustrate plant response with three well-investigated ethylene-dependent phenotypic adaptations.aEthylene coordinates plant response against pathogens, such as hypersensitive response, preventing pathogen spread [20].bEthylene accumulation triggers escape strategy involving accelerated shoot growth in submerged plants, allowing them to regain atmospheric contact [82].cGrowth-reproduction tradeoffs: higher ethylene causes plants to invest more resources into seed production under harsh conditions that may compromise vegetative stage survival. SAMS-adenosylmethionine, ACC 1-aminocyclopropane-1-carboxylic acid, ACS ACC synthase, ACO ACC oxidase, C2H4 plant hormone ethylene, CTR1 constitutive triple response 1, EIN2 ethylene-insensitive protein 2,resulted for instance in a 85
–
90% reduction in ethylene
production in maize [
39
]. Variation in the ethylene
re-sponse can also occur at the level of ethylene perception
across different plant genotypes linked to changes in
re-ceptor affinity, expression pattern, and/or turn-over [
40
].
Furthermore, knocking out
EIN3
showed opposite
ef-fects on salinity tolerance in
Arabidopsis
and rice [
41
].
Ethylene signaling and response are highly dependent on
plant genotype [
42
], organ, growth stage [
43
], and
asso-ciated microbiota.
Microbiota
Importance of microbiota as co-regulator of stress
response
Plants are associated with a complex microbiome,
in-cluding bacteria, fungi, and protists that have impact on
diverse aspects of plant growth, health, and evolution
[
44
]. Plant-associated microbiota can be either vertically
transmitted, as is the case for endophytes that live within
plant tissues, or horizontally for instance by recruiting
microbiota to the rhizosphere from the surrounding soil
species pool. Microbiota form an integral part of a
plant
’
s immune system, metabolism, and hormonal
bal-ance [
45
]. They directly alleviate stress, for instance, by
producing protective compounds that enhance drought
resistance [
34
,
46
], by degrading organic pollutants [
47
],
or by chelating heavy metals [
48
]. Plant-associated
mi-crobes can also fine-tune hormonal balance and
physi-ology by modulated plant hormone levels and the
pathways they steer. In the case of ethylene, several
pos-sible mechanisms have been described by which
micro-organisms can affect plant hormonal levels. Below, we
examine the mechanisms by which the plant-microbe
di-alog determines ethylene-mediated plant responses as
the basis for a more general model on holobiont-level
regulation of plant hormonal balance.
Ethylene modulation as a holobiome process
Ethylene signaling forms a perfect example of a
holobiont-level physiological cascade. From the holobiont
perspec-tive, plant physiology is controlled by a combination of
traits encoded in the host genome as well as its associated
microbes, which collectively form the holobiont [
7
]. This
association offers a broader genetic pool than the plant
alone: ethylene-modulating microbes could increase the
reservoir of genetic information linked to ethylene
signal-ing, enabling a greater plant phenotypic plasticity in
response to stressors. The microbiota can (1) impact
plant-perceived stress, (2) co-regulate ethylene which
af-fects plant fitness, and (3) perceive ethylene, potentially
responding to it.
Plant and ethylene-modulating microbes as unit of selection
Ethylene levels are a strong determinant of fitness in
dy-namic environments. Given that plants and microbes
work in concert to modulate ethylene-mediated
re-sponses, the holobiont level of selection is the most
ap-propriate: the ethylene cascade provides an important
link between the host and its associated microbes, and
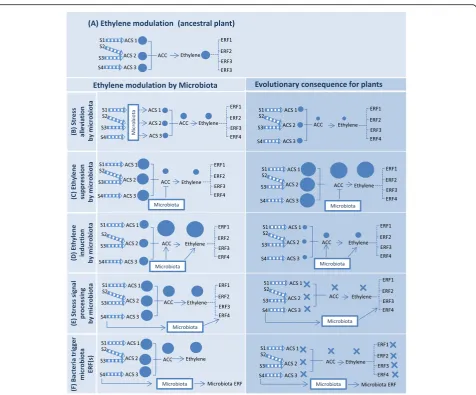
Fig. 2A holobiont-level regulation of ethylene signaling and plant stress response. Ethylene pathway in plants (green area).forms an integrated biological entity [
44
]. This
inter-action even has the potential to be evolutionarily stable:
plants rely on microbes to optimize their fitness and
mi-crobes directly benefit from a more vigorous host that
may provide more nutrients and energy. As plants can
select associated microbes on the basis of the functions
they perform, mutualistic interactions may persist across
generations.
Ethylene modulation by microorganisms:
evolutionary impact on plants
a) Reduction of stress perception by microorganisms
Microorganisms may contribute to plant stress
toler-ance in an ethylene-independent way by providing
pro-tection mechanisms expressed outside of the host plant.
For instance, plant-associated microbiota may reduce
the intensity of stress experienced by the plant by
de-toxifying chemicals or providing protective substances
against desiccation [
46
–
48
]. From an evolutionary
per-spective, a plant
’
s reliance on the microbiome to reduce
stressors may lead to a reduced ability of the plant to
re-spond to the acute stressors (Fig.
3b
), a task delegated to
the associated microbiota.
b) Alteration of ethylene level by microorganisms
Microbes can potentially influence all regulatory steps
of the ethylene pathway (Fig.
2
). The most direct way of
acting on ethylene signaling is to either directly produce
or degrade ethylene. Several plant-associated microbes
can increase plant ethylene levels by directly
synthesiz-ing ethylene or inducsynthesiz-ing plant ACS activity [
49
–
52
].
Ethylene production by microbes was first reported in
the pathogen
Ralstonia solanacearum
, which, among
other symptoms, induces banana premature ripening
[
52
]. Microbial ethylene production was later mainly
in-vestigated in relation to pathogenic bacteria [
50
,
51
].
However, biosynthetic pathway studies [
53
] and the
examination of available bacterial genomes have revealed
that the relevant genes and pathways can be found
across a wide range of microorganisms [
54
,
55
]. For
in-stance, more than one third of all cultivable soil bacteria
can produce ethylene via different pathways [
53
].
Phylo-genetic studies of ethylene-forming enzymes show that
multiple ethylene-producing pathways have evolved
in-dependently and later spread between bacterial phyla by
horizontal gene transfer [
53
,
56
]. Ethylene production by
microbes may have deep effects on plant physiology and
life history, as demonstrated by the accelerated fruit
ripening in plants inoculated with
Escherichia coli
engi-neered to produce ACC oxidase [
57
]. Rhizosphere
mi-crobes can further increase ethylene indirectly by
secreting auxin [
58
,
59
] and cytokinin [
58
,
60
], two
hor-mones that upregulate the expression of ACS-coding
genes [
61
,
62
].
Microbioorganisms can also decrease ethylene levels,
for instance, by producing ACC deaminase. This
en-zyme degrades ACC, ultimately leading to lower plant
ethylene concentrations. ACC deaminase can be found
in both commensal [
63
] and pathogenic microbes [
64
].
ACC deaminase genes are widespread in bacteria,
fungi, and members of stramenopiles [
65
]. ACC
deami-nase genes of bacteria and fungi shared high sequence
identity [
65
], pointing to a single evolutionary origin
and frequent horizontal transfer of this gene in
bac-teria and fungi [
65
–
67
]. The reduction of ethylene
levels caused by ACC-deaminase producing microbes
is of the same magnitude as the one resulting from
knocking out
ACS
genes [
39
,
68
]. In contrast to
com-monly held assumptions, ACC deaminase-producing
microbes are not necessarily good and the effects of
ACC deaminase on plant physiology and plant growth
greatly depend on the interactive effects of plant
geno-type [
69
,
70
] and the environment [
68
,
71
]. For
in-stance, root growth reduction by ethylene is a common
adaptation to avoid salt and pollutants [
72
]. Alleviating
this inhibition may bring a short-term increase in root
growth, but may ultimately be deleterious for the
plant.
Co-evolution of plants with bacteria that increase
or inhibit ethylene levels may have various
conse-quences: co-evolution of plants with microbes
in-creasing ethylene levels may cause the plant to
reduce ethylene production in order to maintain
homeostasis. This could result in a lower ability to
produce ethylene, a higher sensitivity to stressors
and a dependency on microbial ethylene production
(Fig.
3d
).
In
contrast,
plants
associated
with
ethylene-reducing microorganisms such as ACC
de-aminase producers may need to produce more ACC
to compensate for microbial degradation (Fig.
3c
).
Thus, plants may evolve a higher expression of
ACS
genes, which can not only allow a wide range of
re-sponses, but may also lead to overreactions to stress
without modulation by the associated microbiome.
Microbial alteration of plant ethylene response and
intertwined signaling
In
addition
to
direct
manipulation
of
ethylene
ethylene-regulated traits, communicating stress perception to
plants, and monitoring plant stress status. As ethylene
sensitivity in plant-associated microbiota is
wide-spread, we propose that ethylene signaling may be part
of a hologenome-level stress response in which genetic
traits carried by both microbiota and the plant are
acti-vated in response to stressors. Ethylene modulation is
under this perspective the result of the co-evolution of
both plant and microbial traits. Bacteria could receive
signals from environment or plants and trigger plant
ethylene response factors (Fig.
3e
) or even express
their own ethylene response factors in response to
plant ethylene or environmental cues (Fig.
3f
).
Such intertwined signaling between plants and
mi-crobes might contribute to complete [
74
] or a partial
[
75
] loss of plant ethylene pathways over the course of
evolution, as observed in the loss of the ACC
biosyn-thetic route in several gymnosperms [
76
] or the
pro-duction of ethylene via an ACC-independent pathway
in several plants [
75
]. From a co-evolutionary
perspec-tive, such plants will become more dependent on
microbiota for ethylene pathway modulation.
Fig. 3aPotential consequences of evolution of an intertwined ethylene signaling involving both plant and microbiota. In ancestral plant phenotype, ACC (1-aminocyclopropane-1-carboxylic acid) is produced by the action of ACC synthase enzymes (ACS). ACC is then converted to ethylene by the enzyme ACC oxidase, triggering different ethylene response factors (ERFs).bBacteria reduce the intensity of stress experienced by the plant. Plant reliance on the microbiome to reduce stressors may lead to a reduced ability of the plant to respond to the acute stressors.c,dBacteria alter ACC and ethylene in plants, leading to over- or under-expression of ethylene pathway genes in plants.
Box 1
What makes co-evolution possible?
a) Plants live in close association with a wide range of
microbes. Roots select and feed a specific
microbiome [77]. A wide range of the
rhizosphere-enriched microorganisms have the ability to
modu-late plant ethylene signaling. For instance, genes
linked to ethylene production or reduction can be
found in a broad range of bacteria and fungi [65,
78,
79]. The constant contact with an ethylene
signaling-altering microbiota may cause the
evolu-tion of a modified pathway optimizing plant
re-sponse in the presence of external perturbations.
b) Positive feedback loops: under stressful conditions,
plants produce more ACC [14]. This confers an
advantage to microorganisms producing ACC
deaminases that are able to use ACC as a nitrogen
and carbon source. This may result in an increased
density of ACC-degrading microbes, whose effect
can be counteracted by the plant by producing more
ACC. The outcome might be beneficial only for
mi-crobes (parasitism of plant nitrogen), or mutually
beneficial (symbiosis via shared ethylene signaling).
c) Plant adaptation to fluctuating environments
requires a rapid rewiring of stress response
pathways such as ethylene signaling. However,
this adaptation may be too slow in plants,
requiring several generations to acquire and
spread the needed mutations. Emergence of
genetic variation in the microbiome is many
orders of magnitude faster than in plants [80].
Modulation of plant hormone levels via the
microbiome may thus provide a new mechanism
to match plant phenotype to environmental
conditions.
d) Modulation of plant hormone levels via the
microbiome may thus provide a new mechanism
to match plant phenotype to environmental
conditions.
e) Ethylene-modulating microorganisms can be
transmitted vertically, from one generation to the
next generation, thus allowing co-evolution of
mi-crobes and the host as a cohesive unit of selection
[80]. Vertical co-evolution may allow more gene
transfer to the next generation, and the
establish-ment of relatively stable associations. Nonetheless,
vertical transmission is probably essential for the
last of our proposed co-evolutionary dynamics
(intertwined signaling; Fig.
3e,
f
). The ethylene
modulation genes could transfer between
ethylene-modulating bacteria by horizontal
transfer [66,
67] and through symbiotic island
exchange [81].
Evolutionary implications of ethylene manipulation by
plant-associated microorganisms
Based on existing scientific evidence, ethylene signaling
most likely evolved within the context of long-term
co-evolution processes between plants and their associated
microbes. We propose that the joint regulation between
microbes and plants can lead to several implications:
a) Alterations of ethylene signaling may offer new
functions and shift the niche of the holobiont
Altering ethylene levels might allow plants to exploit
new niche space, where other trait combinations are
optimal. Co-evolution leading to ethylene
overpro-duction (Fig.
3c) and insensitivity (Fig.
3d) might
also shift plant niches, as well as restrict the chances
for a plant to re-inhabit its ancestral range.
b) Change in plant-encoded ethylene signaling genes
During co-evolution, some microorganisms are
po-tentially part of holobiont-level ethylene-regulated
traits (Fig.
3e,
f
). This association might reduce some
parts of the plant genome working in parallel with
the microbiota, saving the cost of gene expression
and maintenance of redundant genes. In addition to
losing some part of the ethylene signaling pathway,
based on the amount of plant dependency on
associated microbes, dispersal of seeds to new
environments with completely different microbial
communities might cause them to die before they
are able to adapt to the new conditions and pass the
traits down to their offspring.
c) Uncoupling plant phenotype from mutations in the
plant genome
Mutations can alter plant evolution by affecting
different pathways including the ethylene pathway.
Mutations in the plant ethylene biosynthesis and
signaling pathway (for instance, the ability to
overproduce ethylene) could cause new
morphological traits or functions that promote plant
fitness in a new environment, and therefore increase
the chances for natural selection. Associated
microbiota influence this selection by making the
ancestral microbe-associated plants more successful
in competition, thereby decreasing the advantage of
mutations, as microbes might override the
plant-bacteria co-evolution by altering different parts of
the ethylene pathway.
Conclusion
The plant hormone ethylene mediates many aspects
of plant life history. At the holobiont level, ethylene
signaling is a regulatory cascade composed of both
plant- and microbiota-associated traits, which together
environmental conditions and stressors. The holobiont
perspective in plant hormone regulation also has large
evolutionary
implications
in
which
plants
have
become dependent on their microbiome for fully
adaptive ethylene-mediated responses. From an
agri-cultural perspective, the plant holobiont may facilitate
appropriate or maladapted stress responses depending
on the match or mismatch of plant and microbiome
traits. Many aspects of plant health related to the
microbiome and ethylene signaling may represent a
useful model case to further our general
understand-ing of plant holobiont ecology and evolution.
Funding
Not applicable.
Availability of data and materials
Not applicable.
Authors’contributions
MR and AJ wrote the first draft of the paper. All authors provided and critically reviewed the content. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher
’
s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author details
1Ecology and Biodiversity, Institute of Environmental Biology, Utrecht
University, 3584 CH Utrecht, The Netherlands.2Plant Ecophysiology, Institute
of Environmental Biology, Utrecht University, 3584 CH Utrecht, The Netherlands.
Received: 9 November 2017 Accepted: 5 March 2018
References
1. Bohnert H, Nelson D, Jensen R. Adaptations to environmental stresses. Plant Cell. 1995;7:1099–111.
2. Huot B, Yao J, Montgomery BL, He SY. Growth–defense tradeoffs in plants: a balancing act to optimize fitness. Mol Plant. 2014;7:1267–87.
3. Denison RF. Evolutionary tradeoffs as opportunities to improve yield potential. Field Crops Res. 2015;182:3–8.
4. Melotto M, Underwood W, Koczan J, Nomura K, He SY. Plant stomata function in innate immunity against bacterial invasion. Cell. 2006;126:969–80. 5. Araújo WL, Fernie AR, Nunes-Nesi A. Control of stomatal aperture. Plant
Signal Behav. 2011;6:1305–11.
6. Leon-Reyes A, Spoel SH, Lange ESD, Abe H, Kobayashi M, Tsuda S, et al. Ethylene modulates the role of NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 in cross talk between salicylate and Jasmonate signaling. Plant Physiol. 2009;149:1797–809.
7. Vandenkoornhuyse P, Quaiser A, Duhamel M, Le Van A, Dufresne A. The importance of the microbiome of the plant holobiont. New Phytol. 2015; 206:1196–206.
8. Bleecker AB, Kende H. Ethylene: a gaseous signal molecule in plants [Internet].https://doi.org/10.1146/annurev.cellbio.16.1.1. 2003 [cited 2017 Mar 27]. Available from:http://annualreviews.org/doi/abs/10.1146/annurev. cellbio.16.1.1.
9. Sasidharan R, Voesenek LACJ. Ethylene-mediated acclimations to flooding stress. Plant Physiol. 2015;169:3–12.
10. Thao NP, Khan MIR, Thu NBA, Hoang XLT, Asgher M, Khan NA, et al. Role of ethylene and its cross talk with other signaling molecules in plant responses to heavy metal stress. Plant Physiol. 2015;169:73–84.
11. Pieterse CMJ, Leon-Reyes A, Van der Ent S, Van Wees SCM. Networking by small-molecule hormones in plant immunity. Nat Chem Biol. 2009;5:308–16. 12. Scarpeci TE, Frea VS, Zanor MI, Valle EM. Overexpression of AtERF019 delays
plant growth and senescence, and improves drought tolerance in Arabidopsis. J Exp Bot. 2017;68:673–85.
13. Roux F, Touzet P, Cuguen J, Le Corre V. How to be early flowering: an evolutionary perspective. Trends Plant Sci. 2006;11:375–81.
14. Schellingen K, Van Der Straeten D, Vandenbussche F, Prinsen E, Remans T, Vangronsveld J, et al. Cadmium-induced ethylene production and responses in Arabidopsis thaliana rely on ACS2 and ACS6 gene expression. BMC Plant Biol. 2014;14:214.
15. Barry CS, Blume B, Bouzayen M, Cooper W, Hamilton AJ, Grierson D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996;9:525–35.
16. Johnson PR, Ecker JR. The ethylene gas signal transduction pathway: a molecular perspective. Annu Rev Genet. 1998;32:227–54.
17. Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science. 1999;284:2148–52.
18. An F, Zhao Q, Ji Y, Li W, Jiang Z, Yu X, et al. Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis. Plant Cell Online. 2010;22:2384–401.
19. Groen SC, Whiteman NK, Bahrami AK, Wilczek AM, Cui J, Russell JA, et al. Pathogen-triggered ethylene signaling mediates systemic-induced susceptibility to herbivory in Arabidopsis. Plant Cell Online. 2013;25:4755–66. 20. Kacperska A. Ethylene synthesis and a role in plant responses to different
stressors. Biol Biotechnol Plant Horm Ethyl [Internet]. Springer, Dordrecht; 1997 [cited 2017 Oct 12]. p. 207–16. Available from:https://link.springer. com/chapter/10.1007/978-94-011-5546-5_27.
21. Wees SCMV, Pelt JAV, Bakker PAHM, Pieterse CMJ. Bioassays for assessing Jasmonate-dependent defenses triggered by pathogens, herbivorous insects, or beneficial rhizobacteria. Jasmonate Signal [Internet]. Humana Press, Totowa, NJ; 2013 [cited 2017 Oct 3]. p. 35–49. Available from: https://link.springer.com/protocol/10.1007/978-1-62703-414-2_4. 22. van Loon LC, Rep M, Pieterse CMJ. Significance of inducible
defense-related proteins in infected plants. Annu Rev Phytopathol. 2006;44:135–62.
23. Morgan PW, Drew MC. Ethylene and plant responses to stress. Physiol Plant. 1997;100:620–30.
24. Peng J, Li Z, Wen X, Li W, Shi H, Yang L, et al. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. PLoS Genet. 2014;10:e1004664.
25. Keunen E, Schellingen K, Vangronsveld J, Cuypers A. Ethylene and metal stress: small molecule, big impact. Front Plant Sci [Internet]. 2016 [cited 2017 Feb 22];7. Available from:http://www.ncbi.nlm.nih.gov/pmc/articles/ PMC4735362/.
26. Tao J-J, Chen H-W, Ma B, Zhang W-K, Chen S-Y, Zhang J-S. The role of ethylene in plants under salinity stress. Front Plant Sci [Internet]. 2015 [cited 2017 Feb 24];6. Available from:http://journal.frontiersin.org/article/10.3389/ fpls.2015.01059/.
27. Vassilev A, Lidon F, Scotti P, Graca MD, Yordanov I. Cadmium-induced changes in chloroplast lipids and photosystem activities in barley plants. Biol Plant. 2004;48:153–6.
28. Berumen S, Lownds NK. Ethylene production as a drought stress response in Cassia corymbosa. Hortscience. 1996;31:649.
29. Remans T, Thijs S, Truyens S, Weyens N, Schellingen K, Keunen E, et al. Understanding the development of roots exposed to contaminants and the potential of plant-associated bacteria for optimization of growth. Ann Bot. 2012;110:239–52.
30. Wilkinson S, Davies WJ. Drought, ozone, ABA and ethylene: new insights from cell to plant to community. Plant Cell Environ. 2010;33:510–25. 31. Bailey-Serres J, Voesenek LACJ. Flooding stress: acclimations and genetic
diversity. Annu Rev Plant Biol. 2008;59:313–39.
33. Voesenek LACJ, Pierik R, Sasidharan R. Plant life without ethylene. Trends Plant Sci. 2015;20:783–6.
34. Fan Q-J, Liu J-H. Colonization with arbuscular mycorrhizal fungus affects growth, drought tolerance and expression of stress-responsive genes in Poncirus trifoliata. Acta Physiol Plant. 2011;33:1533.
35. Lasserre E, Bouquin T, Hernandez JA, Pech J-C, Balagué C, Bull J. Structure and expression of three genes encoding ACC oxidase homologs from melon (Cucumis melo L.). Mol Gen Genet MGG. 1996;251:81–90. 36. Peng H-P, Lin T-Y, Wang N-N, Shih M-C. Differential expression of genes
encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis during hypoxia. Plant Mol Biol. 2005;58:15–25.
37. Li T, Tan D, Yang X, Wang A. Exploring the apple genome reveals six ACC synthase genes expressed during fruit ripening. Sci Hortic. 2013;157:119–23. 38. Vogel JP, Woeste KE, Theologis A, Kieber JJ. Recessive and dominant
mutations in the ethylene biosynthetic gene ACS5 of Arabidopsis confer cytokinin insensitivity and ethylene overproduction, respectively. Proc Natl Acad Sci. 1998;95:4766–71.
39. Young TE, Meeley RB, Gallie DR. ACC synthase expression regulates leaf performance and drought tolerance in maize. Plant J. 2004;40:813–25. 40. Dal Cin V, Rizzini FM, Botton A, Tonutti P. The ethylene biosynthetic and
signal transduction pathways are differently affected by 1-MCP in apple and peach fruit. Postharvest Biol Technol. 2006;42:125–33.
41. Yang C, Ma B, He S-J, Xiong Q, Duan K-X, Yin C-C, et al. MHZ6/OsEIL1 and OsEIL2 regulate ethylene response of roots and coleoptiles and negatively affect salt tolerance in rice. Plant Physiol. 2015;169:148. pp.00353.2015 42. Pierik R, Sasidharan R, Voesenek LACJ. Growth control by ethylene: adjusting
phenotypes to the environment. J Plant Growth Regul. 2007;26:188–200. 43. Dugardeyn J, Van Der Straeten D. Ethylene: fine-tuning plant growth and
development by stimulation and inhibition of elongation. Plant Sci. 2008; 175:59–70.
44. Zilber-Rosenberg I, Rosenberg E. Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiol Rev. 2008;32:723–35.
45. Berendsen RL, Pieterse CMJ, Bakker PAHM. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012;17:478–86.
46. Ruíz-Sánchez M, Armada E, Muñoz Y, García de Salamone IE, Aroca R, Ruíz-Lozano JM, et al. Azospirillum and arbuscular mycorrhizal colonization enhance rice growth and physiological traits under well-watered and drought conditions. J Plant Physiol. 2011;168:1031–7.
47. Horvath RS. Microbial co-metabolism and the degradation of organic compounds in nature. Bacteriol Rev. 1972;36:146–55.
48. Lloyd JR, Lovley DR. Microbial detoxification of metals and radionuclides. Curr Opin Biotechnol. 2001;12:248–53.
49. Suganuma N, Yamauchi H, Yamamoto K. Enhanced production of ethylene by soybean roots after inoculation with Bradyrhizobium japonicum. Plant Sci. 1995;111:163–8.
50. Weingart H, Volksch B. Ethylene production by Pseudomonas syringae pathovars in vitro and in planta. Appl Environ Microbiol. 1997;63:156–61. 51. Fukuda H, Ogawa T, Tanase S. Ethylene production by micro-organisms. In:
Rose AH, editor. Adv Microb Physiol [Internet]. Academic Press; 1993 [cited 2017 Sep 28]. p. 275–306. Available from:http://www.sciencedirect.com/ science/article/pii/S0065291108601010
52. Freebairn HT, Buddenhagen IW. Ethylene production by Pseudomonas solanacearum. Nature. 1964;202:313–4.
53. Nagahama K, Ogawa T, Fujii T, Fukuda H. Classification of ethylene-producing bacteria in terms of biosynthetic pathways to ethylene. J Ferment Bioeng. 1992;73:1–5.
54. Eckert C, Xu W, Xiong W, Lynch S, Ungerer J, Tao L, et al. Ethylene-forming enzyme and bioethylene production. Biotechnol Biofuels. 2014;7:33. 55. Sato M, Watanabe K, Yazawa M, Takikawa Y, Nishiyama K. Detection of new
ethylene-producing bacteria, Pseudomonas syringae pvs. cannabina and sesami, by PCR amplification of genes for the ethylene-forming enzyme. Phytopathology. 1997;87:1192–6.
56. Jäckel U, Schnell S, Conrad R. Microbial ethylene production and inhibition of methanotrophic activity in a deciduous forest soil. Soil Biol Biochem. 2004;36:835–40.
57. Digiacomo F, Girelli G, Aor B, Marchioretti C, Pedrotti M, Perli T, et al. Ethylene-producing bacteria that ripen fruit. ACS Synth Biol. 2014;3:935–8.
58. Veselov D, Langhans M, Hartung W, Aloni R, Feussner I, Götz C, et al. Development of Agrobacterium tumefaciens C58-induced plant tumors and
impact on host shoots are controlled by a cascade of jasmonic acid, auxin, cytokinin, ethylene and abscisic acid. Planta. 2003;216:512–22.
59. Patten CL, Glick BR. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol. 2002;68:3795–801. 60. Vogel JP, Schuerman P, Woeste K, Brandstatter I, Kieber JJ. Isolation and
characterization of Arabidopsis mutants defective in the induction of ethylene biosynthesis by cytokinin. Genetics. 1998;149:417–27.
61. Yeong-Biau Y, Shang Fa Y. Auxin-induced ethylene production and its inhibition by aminoethyoxyvinylglycine and cobalt ion|Plant Physiology [Internet]. [cited 2017 Oct 12]. Available from:http://www.plantphysiol.org/content/64/6/1074. 62. Lorteau M-A, Ferguson BJ, Guinel FC. Effects of cytokinin on ethylene
production and nodulation in pea (Pisumsativum) cv. Sparkle Physiol Plant. 2001;112:421–8.
63. Glick BR. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res. 2014;169:30–9.
64. Singh RP, Shelke GM, Kumar A, Jha PN. Biochemistry and genetics of ACC deaminase: a weapon to“stress ethylene”produced in plants. Front Microbiol [Internet]. 2015 [cited 2017 Mar 27];6. Available from:http:// journal.frontiersin.org/article/10.3389/fmicb.2015.00937/abstract.
65. Nascimento FX, Rossi MJ, Soares CRFS, McConkey BJ, Glick BR. New insights into 1-aminocyclopropane-1-carboxylate (ACC) deaminase phylogeny. Evolution and Ecological Significance PLOS ONE. 2014;9:e99168.
66. Blaha D, Prigent-Combaret C, Mirza MS, Moënne-Loccoz Y. Phylogeny of the 1-aminocyclopropane-1-carboxylic acid deaminase-encoding gene acdS in phytobeneficial and pathogenic Proteobacteria and relation with strain biogeography. FEMS Microbiol Ecol. 2006;56:455–70.
67. Hontzeas N, Richardson AO, Belimov A, Safronova V, Abu-Omar MM, Glick BR. Evidence for horizontal transfer of 1-aminocyclopropane-1-carboxylate deaminase genes. Appl Environ Microbiol. 2005;71:7556–8.
68. Ravanbakhsh M, Sasidharan R, Voesenek LACJ, Kowalchuk GA, Jousset A. ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J Ecol. 2017;105:979.
69. Hall JA, Peirson D, Ghosh S, Glick BR. Root elongation in various agronomic crops by the plant growth promoting rhizobacterium Pseudomonas putida Gr12–2. Isr J Plant Sci. 1996;44:37–42.
70. Vacheron J, Combes-Meynet E, Walker V, Gouesnard B, Muller D, Moënne-Loccoz Y, et al. Expression on roots and contribution to maize
phytostimulation of 1-aminocyclopropane-1-decarboxylate deaminase gene acdS in Pseudomonas fluorescens F113. Plant Soil. 2016;407:187–202. 71. Desbrosses G, Contesto C, Varoquaux F, Galland M, Touraine B.
PGPR-Arabidopsis interactions is a useful system to study signaling pathways involved in plant developmental control. Plant Signal Behav. 2009;4:319–21. 72. Koevoets IT, Venema JH, Elzenga JTM, Testerink C. Roots withstanding their environment: exploiting root system architecture responses to abiotic stress to improve crop tolerance. Front Plant Sci [Internet]. 2016 [cited 2018 Jan 26];7. Available from:https://www.ncbi.nlm.nih.gov/pmc/articles/ PMC5005332/.
73. Carlos M-HJ, Stefani P-VY, Janette A-M, Melani M-SS, Gabriela P-O. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol Res. 2016;188:53–61.
74. Golicz AA, Schliep M, Lee HT, Larkum AWD, Dolferus R, Batley J, et al. Genome-wide survey of the seagrass Zostera muelleri suggests modification of the ethylene signalling network. J Exp Bot. 2015;66:1489–98.
75. Osborne DJ, Walters J, Milborrow BV, Norville A, Stange LMC. Special publication evidence for a non-ACC ethylene biosynthesis pathway in lower plants. Phytochemistry. 1996;42:51–60.
76. Groen SC, Whiteman NK. The evolution of ethylene signaling in plant chemical ecology. J Chem Ecol. 2014;40:700–16.
77. Campbell R, Greaves MP. Anatomy and community structure of the rhizosphere. In: Lynch JM, editor. The rhizosphere. Chichester, England: Wiley 1990:11–34.
78. Duan J, Müller KM, Charles TC, Vesely S, Glick BR. 1-Aminocyclopropane-1-carboxylate (ACC) deaminase genes in rhizobia from Southern Saskatchewan. Microb Ecol. 2009;57:423–36.
79. Smith AM, Cook RJ. Implications of ethylene production by bacteria for biological balance of soil. Nature. 1974;252:703–5.
80. Rosenberg E, Zilber-Rosenberg I. Microbes drive evolution of animals and plants: the Hologenome concept. MBio. 2016;7:e01395–15.
82. van Veen H, Mustroph A, Barding GA, Eijk MV, Welschen-Evertman RAM, Pedersen O, et al. Two Rumex species from contrasting hydrological niches regulate flooding tolerance through distinct mechanisms. Plant Cell Online. 2013;25:4691–707.
83. Guan R, Su J, Meng X, Li S, Li Y, Xu J, et al. Multi-layered regulation of ethylene induction plays a positive role in Arabidopsis resistance against Pseudomonas syringae. Plant Physiol. 2015;169:299–312.
84. Arteca RN, Arteca JM. Effects of brassinosteroid, auxin, and cytokinin on ethylene production in Arabidopsis thaliana plants. J Exp Bot. 2008;59:3019–26.
• We accept pre-submission inquiries
• Our selector tool helps you to find the most relevant journal
• We provide round the clock customer support
• Convenient online submission
• Thorough peer review
• Inclusion in PubMed and all major indexing services
• Maximum visibility for your research
Submit your manuscript at www.biomedcentral.com/submit