lis'; ill! ! ' ! 1 )
H
l'\ 1? ii * ! 1 'ii'';'.;!-I 1 Ju
) 1 [! ) i Ih'i'M
. *!hi
!* •lliVV^il,ViM
I( tipiiiio'V':.":
1 'I!}bH
'! 1 i t I )StP'i'ir"
' I »• ill. ) !i. 1 1 * hliw.^:!', J iaii! film illliilP:,,:.:,'!!;:'.!ili!fei!!|!:!i;-^Vv:n::ii;:i;iM,
P
ii|ii;iii ill.li''!'.'!::- ','•! -'..I'The
person chargingthismaterial isresponsiblefor
Itsreturntothe
libraryfrom whichitwaswithdrawn onor before the Latest
Date stamped below I^r;T*"'*"L"'""**""«'«^i"*"fl 01booksare reasonsfor
discipli-naryactionandmayresultindismissal from the University
TorenewcallTelephoneCenter,333-8400
UNIVERSITY OF ILLINOIS LIBRARY AT URBANA-CHAMPAIGN
NOVOy
/RfiiST.JRBAMa
C
PAIGN
FIELDIANA
Geology
NEW
SERIES,
NO.
32
Pachypleurosaurs
(Reptilia:
Sauropterygia)
from
the
Lower
Muschelkalk, and
a
Review
of the
Pachypleurosauroidea
Olivier
Rieppel
Lin
Kebang
December
29,1995
Publication1473
Fll
1
1
nil
Information
forContributors
toFieldiana
Generak Fieldiana is primarily a journal for Field Museum staff members and research associates, although manuscriptsfromnonaffiliatedauthorsmaybeconsidered as space permits.
TheJournalcarriesapage chargeof $65.00perprintedpage orfraction thereof.Paymentofat least
50%
ofpage chargesqualifiesapaperfor expedited processing,whichreduces the publication time. Contributionsfromstaff,researchassociates,andinvitedauthorswillbeconsidered for publication regardless ofabilitytopay pagecharges,however,thefull
chargeismandatoryfor nonaffiliatedauthorsof unsolicitedmanuscripts.Threecompletecopies of thetext(includingtitle
pageandabstract)andof theillustrationsshouldbe submitted (oneoriginalcopyplustworeview copieswhichmaybe machine-copies).
No
manuscriptswill beconsidered for publicationor submitted to reviewers beforeallmaterials are complete andinthehands oftheScientificEditor.Manuscripts should be submittedtoScientificEditor, Fieldiana, Field
Museum
ofNaturalHistory,Chicago,Illinois60605-24%,USA.
Text: Manuscriptsmustbetypewrittendouble-spaced onstandard-weight,8Vi-by11-inch paperwithwide margins on all four sides. If typed on an IBM-compatible computer using MS-DOS, also submit text on 5y4-inch diskette (WordPerfect4.1, 4.2, or5.0, MultiMate, Displaywrite 2, 3
&
4,Wang
PC, Samna, Microsoft Word, Volkswriter, or WordStarprograms or ASCII).For papers over100manuscriptpages, authors are requested tosubmitaTableof Contents," a"ListofIllustrations,"
anda"Listof Tables"immediatelyfollowingtitlepage.Inmostcases,thetextshouldbe precededbyan"Abstract"and should concludewith"Acknowledgments"(ifany)and"LiteratureCited."
Allmeasurementsshouldbeinthe metricsystem(periods are notusedafterabbreviatedmeasurements).Theformat andstyleofheadings shouldfollowthatof recentissuesof Fieldiana.
Formoredetailedstyle information, seeThe ChicagoManualofStyle(13thed.),publishedbyThe University of ChicagoPress,andalso recent issues of Fieldiana.
References: In "Literature Cited,"book andjournaltitlesshouldbegiveninfull.Whereabbreviationsare desirable
(e.g.,in citationof synonymies), authorsconsistentlyshouldfoWowBotanico-Periodicum-Huntianum and TL-2 Taxonomic
LiteraturebyF.A.Stafleu«&
R
S.Cowan(1976etseq.)(botanical papers)orSerialSources fortheBiosisDataBase(1983) publishedbytheBioSciences InformationService.Namesof botanicalauthorsshouldfollow the "DraftIndexofAuthor Abbreviations,Royal Botanic Gardens, Kew," 1984edition,or TL-2.References shouldbetypedinthe following form:
Croat, T. B. 1978. Flora ofBarro ColoradoIsland.Stanford University Press, Stanford,Calif.,943pp.
Grubb,P.J.,J.R. Lloyd,andT.D. Pennington.1963.
A
comparisonofmontane andlowlandrainforest inEcuador.I.Theforest structure,physiognomy, andfloristics.Journal of Ecology,51:567-601. Langdon,E.J.M.1979.Yageamongthe Siona: Cultural patterns
in visions,pp.63-80.InBrowman,D.L.,and R.A. Schwarz,eds., Spirits,Shamans, andStars.MoutonPublishers,TheHague,Netherlands.
Murra, J. 1946.The historic tribesof Ecuador,pp. 785-821. In Steward,J. H.,ed.. HandbookofSouthAmerican Indians. Vol. 2,The Andean Civilizations. Bulletin 143, Bureau ofAmerican Ethnology, Smithsonian
Institution,Washington, D.C.
Stolze,
R
G.1981.FernsandfernalliesofGuatemala.PartII.Polypodiaceae. Fieldiana:Botany,n.s., 6:1-522. Illustrations: Illustrationsare referred to as"figures" inthetext(not as"plates").Figuresmustbe accompaniedby someindicationofscale, normallya referencebar.Statementsinfigurecaptions alone,suchas "x0.8,"arenot acceptable. Captions should betypeddouble-spaced andconsecutively.Seerecentissuesof Fieldiana fordetailsofstyle.
Allillustrationsshouldbemarked onthe reverse with author'sname, figurenumber(s), and"top."
Figures assubmittedshould,wheneverpracticable,be8V4x11inches (22x28 cm),andmaynotexceed W/tx16V4 inches (30x 42cm).Illustrationsshouldbemounted onboardsinthearrangementtobe obtainedinthe printedwork.This
original setshouldbesuitable for transmission to theprinter asfollows:Pen andinkdrawingsmaybeoriginals(preferred) orphotostats;shaded drawings must beoriginals,but within thesize limitation;andphotostatsmustbehigh-quality, glossy,
black-and-whiteprints.Originalillustrations willbereturnedtothecorresponding authoruponpublication unless otherwise
specified.
Authors whowish to publish figures that require costlyspecial
paper or color reproduction must make prior arrangementswith theScientificEditor.
PageProofs: Fieldianaemploysa two-step correction system.The
corresponding authorwillnormallyreceive acopy of the editedmanuscriptonwhichdeletions,additions,and changes can bemadeandqueriesanswered.Only onesetof pageproofswillbesent.All desired corrections oftypemust bemadeonthesingle setofpageproofs.Changesinpage
proofs(asopposedto corrections) areveryexpensive.Author-generated changesinpageproofscanonlybe madeifthe authoragreesinadvancetopayforthem.
aeOlOflY
UBRAI^fY
FIELDIANA
Geology
NEW
SERIES,
NO.
32
Pachypleurosaurs
(Reptilia:
Sauropterygia)
from
the
Lower
Muschelkalk, and
a
Review
of the
Pachypleurosauroidea
Olivier
Rieppel
Department of Geology
Field
Museum
ofNatural History RooseveltRoad
atLakeShoreDrive Chicago,Illinois60605-2496 U.S.A.Lin
Kebang
Redpath
Museum
McGillUniversityMontreal, Quebec
H3A
2K6 CanadaAccepted
June
1,1995
Published
December
29,1995
Publication
1473
©
1995FieldMuseum
ofNatural History LibraryofCongressCatalogCard Number:
95-70458ISSN
0096-2651Table
of
Contents
Abstract
1zusammenfassung
1Introduction
1 SystematicPaleontology
2 Phylogenetic Analysis 21 DefinitionofCharacters 24Genera
and
Speciesof
Pachypleuro-SAUROIDEA inTHE
MIDDLE
AND
UPPER
Triassic 34
Historical
Biogeography
of
the
Pachypleurosauroidea
36The
Paleoeoology of Pachypleurosaurs .. 40Acknowledgments
40Literature Cited 41
Appendix:
Mateiual Included
and
Institutional Abbrevl\tions 43
List
of
Illustrations1. Castsofholotypeoi
Anarosaums
pumilio
Dames
32. Charitosaurustschudii
Meyer
43. Skullof Anarosaurus pumilio
Dames
(castofholotype) 5 4. Skullof Anarosaurus pumilio
Dames
... 55. GastralribsofAnarosauruspumilio
Dames
76. Pubisof Anarosaurus pumilio
Dames
.. 77.
Humerus
and femurofAnarosauruspumilio
Dames
8. Holotype of Anarosaurusheterodontus
n. sp 9
9. Leftpremaxillaof Anarosaurus hetero-dontusn. sp 9 10. SkullofAnarosaurusheterodontus
n. sp 10
1 1. Holotype ofDactylosaurusgracilis
Giirich 12
12. Holotype ofDactylosaurusschroederi
Nopcsa
1313. DactylosaurusgracilisGiirichfrom the lower Muschelkalk ofGogolin,
Upper
Silesia
(Gomy
Slask, Poland) 14 14. Dactylosaurusgracilis Giirich,humerus,fromthelower MuschelkalkofBobrek
near Beuthen 14
15. Right forelimbofDactylosaurusgracilis
Giirich 15
16. SVxAl ofDactylosaurusgracilisGurich
..16
17. Pectoral girdleofDactylosaurusgracilisGiirich 17
18. PubisofDactylosaurusgracilisGurich
..18
19.
Humerus
ofDactylosaurusgracilisGiirich 18
20.
Manus
andpesofDactylosaurusgracilisGiirich 19
21. Sauropterygiaindet. fromthelower
Muschelkalk ofGogolin,
Upper
Silesia(Gomy
Slask, Poland) 2122. Strictconsensustreeofsauropterygian interrelationships 23
23. Unrooted networkforPlacodus,
Pachy-pleiu-osauroidea,
and
selected represen-tativesofEusauropterygia 31 24.The
singlemost
parsimonioustreeforpachypleurosaurinterrelationships rooted
on
themonophyleticEusau-ropterygia 32
25. Strictconsensustreefor pachypleuro-saursandselectedeusauropterygians, rooted
on
anall-0 ancestor 3326. Strictconsensustreefor Placodus, pachy-pleurosaurs,andselected eusauroptery-gians,rooted
on
anall-0ancestor 3327. Strictconsensustreefor pachypleuro-saurinterrelationships,rootingthe pachypleurosaursplus selected eusau-ropterygian taxaon Placodus 34
28. Reconstructionsof Anarosauruspumilio
and
Dactylosaurusgracilis 378 List
of
Tables
1. Measurements ofthe
humerus
ofAnaro-saurus pumilio
Dames
8 2. Measurements ofthefemur ofAnarosau-ruspumilio
Dames
83.
Body
proportionsinPachypleurosauroi-dea 11
4. Measurements ofthe
humerus
ofDactylosaurusgracilisGiirich 18
5. Measurements ofthelimb bonesof
Dac-tylosaurusgracilis 19 6. Datamatrixfor 14 taxaand 50
Pachypleurosaurs
(Reptilia:
Sauropterygia)
from
the
Lower
Muschelkalk, and
a
Review
of the
Pachypleurosauroidea
Olivier
Rieppel
Lin
Kebang
Abstract
The
type material ofall pachypleurosaurs from thelower Muschelkalkis redescribedandpachypleurosaur systematicsare reviewed. Three speciesoflower Muschelkalk pachypleuro-saursarerecognized,Anarosauruspumilio
Dames,
Anarosaurusheterodontusn. sp.,andDac-tylosaurnsgracilisGiirich.
A
cladisticanalysisbasedon
50charactersshowsKeichousaurustobe the sister-taxon toall other pachypleurosaurs; Dactylosaurusis thesister-taxonto
Anaro-saurus plus the Serpianosaurus-Neusticosaurusclade; Anarosaurus is the sister-taxon to the Serpianosaurus-Neusticosaurusclade;Serpianosaurusmirigiolensisisthesister-taxon to
Neus-ticosaurus.
The
genusNeusticosaurus includes fourspecies,A^. edwardsii,N.peyeri,N.pusillus,and
N. toeplitschi.The
stratigraphicand
biogeographicrelations ofpachypleurosaurs indicate thatpachypleurosaurs reachedtheEuropeanepicontinentalsea(MuschelkalkBasin)byinvasionfromtheeast inAnisiantimes,andthata faunal interchangewaspossiblebetweenthe
Muschel-kalkBasin
and
thesouthern Alpine intraplatform basinfacies at leastduringlateAnisianand Ladiniantimes.Zusammenfassung
Das
Typusmaterial aller Pachypleurosaurier ausdem
unteren Muschelkalk wird neu be-schrieben.DreiArtenvon
Pachypleurosauriemlassen sichim
unteren Muschelkalkfeststellen:Anarosauruspumilio
Dames,
Anarosaurusheterodontusn.sp.,und
Dactylosaurusgracilis Gur-ich. EinekladistischeAnalyseweist Keichousaurusals SchwestergruppealleranderenPachy-pleurosaurieraus;DactylosaurusistdieSchwestergruppe
von
Anarosaurusund
der Serpiano-saurus-Neusticosaurus-Linie;Anarosaurus istdieSchwestergruppe der Serpianosaurus-Neus-ticosaurus-lAme;SerpianosaurusmirigiolensisistdieSchwestergruppevon
Neusticosaurus.Die Gattung Neusticosaurus umfasst vierArten: A^. edwardsii, N. peyeri, N. pusillus,und
A^. toe-plitschi.Stratigraphischeund
biogeographischeBeziehungenzeigen,dassdiePachypleurosaurier das Germanische Beckenvon
Ostenherkommend
im
Unteren Aniserreichten.ZwischenderGermanischen
und
derAlpinenTriasfandzumindestwarenddem
spatenAnisund
dem
Ladin einFaunenaustauschstatt.Introduction
lesia(Dactylosaurusgracilis,Giirich, 1884)andin thelowerMiddleTriassicofChina (Young, 1958,The
Pachypleurosauroidea (Pachypleurosauri- 1965).The
clade has not been recorded beyonddae) constitutea monophyletic clade ofthe sau- the upperLadinian (upperMiddleTriassic), per-ropterygian radiation (Rieppel, 1987, 1989;Storrs, haps lower
Camian
(lowerUpper
Triassic), de-1991; Sues, 1987)thatfirstappears inthelower- posits in the southernAlps (Ca' del Frate, Italy:iswell
known
fortheMiddleTriassictaxaabun-dantly represented in uppermost Anisian
and
lower Ladinian deposits at
Monte
San Giorgio, Switzerland (genera Neusticosaurusand
Serpiano-saurus: Carroll&
Gaskill, 1985; Rieppel, 1989; Sander, 1989), but the interrelationships of pa-chypleurosaursin general stillremainpoorly un-derstood. This results from the relatively poor knowledge ofearlierrepresentativesoftheclade.Of
these,only aguttaperchacast oftheholotype of DactylosaurusschroederiNopcsa
(1928)from theMuschelkalk ofGross-Stein,Upper
Silesia,has recently been restudied (Sues&
Carroll, 1985; DactylosaurusschroederiNopcsa
is considered ajunior
synonym
oiDactylosaurusgracilisGiirich: see below).A
second pachypleurosaur from the lowermiddle Muschelkalk, Anarosaurus pumilioDames,
1890,from RemkerslebeninSaxony, has not been restudied since Arthaber's (1924) and Nopcsa's (1928) contradictorycomments on
itsmorphology(areconstructionoftheskull
was
pub-lishedbyCarroll, 1981,Fig. 32a).Since the early original description of pachy-pleurosaurid taxa from the lower Muschelkalk
(lowerto middle Anisian),
new
material hasac-cumulated in various
museum
collections, the identificationofwhich remainsproblematicuntil the diagnosesoftheoriginally described taxaare refined. Itisthe purpose ofthispaperto pull to-gether all available information on the original type materialofpachypleurosaursfromthelower Muschelkalk.A
redescriptionoftheoriginaltype material will alsoprovide the basis fora reviewofpachypleurosaurinterrelationshipsandthe re-constructionoftheir evolutionary
and
paleobio-geographichistory.Systematic Paleontology
Sauropterygia
Owen,
1860PachypleurosauroideaHuene, 1956
PachypleurosauridaeNopcsa, 1928
Definition—
A
monophyletic group includingthe
common
ancestor ofAnarosaurus, Dactylo-saurus, Keichousaurus,and
allofitsdescendants.Diagnosis—
Smalltomedium-sizedSauropter-ygiawith thepreorbitalregion oftheskulllonger than the postorbital region; frontalwithconcave
lateraledge, enteringthedorsalmargin ofthe
or-bit; broad parietal skulltable; mandibular
artic-ulation at level with occipital condyle; quadrate withconcaveposteriormargin; ectopterygoid ab-sent;trough
on
dorsal surfaceofretroarticular pro-cess present; anterior teethnot strongly procum-bent; posteriorprocesson
interclaviclerudimen-tary or absent; radius slightly longer than ulna; iliacblade absent (i.e., reducedto narrowdorsal
process).
Distribution—
Middle Triassic ofChinaand
Europe (TethyanProvince).Anarosaurus
Dames,
1890Type
SPECIES—Aruirosaurus pumilioDames,
1890.
Generic Diagnosis—
A
small pachypleurosaur with aspatulateexpansion ofthetooth crown; a high dorsal vertebral count (25 or 26); an open obturatorforameninthepubis; pubis with a dis-tinct anteroventral spine;and
a distinctly elon-gatedfemur(femur length/standardlength=
1.55).Distribution—
Lower and
middleMuschelk-alk,centralEurope.
Anarosaurus pumilio
Dames,
18901890 Anarosauruspumilio.
Dames,
p. 74,PI. 1,textfigs. 1,2.
1899 Anarosauruspumilio,
Schrammen,
p.408, PI. 25,Fig. 8.1910 Anarosauruspumilio,Jaekel,pp.325, 335, textfig. 1.
1924 Anarosauruspumilio, Arthaber, pp.
480-483,textfigs. 12a,b, 13.1928 Anarosauruspumilio, Nopcsa, pp. 30-31, 43,PI. 4, Figs. 1, 2.
1935 Anarosauruspumilio, Zangerl, pp. 64-65, 68.
1959 Anarosauruspumilio, Kuhn-Schnyder, p. 652.
1981 Anarosauruspumilio,Carroll, textfig.32a.
1985 Anarosauruspumilio. Sues
and
Carroll, p. 1349.1987 Anarosauruspumilio, Rieppel, pp. 1110,
1115-1116.
1993 Anarosauruspumilio, Rieppel,p. 10.
Holotype—
The
holotypewasoriginallyhousedattheInstitut
und
Museum
fiirGeologicund
Pa-laontologie, Georg-August-Universitat, Gottin-gen, butcanno
longerbe located today(lost orFig.1. Castsoftheholotype o{AnarosauruspumilioDames,depositedbyNopcsainthe British
Museum
(NaturalHistory).A, Dorsalview(bmnhR-5691);B, ventralview(bmnhR-5866).Scalebar
=
20mm.
destroyed during
World
War
II;H.Jahnke,inlitt.1991).
Two
castsare available at this institution (Orig. Nr. 409-1).The
specimencame
from the orbicularis beds near Remkersleben,approxi-mately 15
km
westof Magdeburg.The
orbicularis beds were assigned to the top ofthe lowerMu-schelkalkby
Dames
(1890), but arenow
consid-ered the base ofthe middle Muschelkalk(Hag-dom,
1991).Arthaber(1924)describesa guttaperchacastof
the holotype in the Natural History
Museum,
Humboldt
University, Berlin(mbR. 57.1-3, dor-sal view;mb
R. 58, ventral view), deposited byJaekel (1910)afterfurtherpreparationofthe ho-lotype.
Additional casts (Fig. 1) were deposited by
Nopcsa
inthe BritishMuseum
(Natural History)(BMNH
R-5691, dorsal view;bmnh
R-5866, ven-tralview).Othercastsare keptattheInstitutfur Geowissenschaften, Martin-Luther-Universitat,Fig. 2. CharitosaurustschudiiMeyer. A, Cast oftheEsperstadtspecimen(original of Meyer, 1851,PI. 31,Fig.
22;
MB
R. 62);B, castoftheQuerfurtspecimen(originalof Meyer, 1851,PI. 31,Fig.23;mb
R.61).Scalebar=
20Halle (M4/12, skull only),
and
at the StaatlichesMuseum
furNaturkunde,Stuttgart(smns59073).Diagnosis—
Same
as forgenus, butabout half the sizeofAnarosaurusheterodontus,homodont
dentitionwitheight teethinpremaxilla,andaweak
sculpturingofdermalskullbones.
Locus
Typicvs—
Orbicularis beds (middleMuschelkalk) of Remkersleben, 15
km
west of Magdeburg,Germany.
Distribution—
Lower
middle Muschelkalk,Anisian, Europe.
Comments—
Anarosaurus pumilio was origi-nally described byDames
(1890).Schrammen
(1899) found Anarosaurus to be closelycompa-rable to Cymatosaurus, perhaps because of the spatulate expansion ofthe tooth crown. Jaekel (1910) reconstructed theskullof^4narosaurusafter furtherpreparationofthe holotype,
and
usedAn-arosaurus (as well as Simosaurus) in support of
hisargumentthatsauropterygiansareasubgroup
ofthe Diapsida. Arthaber (1924) criticized Jae-kel's (1910) reconstructionofthe skull, and pre-sented
new
vertebral countsin his redescriptionofAnarosaurus.
He
alsonotedthesimilarityofthe pubisofAnarosaurusto other, isolated elementsfrom thelower Muschelkalkreferred to
'''Macro-tracheW
by H.v.Meyer
(1847-1855).Nopcsa
(1928)rejectedArthaber's(1924)vertebralcounts
inAnarosaurus,
and compared
thetaxontoDac-tylosaurus.
He
considered theskulltobevery sim-ilarin both genera, but found differences in the postcranial skeleton to support the generic dis-tinctionofthetwotaxa.In1958,
Huene
described asecondspecies,"^«-arosaurus"multidentatus,fromthelowerAnisian
ofthe Lechtaler Alps,Austria.
The
specimen con-sistsofalowerjaw
characterizedby an elongatesymphysis
and
aheterodontdentition.Inarecent redescriptionofthe holotype, thespecieshasbeen referred to the genus Cymatosaurus (Rieppel,1995).
In 1838,
Meyer
reported the collection oftwolower
jaw
fragments(leftdentaries), onefromthe Saurierkalk (lowermiddle Muschelkalk) of Esper-stadt, the other from the orbicularis beds(lowermiddle Muschelkalk) ofQuerfurt.
The
originalof the Esperstadtspecimenwas
keptinDresden,thatfromQuerfurt
was
kept in Jena. Both specimensare
now
lost,butcastsof both specimensare pre-servedintheStaatlichesMuseum
fiirNaturkundein Stuttgart (smns 80082-83), and in the
Hum-boldtMuseum
in Berlin (mb R. 61-62) (Fig. 2).The
specimensfirst reportedin 1838 were rede-scribed as lowerjaws offishes byMeyer
(1851),who
renamed
them
Charitodon tschudii. Chari-todon {Charitosaurus) tschudii resemblesAnaro-saurusintoothstructure, with aconstrictedbase
and an expandedcrown, but thetooth crown of
Anarosaurusis
more
distinctly lanceolatethanthe ratherbulboustooth crown ofCharitosaurus.At
the presenttime, Charitosaurustschudii
must
re-main
anomen
dubium.Morphological Description— The
holotypeofAnarosaurus pumiliois
known
fromcasts only.The
original specimen consisted ofpart (dorsal view)and
counterpart(ventralview).The
Instituteand
Museum
of Geology andPaleontologyinGot-tingen holdstwocastsofthedorsalviewthatdiffer in size; measurements of limb bones takenfrom
one cast
(Go
409-lb) beingconsistently smaller than those takenfromtheother(Go
409-la).Oth-er casts kept at the natural history
museums
in Berlin, London, and Stuttgart (see above) yield different values again, indicatingthe limitedFig. 3. Cast ofthe skull ofAnarosauruspumilio
Dames
(holotype)(InstitutfiirGeowissenschaften, Mar-tin-Luther-Universitat, Halle, M4/12). Scale bar=
20mm.
curacyofthosecasts.Unlessnotedotherwise, the
measurementsgivenbelowarefromtheGottingen cast.
Go
409-la.The
skull(Figs. 3,4)isincompletely preserved,partially crushed,and (or)destroyed by prepara-tion. In dorsal view, both premaxillae, the right maxilla, right prefrontal,
and most
ofthe skull tableare wellexposed.The
distancefromthetipofthe snout to the posterior margin ofthe skull table(aspreserved)is34.5
mm,
thedistancefromthetipofthesnouttotheposteriormargin ofthe supraoccipital(as preserved) is38
mm.
The
lon-gitudinaldiameterofthe (right)orbitis 13mm,
the longitudinal diameterofthe upper temporalfossa is 5
mm
(right)and
4.5mm
(left), respec-tively.The
premaxilla formstheanteriorandantero-FiG.4. SkullofAnarosauruspumilio
Dames
(holotype).A,Drawingofthe castkeptattheInstitutundMuseum
furGeologicundPalaontologie,Georg-August-Universitat, Gottingen,Orig.Nr.409-1); B, reconstructionoftheskull
indorsal view. Scale bar
=
10mm.
Abbreviations:ar,articular;f, frontal;m,maxilla;p, parietal;pm,premaxilla; po,i>ostorbital;pof,postfrontal;prf,prefrontal;so,supraoccipital;sq,squamosal.dorsalmargin oftheexternalnaris.
A
slender pos-teriorprocess enteredbetweentheexternal nares, but its relation to the nasaland
frontal remainsunknown. The
posteriorprocessofthepremaxilladoesnotextend
beyond
theleveloftheposteriormargin oftheexternalnaris.Jaekel (1910)counted
four, Arthaber (1924) five premaxillary teeth,
numbers
thatseem
to refer to the incompletely preserved left premaxilla.The
exact tooth countisdifficult toestablish
on
theavailablecasts, but the completely preserved right premaxilla shows a possiblemaximum
ofeight teeth ortooth po-sitions.Dames
(1890)and
Jaekel (1910)showed
acharacteristicleaflikeexpansion ofthe
crown
ofthe premaxillaryteeth,which cannot be confirmed
on the cast kept atthe Institute
and
Museum
of Geology and Paleontologyin Gottingen; a spatu-lateexpansion ofthetoothcrown
isdistinct inoneanteriorpremaxillary toothinthecastskeptatthe Staatliches
Museum
fiirNaturkunde in Stuttgartas well as in the castskeptattheNatural History
Museum
in Berlin.The
remaining premaxillary teeth areabout3mm
long,slightlyrecurved,and
pointed.The
exact structure of the maxillare-mainsobscure, asdoesthe
number
ofmaxillary teeth.Posteriorly,themaxillaformsashort slen-derprocess, underlying the prefrontal in the an-teroventralmargin oftheorbit.The
prefrontalisa broad,domed
element,asistypical forpachypleurosaurs.Itremainedbroadly separated from the postfrontal along the dorsal
margin oftheorbit,whichisformed bytheslightly
concavelateralmargin ofthefrontal.
The
partially fused frontalsare longandslender elements, but theiranteriorportion ismissing. Posteriorly, the frontals form an interdigitating suture with the parietalsinaratherforwardposition.Slender pos-terolateral lappetsofthefrontalsextendbackwardalongthemedialsideofthepostfrontals.
The
pa-rietals enclosea small parietal foramen that liesmidway
betweenthefrontoparietalsutureand
the posterior margin ofthe parietals. Laterally, the parietalformsthemedial margin oftheuppertem-poralfossa. Posteriorly, tapering processesofthe parietalextendlaterally
on
totheocciput.The
upper temporal fossa inAnarosaurus hasrounded contours, and is smaller than the orbit but distinctly larger than the temporal fossa in
Serpianosaums andNeusticosaurus.
The
postfron-tal forms a considerable portion ofthe anterior
andanteromedial margin ofthe upper temporal fossa, the triradiate postorbital forms the lateral margin, andthesquamosalborders the temporal fossa posteriorly.
The
upper temporalarchisnar-row, leaving the cheekdeeply "excavated."
The
ventralprocessoftheleftpostorbital carriesafaceton
itsanteriormarginthatreceivedthedorsalend ofthejugal. Details ofthe suspensoriumand
ofthe occiputaredifficulttoascertainexceptforthe presenceofalarge supraoccipitalwith alow
sag-ittalcrest.
j
The
palate isdifficultto analyze. Littlecan be'
seen
beyond
the generaloutlinesofthemajorel-ements
and
thelocationofthe internalnares(Car-roll, 1981). Inparticular, itisimpossibleto con-firm the presence or absenceofthe ectopterygoid.
A
distinct ventral ridgemarks
thelateral edgeofthe pterygoid along the anteromedialand medial margin ofthesubtemporalfossa.
Lateral to the left suspensorium, the articular facetofthelower
jaw
ispreservedinsitu,withthe retroarticularprocessextendingbackward behindthemandibulararticulation.
A
distincttroughcan be identifiedon
thedorsal surfaceofthe retroar-ticular process.The
rightlowerjaw
isexposedin ventralviewbutshowslittlemorphologicaldetail. Its total length, including the retroarticular pro- icess, is47.5
mm.
1Posteriorto theskull, 16 cervicalvertebraeare
preservedin articulation.Arthaber(1924)counted 15 cervicals, but obviouslyomittedcounting the
poorlyexposedatlas. In dorsalviewthe cervicals
j
show
pachyostotic neural arches with alowneural Ispine; theventral surfaceofthecentrumis
orna-mented
with pairedlongitudinalkeels.The
sizeofthe cervical vertebrae slightly and gradually in-creasesfromfronttoback.
The
lengthofthecen-trum
is 4.2mm
in the axis, 4mm
in the 10th cervical,and
4.8mm
inthe16thcervial;thewidthofthe corresponding elements, measuredacross the zygapophysealarticulations, is5
mm,
6mm,
and
8mm,
respectively. Three cervical ribs (anincomplete
number)
can unequivocally beiden-tified,theanteriormostone of whichliesadjacent tothe 15th cervicalvertebra
and
carriesadistinct freeanteriorprocessclose toitsproximalarticular head.Separatedfromthecervicalvertebraebyagap, atotal of14 dorsalvertebrae (13 complete, 1 in-complete) arepreserved inarticulation, followed
by the first sacral vertebra.
The
dorsal vertebraeshow
aweak
constrictionofthecentruminventral view. Within the gap between the cervical anddorsalvertebrae series, 10(left)ribs are exposed,
plusthe spaceforoneadditionalribthatwasnot completely prepared. Followingthe 16 preserved
cervical vertebrae,thecontoursoffour additional elementscan beidentified (see alsoNopcsa,1928);
obt.f.
Fig.5. GastralribsofAnarosauruspumilio
Dames
(holotype)(drawnaftersmns59073).A, Anteriorgastral
ribs;B, posterior gastralribs. Scalebar
=
10mm.
all liein frontofthefirstexposedOeft) rib.
With
a total of 15 vertebralelements bridging thegap betweenthe preservedseriesofcervicaland
dorsal vertebrae,acountof45presacralvertebraeresults.Dames
(1890) counted 43 presacrals, Arthaber(1924) postulated48presacrals,and
Nopcsa
(1928)counted 39—40presacrals. Since theanteriormost ribexposedwithin thegap betweencervicals
and
dorsalsshowsthemorphology ofa holocephalous dorsal rib withoutfreeanterior process,
we
con-clude thatAnarosaunis hadno
more
than 19-20cervical vertebrae (in agreement with Nopcsa, 1928)
and
25-26 dorsal vertebrae.The
standard lengthwas
defined as the length ofthe last four presacral centra(Rieppel, 1989;Sander, 1989),andit is 23
mm
(the length of the second but last presacralcentrumis5.5mm;
thestandardlengthisthe
same
inthecastsfromGottingenandBerlin, butitis21.3mm
inthe castfrom Stuttgart).The
dorsal ribs lack pachyostosis.The
gastral ribs arecomposed
offive elements each (Peyer,quotedin Zangerl, 1935),the angled medioventral
elementbearing adistinctanteriortipat leastin the anteriorand middlepartofthegastral region,
and
two lateral elementson
either side (Fig. 5).The
ratio ofvertebral elements to gastral ribsis1:2.
Of
the three sacral vertebrae, only theanteri-ormost is preserved. However, three sacral ribs
canclearlybe identified,of whichthesecondone
(11.5
mm
long)isslightlylargerthanthefirstone(10,8
mm
long),whereasthethirdoneisonly in-completely preserved.The
sacral ribs are slightlyexpandedattheirproximalend.Nothingis
known
ofthecaudalaxialskeleton.
The
pectoral girdle is poorly preserved and/orexposed,and dermalelementscannot beidentified unequivocally.
The
right scapulais preserved in ventral view,possibly in association withafrag-FiG. 6. The pubis ofAnarosauruspumilio
Dames
(holotype). Scalebar=
5mm.
Abbreviation:obt.f, ob-turatorforamen.mentaryclavicle.
The
rightcoracoidlacksitsdistal portion, but showsa rounded proximal head as wellasastronglyconcaveanteriormarginoppos-ing the lessdistinctlyconcaveposteriormargin.
Of
the pelvic girdle,the ilium is not exposed.The
twoischiaarepreservedinarticulation.They
show
anarrow("rod-shaped" accordingtoNopcsa
[1928,p.31])acetabularportion, stronglyconvex anteriorand
posterior margins, anda widelyex-panded
ventralportion with aconvexventralmar-gin.
The
lengthoftheischiumis 19mm,
its prox-imalwidthis6.5mm,
theminimal widthis4mm,
and
thewidth ofthe ventral partis 20mm.
The
pubisoiAnarosaurusshows weakly concave an-terior
and
posteriormargins.The
obturatorfora-men
is not closed, but rather forms an open slitatthe posterior margin ofthe pubisclose tothe acetabularmargin ofthe bone.
The
concavityof thecentral(medial)margin ofthepubisresultsin adistinctspine protrudingfromtheanteroventral edgeofthepubis(Fig.6).A
largethyroidfenestrawas
developed between the pubis and ischium.The
length ofthepubisis 19.5mm,
its proximalwidthis 10
mm,
itsminimal width is 7mm,
the distal widthis9mm
excludingtheanteroventral spine,but 13mm
including the latter.The humerus
(Fig. 7A) is the only elementofthe forelimb that is preserved.
The
element isslightly curved with a concave medial(preaxial) margin, a well-developeddeltopectoralcrest,and a broadlyexpandeddistalportion.
The
entepicon-dylarforameniswellsetofffromthedistalartic-Table2. MeasurementsofthefemurofAnarosaurus pumilioDames,indifferentcastsofthe holotype.
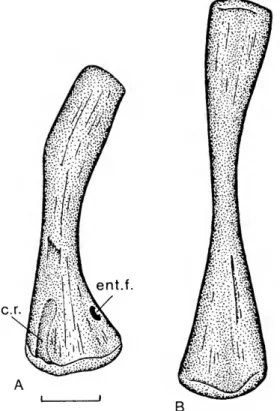
ec.r
nt.f.
Fig.8. Holotype ofAnarosaurusheterodontusn. sp.,
rightdentary (Institut fur Geowissenschaften, Martin-Luther-Universitat,Halle,M4/12).Scalebar
=
10mm.
share theformdiagnosticofthegenus
Anarosaums
(i.e.,aconstrictedtoothbaseandalanceolatetooth
crown).
Of
the preserved teeth, theanterior twoare distinctly larger (4.5
mm
high)than the suc-ceedingteeth (2.5mm
high). Preparation ofthe dentaryfrom the ventral side revealed a narrow symphysisasischaracteristicofpachypleurosaurs. Overallsize,toothmorphology, andtheincreased sizeoftheanteriordentaryteethrender thefossil closely similar toa lowerjaw
reported from the lower Muschelkalk of Winterswijk, erroneously referred to Cymatosaurus(Oosterink, 1986, Foto 40).A
perfectly preserved left premaxillafrom theSchaumkalk
of Freyburg/Unstrut (Fig. 9) has a dentition thatmatchesthemorphologydiagnosticofAnarosaurus.
The
premaxilla bears five teeth, ifa posterolateraltrough isinterpreted as afacet forthe maxillaratherthanas partofanalveolus forasixthtooth.Butevenwithsixfunctional teethon
the premaxilla thenumber
islessthanthe eight teethobservedon the premaxillaof Anarosauruspumilio.
The
premaxillafromtheSchaumkalk ofFreyburg/Unstrutisagainabout twicethesizeof
the premaxilla in the holotype of Anarosaurus
pumilio: thedistancefromthetipofthesnoutto the anterior margin ofthe external naris is 25.7
mm
inthefirst, 13.2mm
inthelatter.Interms ofsize
and
morphology(fivetooth po-sitions), the premaxilla from theSchaumkalk
ofFreyburg/Unstrut again comparesclosely to
ma-FiG. 9. Anarosaurus heterodontus n. sp., left pre-maxilla(InstitutfurGeowissenschaften, Martin-Luther-Universitat, Halle,M4/12).Scalebar
=
10mm.
Abbre-viations:mf, maxillaryfacet;np, nasal process; vp, vo-merineprocess.terialfromthelowerMuschelkalk ofWinterswijk,
which includesa skull depositedin the
Museum
Freriks, as well asa largeamount
ofundescribed material kept in private collections (Oosterink, 1986,Foto 38).The
skull in theMuseum
Freriks (#20778; Fig. 10) showsanumber
ofcharacters diagnostic ofpachypleurosaurs, suchasthe rela-tively shortandunconstrictedsnout,thelarge pre-frontal,the presenceofascleral ring,andthe rel-atively short postorbital portionoftheskull,and
it shares with other pachypleurosaurs from the lower Muschelkalk the deep
embayment
ofthecheekregion
and
the relatively large uppertem-poral fossa(as
compared
tothe Serpianosaurus-Neusticosaurus dade).Apart fromrelativesizeand
the heterodontdentition (enlargementofthe an-terior dentary
and
premaxillary teeth), the skull differs fromthe holotypeofAnarosauruspumiliobya closeapproximation ofprefrontaland post-frontal, restricting the frontal to a very narrow
entryintothedorsalmargin oftheorbit.
Schaum-Fig.10.
A
narosaurus heterodontusn. sp.,skull inleftlateralview,fromthelowerMuschelkalkof Winterswijk
(MuseumFreriks,#20778).Scalebar
=
20mm.
kalk (upperlower Muschelkalk) of
Freyburg/Un-strutandthatfromthelowermost Muschelkalk of
Winterswijk represent a
new
speciesof Anarosau-rus thatisgeologicallysomewhat
olderthanAn-arosauruspumilio from the lower middle
Mus-chelkalk. Morphological differences include ab-solutesize,the
number
ofpremaxillaryteeth,the relative sizeofthe anteriordentaryand
premax-illary teeth,
and
the configurationoftheprefrontaland
postfrontal bones. Itmay
be noted that the lowerMuschelkalkinthewesternpartoftheGer-manic
basin (Winterswijk) is geologicallysome-what
youngerthatthelowerMuschelkalkdeposits intheeastern partofthebasin (Freyburg/Unstrut),whichbringsthetwolocalititesclose in geological time.
Using Captorhiniis (Heaton, 1979)
Petrolaco-5aMms
(Reisz, 1981),C/aw^/osawms(Carroll,1981),and Youngina (Gow, 1975) as successive
out-groups(Storrs, 1991, 1993a), v4narc>5aMrusshows the following plesiomorphiccharacters: relatively large upper temporal fossae with both the post-frontal
and
the parietal broadly entering the an-terior and anteromedial margin ofthe fenestra; postorbital distinctlytriradiate;humerus
retaininganectepicondylargrooveinadditiontoan entepi-condylar foramen, and a well-developed delto-pectoral crest; dorsal ribs without pachyostosis; gastralribs
composed
offiveelementseach.Apomorphic
featuresof Anarosaurusincludethe spatulate expansion ofthe crown ofthe teeth, a character discussedand
figured in detailbyDames
(1890) but which is difficult to ascertain on the basisoftheavailable castsofthe holotypeof An-arosaumspumilio.
The
distinctionofcervicalfromdorsalvertebraeisdifficultin reptilesin general, aproblemaggravatedbythefactthatthe transition
is very poorly preservedin Anarosaurus.
Never-theless,acount of25-26dorsalvertebraeis
most
probable,anditisdistinctlyhigher thaninallother pachypleurosaursknown.
Both overallsize
and
the positionofthe ente-picondylarforamenatsome
distancefromthe dis-talarticularsurfaceofthehumerus
indicateama-ture individual, yetthe obturatorforamenin the pubis remains widely open.
The
pubis carriesa distinctanteroventral spineresultingfroma con-cavity ofits ventral (medial) margin, otherwiseknown
onlyinSerpianosaurusamong
otherpachy-pleurosaurs(Rieppel, 1989). Isolatedelements with
an open obturatorforamen and ananteroventral spinehavealsobeenreportedfromthebaseofthe
middle Muschelkalk
(Hagdom
8l Simon, 1993),and
aconcavelowermarginofthepubisisa gen-eral character for Simosaurus and Nothosaurusamong
eusauropterygians.CarrollandGaskill(1985,p.349; see also
Kuhn-Schnyder, 1959; Rieppel, 1993) considered the elongatedfemurdiagnosticofAnarosaurus. Mea-surements ofthe femurdifferon
different casts, but inspectionof Table2indicates that35.75mm
isanaverage estimateofthelengthofthefemur.
Using 23
mm
asstandardlength, the ratiofemur length/standard length is 1.55, an exceptionally high valueamong
all other pachypleurosaursknown
(Rieppel, 1993), corroboratingrelativefe-mur
length asa diagnosticfeatureofAnarosaurus(Table3).
The
corresponding valuesforother taxa are 0.86 for "Psilotrachelosaurus," 0.97-1.2 for Serpianosaurus(juvenilesandadults,bothsexes), 0.9 1-1.5forNeusticosauruspusillus (juvenilesandadults, bothsexes), 0.79-1.06 forNeusticosaurus peyeri(juveniles
and
adults,bothsexes),and 0.65-0.96 forNeusticosaurus edwardsii(juveniles and adults, bothsexes).The
distinct striationson
the surfaceofhumerus and
femurnearthe proximaland
distal heads form anothercharacteristic fea-ture ofthis genus (sometimes also observed onwell-preserved humeri and femora of Neustico-saurus: H.-D. Sues, pers. comm.). Ifthese stria-tions aretaken as indicatorsofadult individual age,they
document
that differences inoverallbody
size
and
degreeofbonesculpturing arenotmerelyontogeneticvariations inAnarosaurus pumilio and Anarosaurusheterodontus.
DactylosaurusGiirich, 1884
Type
SvEdES—
Dactylosaurus gracilis Giirich,1884.
Generic Diagnosis—
A
small pachypleurosaur with a narrow interorbital bridgeformed by the frontals;acloseapproximation ofthe postfrontaland
prefrontalalong thedorsalmargin oftheorbitdue toa long
and
slenderanteriorprocessofthe postfrontal (leavinga narrowentry ofthe frontal inthedorsalmargin oftheorbit);long butnarrow (kidney-shaped)upper temporalfenestrae;no
con-striction ofthe dorsal vertebral centra; clavicles withananterolateral expansion; preaxial margin ofthe radius with adistinctlyconcavemargin.The
humerus
is plesiomorphic at the level of the Pachypleurosauroidea butuniquewithinthegroup bythe retentionof anentepicondyledistinctlyset offfromthe shaft(insexy).Distribution—
Lower
Muschelkalk, central Europe.DactylosaurusgracilisGiirich,1884
1884 Dactylosaurusgracilis, Giirich,p. 125,PI. 2, Figs. 1, 2.
1886 Dactylosaurus gracilis, Giirich, pp.
457-458, textfig. 1.1886 Dactylosaurusgracilis, Deecke, p. 187.
1899 Dactylosaurusgracilis,
Schrammen,
PI.25, Fig. 10.1924 Dactylosaurusgracilis, Arthaber,pp.
483-484, textfig. 14.1928 Dactylosaurusschroederi, Nopcsa,pp. 3 1-37,PI. 3, Figs. 1-6.
1935 Dactylosaurusgracilis, Zangerl,p. 66.
1935 Dactylosaurusschroederi, Zangerl,p. 67.
1959 Dactylosaurusgracilis, Kuhn-Schnyder,p. 652.
1959 Dactylosaurusschroederi,Kuhn-Schnyder,
p. 652.
1985 Dactylosaurus gracilis. Sues
and
Carroll, pp. 1602, 1608.1985 Dactylosaurusschroederi.Sues
and
Carroll, pp. 1602-1608, textfigs. 1-3.1987 Dactylosaurusgracilis, Rieppel,pp. 1
109-1110.
1987 Dactylosaurus schroederi, Rieppel, pp.
1109-1110.
1993 Dactylosaurusgracilis, Rieppel,p. 10.
1993 Dactylosaurusschroederi, Rieppel, p. 10.
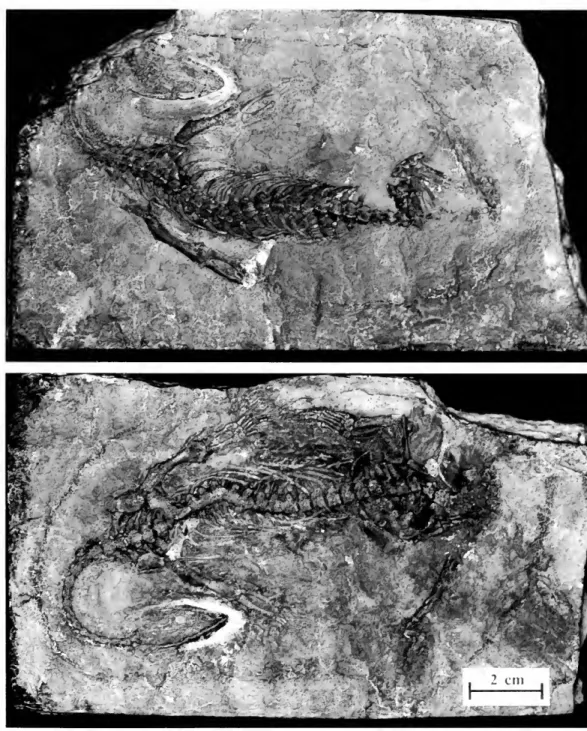
HoLOTYPE.—
Dactylosaurus graciliswas
de-scribed by Giirich (1884) on the basis ofa castmade
froma naturalmold
(Fig. 11)depositedatthe InstituteofGeologicalSciences,University of
Wroclaw (mgu
Wr
3871s).A
cast correspondingc
-a
S¥
3Fig. 11. Holotypeof DactylosaurusgracilisGiirich(mgu
Wr
3871s). Scalebar=
20mm.
tothe figuregiven byGiirich (1884, PI. II, Figs.
1
,2)iskeptattheNatural History
Museum,
Hum-boldt University,Berlin(mbR.63).
The
holotypecomes
from Michalkowitz,Upper
Silesia(now
Michalowice, Poland), assignedtothelowerMus-chelkalk.
The
holotype of Dactylosaurus schroederiNopcsa, 1928,
was
depositedinthecollectionsofthe Preussische Geologische Landesanstaltin Ber-lin(Kuhn, 1934),and
now
isinthecollectionsofthe Bundesanstalt fur Geowissenschaften
und
Rohstoffe, Berlin(uncatalogued)(Fig. 12).
A
rubbercastofthe specimen, describedbySuesandCarroll (1985), iskeptatthe Forschungsinstitut
und
Na-turmuseum
Senckenberg, Frankfurt a.M. (smfR-4097
a, b).The
holotypecomes
from deposits assignedtothelowerMuschelkalk ofGross-Stein,Upper
Silesia.The
holotypesofDactylosaurusgracilisas well asD. schroederi arebothnaturalmoldspreserved inyellowmatrix.The
colorofthe matrix,as well as thepreservationofvertebratefossilsasnatural molds,ischaracteristicoftheupperBuntsandstein (Rot)whichunderliesthelowerGogolin Beds(H.Hagdom,
pers.comm.).Referred
Material—
Institutund
Museum
fur Geologicund
Palaontologie, University of Tii-bingen, uncatalogued (Fig. 13), lowermost Mus-chelkalkof Gogolin(lowerGogolinbeds).A
poor-lypreserved specimenrepresentedbybarely vis-ibleimpressionsoftheskull,cervicalandanteriordorsal regionofthevertebral
column
preservedin dorsalvieworasfaintimpression(oftheventral aspect), right humerus, and left forelimb.The
specimenisslightlylargerthan theonedescribedby
Nopcsa
(1928).Nopcsa
(1928)and
SuesandCarroll(1985)refer toMeyer
(1847-1855)who
figuredhumericlosely similar tothoseofDactylosaurusgracilisfromtheMuschelkalk ("Saurier-Kalk") from Jena. Other humeri ofthis type are
known
from the lowerMuschelkalk ofBobreknearBeuthen (Bundesan-staltfurGeowissenschaften
und
Rohstoffe, Berlin,drawer S45/5 right) (Fig. 14),
and
fromthe Mus-chelkalk of Gogolin(Museum
fur Naturkunde, Berlin,mb
R. 772.1.,MB
R. 769,anduncatalogued;Institut
und
Museum
fur Geologicund
Palaon-tologie, University of Tubingen gpit 1744/1-10: Rieppel, 1993).Diagnosis—
Same
as forgenus,ofwhichthisisthe only
known
species.Locus
Typicus— Upper
Buntsandstein ofMichalowice,
Upper
Silesia(Gomy
Slask, Po-land).Distribution—
Upper
Buntsandsteinand low-erMuschelkalk, Europe.CoMMEt^TS—
Dactylosaurusgracilis is the type speciesofthegenus(Giirich,1884).The
cast avail-able in theNatural HistoryMuseum
in Berlinisofrather poorquality, and showslittle
morpho-logical detail.
The
following description of thespecimenisbased
on
anew
siliconcastmade
fromFig. 12. Holotypeof DactylosaurusschroederiNopcsa(bor,uncatalogued).
the holotypein 1994.Preservedare the posterior
part ofthe skull, the cervical vertebral column,
thepectoralgirdle,andthe right forelimb,all
ex-posedindorsalviewon thecast.
The
posterior partofthe skull showsa deeply excavatedcheekandrelatively large,kidney-shapedupper temporal fossae. Sutures are difficult to
identifyexceptforthe posteriorend ofthe post-frontal,whichdefinestheanteromedialmargin of
the upper temporal fossa. Sixteen cervical verte-brae(including theatlas)arepreservedin articu-lation; an additional vertebra
would
have beenpositionedattheleveloftheclavicles.Thiscount of17 cervical vertebrae correspondstothe
holo-Fig. 13. Dactylosaurus gracilis Giirich from the lower Muschelkalk of Gogolin, Upper Silesia
(Gomy
Slask, Poland)(gpit,uncatalogued).Fig.14. Dactylosaurus
gracilisGiirich,humerus,from thelower Muschelkalk ofBobrek near Beuthen (bgr, drawerS 45/5right).Scalebar
=
10mm.
type ofDactylosaurus schroederi Nopcsa, 1928. Cervicalribscanonlybeidentifiedalongthe pos-terior part ofthe cervical vertebralcolumn, and
they are oftypical structurewith a freeanterior process.
The
centra offouranteriordorsal verte-braearepreservedand
exposedinventralviewonthe actual specimen: they
show no
sign of con-striction, but slightly convex lateral margins in-stead.The
proximalpartofthe dorsalribs(as far as preserved)showsno
pachyostosis.In thepectoralgirdle,thedistalend ofthe clav-icleisseentooverlap with theanterior
and
medialaspectofthe scapulainatypical pachypleurosau-roid pattern.
The
interclavicula cannot beiden-tified.
The
clavicleshows expandedlateralcomers that arenotaspronouncedas inthe holotype ofDactylosaurusschroederi,possibly ajuvenile char-acter.
The
coracoids are represented by impres-sions only.Theirlengthis7.4(8)mm,
theirprox-imal widthis 3.6(3)
mm,
theirminimal widthis3.4(3.4)
mm,
and
theirdistal width is 4.6 (5.3)mm
(values inparentheses referto the rightele-ment;allmeasurements weretaken
on
the natural mold).The
coracoidsof bothsidesmeetventrally in a well-defined suture, but their proximalex-pansionisrelativelyweak,
and
theproximal mar-ginshowsno
notchindicatingthepositionofthemc
hu
Fig.15. DactylosaurusgracilisGmich,holotype(mgu
Wr
387Is),rightforelimb.Scalebar=
5mm.
Abbreviations: dc4,distalcarpal4;hu,humerus;in,intermedium; mc,, metacarpal 1;mcj, metacarpal5; ra,radius;ul, ulna;uln,ulnare.
coracoidforamen.This mightindicateincomplete ossificationofthecoracoidina juvenilesi)ecimen.
The
right frontlimbiswellpreserved(Fig. 15).The
righthumerus
isaslightlycurved elementthatshows hardly any morphological differentiation, as is characteristic ofjuvenile pachypleurosaurs (Rieppel, 1989; Sander, 1989).
The
deltopectoral crest is hardly differentiated,and
an entepicon-dylar foramen cannot be identified. In juvenile pachypleurosaurs, the entepicondylarforamenliesatthedistalmargin oftheossified partofthe
hu-merus
or,inevenearlier stages,beyond
that (Riep-pel, 1989;Sander, 1989).The
total length ofthehumerus
is 11.9mm,
its proximal width is 2.1mm,
itsminimal widthis 1.85mm,
and
itsdistalwidthis2
mm.
The
radiusisincompletelyexposeddue to the overlappingulna,but appearsslightly longer than theulna. Totallengthofthe radiusis
6.3
mm,
that ofthe ulnais 5.8mm.
Some
con-troversy surrounds the structure of the carpus (Giirich, 1886), but thenew
casttaken from the holotype corroboratesGiirich's(1886)descriptionofthree ossified carpalelements.
The
intermediumiselongatedandrectangularinshape;posterior to
itlietheroundedulnareanddistalcarpal4.
The
phalanges are incompletely preserved, probably
due to incompleteossification.
The
cast shows anumber
ofossified elements that correspond to Giirich's (1886) illustration of 2-3-3-4-3.Un-guals areunequivocallyidentifiedondigits 1 and
2 only; the count for the remaining digits
may
therefore be incomplete. Sander (1989)docu-mented
aproximo-distal sequence ofossificationofthephalangesinNeusticosauruspusillns.
The
overall sizeofDactylosaurusgracilis, and detailsof morphologythroughoutthe skeleton, in-dicate thatthe holotypeofthatsiieciesisa juvenilespecimen.
The
only characterpotentially contra-dictingthisconclusion istheadvanceddegreeof ossification inthecarpus,butvariation in the tim-ingofossificationcan beextensiveinextantrep-tiles(Rieppel, 1994a).
The
relativelylow count ofcervicalvertebrae(17)isshared with
Dactylosau-rus schroederi Nopcsa, 1928, and is otherwise
known
only from lower Ladinianpachypleuro-saurs{Neusticosauruspeyeri:Sander, 1989;
Neus-ticosaurus edwardsii: Carroll
&
Gaskill,1985).The
lattertaxaarecharacterizedbyarelativelysmaller
upper temporal fossathan is observedin Dacty-losaurusgracilis
and
D. schroederi.On
thebasisofthe limited data available,
we
conclude that Dactylosaurus gracilis Giirich, 1884 andDacty-losaurus schroederiNopcsa, 1928, are subjective
synonyms
(see alsoSues&
Carroll, 1985);thefirstname
haspriority.Morphological
Descrifhon—
The
morpho-logicaldescriptionoiDactylosaurusgracilisisbased
on
new
latexcastsmade
from the originalspeci-men
(natural mold) described byNopcsa
(1928, holotype ofDactylosaurusschroederi), as well ason
therubbercastskeptat theSenckenberg Mu-seum, Frankfurt.The
specimenisrepresentedbynatural molds ofpart
and
coimterpart.The
total lengthofthefossilmust
remainunknown
becauseofthe incompletely preservedtail,butit
must
haveexceeded213
mm;
snout-ventlengthis 158mm.
Skull length (the lengthoftheleftlowerjawramus
in ventral view) is 27
mm;
glenoid-acetabulumlength(measured on theright sideofthebody)is
68
mm;
standard length (length ofthe last four dorsal vertebral centra)is 15.5mm
(the lengthof thelastdorsal vertebralcentrumis3.5mm).
In the skull (Fig. 16), thesnoutcomplexisnot wellexposed.
The
nasalprocessofthe premaxillaremainsseparated from the frontalby thebroad
nasals,whichmeetalong a middorsalsuture.
The
frontalformsa shortanteromedial process enter-ingbetween the nasals, as well as short anterolat-eral processesthatmeet the maxilla
and
thereby separate the prefrontal from the nasal.The
pre-frontal reaches highup
along the anterodorsalslen-Fig. 16. Skullof DactylosaunisgracilisGiirich (bgr, uncatalogued). A,Drawnafteranewcastmadefromthe originalofNopcsa(1928);B,reconstructionofthe skullindorsalview. Scalebar
=
5mm.
Abbreviations:ar,articular,f,frontal;m,maxilla;n,nasal;p,parietal;pm,premaxilla; po, postorbital; pof, postfrontal;prf,prefrontal;q,quadrate;
so,supraoccipital;sq,squamosal.
der anterior process running along the postero-dorsal margin of the orbit.
The
prefrontal and postfrontal fail to meet, however, allowing the frontal toenterintothe dorsalmargin oftheorbit forashort distance.The
interorbitalbridgeformed bythepairedfrontalsisnarrow(2mm
inthelatex casttakenfromtheoriginalspecimen).The
fron-toparietal sutureis in a ratherforward position.The
posterolateralprocessesofthefrontalsextend posteriorlybetweentheparietaland
postfrontal.The
skull tableiswellexposed.The
uppertem-poral fossae are relatively large
and
ofanelongatedkidney shape.
The
length ofthe upper temporalfossais4.3
mm
(right)and
4.5mm
(left), respec-tively;thewidth oftheupper temporalfossais 1.5mm
(right) and 1.3mm
(left), respectively.The
postfrontal,thetriradiate postorbital, theparietal,and
thesquamosalallparticipate in theformationofthe margin ofthe upper temporal fossa.
The
pairedparietalsenclose thepinealforamen,which
ispositionedhalfway along the length ofthe pa-rietals.
The
occiputisdifficultto analyze.Inparticular, theoccipital exposure oftheparietal,and
its re-lation tothe squamosal, remainunclear.The
ventralview oftheskull displaysadermalpalateoftypicalpachypleurosauroidstructure;
lit-tlecan be addedtoitsdescription
and
illustrationbySues
and
Carroll (1985).The
pterygoidshows a thickening (arudimentary "transverse flange")along theanteriormargin ofthesubtemporal fos-sa.Thereis
no
positiveevidenceforthe presence or absenceofanectopterygoid.A
ceratobranchialelementispreserved, overlying theposterior part
oftheleftpterygoid.
Dactylosaunisgracilisshowsa totalof 36 pre-sacral vertebrae, ofwhich 17 are countedas
cer-vical elements (Sues
&
Carroll, 1985), character-ized by a keeled ventral surface ofthe centrum.The
ventral surfaceofthedorsalcentraissmooth,and
the centra are not constricted.The
neural archesare pachyostotic,and
the neural spinesare consistentlylow.Basedon
theSenckenbergcasts. Suesand
Carroll(1985) believed the neurocentral sutures tobeclosedinDactylosaunisgracilis,but they are quite distinct in the cervical, posterior dorsal(wherethey passbelowthe transverse pro-cesses), sacral,and
preserved caudal region (where they pass through the transverseprocesses).The
material doesnot allow the presenceofa neuro-central suture in theanterior dorsal region tobeascertained.
Sues
and
Carroll(1985) describe atlantal ribs,which aredifficultto identify, however.
The
cer-vical ribsbear adistinctfreeanterior process.The
dorsalribs arenotpachyostotic,and
thelastthree dorsal ("lumbar") ribs are distinctly reduced in length.Dactylosaunis gracilis shows three sacral ver-tebrae.
The
sacral ribs are broad, stoutelements with aweak
proximal expansion; theyarenot fused totherespective vertebrae.Caudalribs (againnot fusedtocaudalvertebrae)and
chevron bonesare notwellexposed. Gastralribsare likewisepoorly preserved, but whatlittle is presentsuggests that theyarecomposed
offiveelementseach,one me-dioventraland twolateralones oneitherside.Littlecan be addedtoSues
and
Carroll's(1985) descriptionofthe pectoralgirdle.The
right clav-icleshowsarounded andsomewhat
expanded an-terolateral edge (Fig. 17). This character is oth-erwiseknown
fromA
narosaurtdsonlyamong
otherpachypleurosaurs (undescribed material
from
Winterswijk), but is distinctly developed in the eusauropterygiangeneraNothosaurus and Simo-saurus (pers. obs.).
The
overlap ofthe clavicles with theanteriorand
medialsurfaceofthescapulaiswellexposedindorsalview
on
theright sideof the specimen.The
shape ofthe interclaviclere-mains
unknown. The
scapulaisofthetypical sau-ropterygian structure with a reduced and poste-riorlydirecteddorsalwing.The
coracoidforamenisdistinctandenclosedbetweenthenotched pos-teromedialmargin ofthe scapula
and
theopposingFig. 17. Pectoral girdle of Dactylosaunis gracilis
Giirich(bgr, uncatalogued),drawnafteranewcastmade
fromthe originalofNopcsa(1928). Scalebar
=
5mm.
Abbreviations: cl,clavicle;co, coracoid;co.f.,coracoidforamen;cr,cervicalrib;icl,interclavicle; sc,scapula.
posterolateral margin ofthe coracoid.
The
right coracoidmeasures 12.5mm
in length. Inthe pel-vicgirdle, the pubisshowsacomplete enclosureofthe obturator foramen by bone (Fig. 18); the
bone
is completely fused between the obturatorforamen andtheposterioredgeofthepubis.
The
pubis isa relativelybroad element with alengthof11
mm,
aproximal width of6.8mm,
aminimalwidth of5
mm,
andadistalwidth of7.8mm.
The
rightischiumiswellexposed,and showsa mark-edly thickenedproximal(acetabular) portion.
The
boneis12
mm
long,itsproximal widthis4.5mm,
itsminimal widthis3.5
mm,
itsdistalwidthis 13mm.
Suesand
Carroll (1985) describe awell-de-veloped supra-acetabularbuttress, butin ventral
viewthe iliumdoesnot displayitsacetabular por-tion; indorsal view; theleftiliumisconcealedby
obt.f.
Fig.18. Right pubis of Z)ac/>'/o5aMrus^rac/7wGurich
(bgr, uncatalogued),drawn aftersmns R-4907a. Scale bar
=
5mm.
Abbreviations: il.f.,facetfor ilium; is.f.,facetforischium;obt.f,obturatorforamen.
The humerus
ofDactylosaurus gracilisis verydistinctive(Fig. 19),at leastinsex
y
{sensu Riep-pel, 1989, andSander, 1989; seealso the discus-sioninRieppel, 1993).The
elementbears awell-developeddeltopectoralcrest distinctly setofffrom
the waisteddiaphysis.
The
distal headisbroadlyexpanded, with the entepicondyle forminga
me-dial process distinctly set offfrom the articular head.
The
entepicondylar foramen lies atsome
distance from the distal articulation.
The
distal articular heads are level.A
deep ectepicondylar ridgeispresent.The
radiusofDactylosaurusgracilisappearstobecharacterizedbyadistinctnotchinthemedial
(preaxial)margin oftheshaft(Fig.20A).Suesand Carroll(1985, p. 1607)attribute thisfeature toa preservationalartifact.Latexcaststakenfromthe originalspecimen
show
the concavityon
the pre-axial margin ofthe radius in both limbs,more
distinctly in their ventralexposure(and asimilar concavityisalso observedin theradiusofunde-scribedAnarosaurusmaterial fromWinterswijk).
ec
rec.r.
ent.f.
B
Fig. 19. Humerusof DactylosaurusgracilisGiirich (bgr,uncatalogued). A,Righthumerusinventralview, drawn after smns R-4907a; B, left humerus in dorsal view, drawnafter smns R-4907b. Scale bar
=
5mm.
Abbreviations: ec.r., ectepicondylarridge; ent, entepi-condyle;ent.f,entepicondylar foramen.The
ulnaissomewhat
shorterthan the radiusandbears a weakly developed olecranon on its
ex-panded proximalhead.
The
carpus (Fig.20A)
comprises threeundis-putedossifiedelements,viz.theintermedium,the ulnare,
and
the fourth distal carpal.The
inter-medium
is ofsubcircularshapeand
lies distaltothespatiuminterosseumbetweenradiusandulna.
A
pisiformisdistinct inNopcsa's(1928) retouchedphotograph ofthe specimen,
and
itspresence (or thatofaneomorph
initsposition)was
confirmed bySuesand
Carroll (1985).Latexcaststakenfromthe original sp)ecimen indicate the presence ofa
minutestructurenexttotheulnareintheleftlimb
only. Ifthat structure represents a carpal ossifi-cation, the pisiform (or a
neomorph) would
beextremely small
and
presentinone limbonly. In extantreptiles,the pisiformgenerallyisthelastofallcarpalelementsto ossify (Rieppel, 1992).
Ev-idencethatthisbone(ora
neomorph)
is, indeed.mc
B
Fig. 20. Manus and pes ofDactylosaurusgracilisGiirich (bgr, uncatalogued). A,Left forelimb, dorsal view, drawnaftersmnsR-4907b;B,righthindlimb, ventralview,drawnaftersmnsR-4907a.Scalebar
=
5mm.
Abbre-viations: as, astragalus;ca, calcaneum; dc4, distal carpal 4; dt4, distal tarsal 4; fi, fibula; in, intermedium; mci, metacarpal 1;mcj, metacarpal5;mt,,metatarsal 1;mt,,metatarsal5;pi,pisiform;ra,radius;ti,tibia;ul,ulna; uln, ulnare.present in Dactylosaurusgracilis is here consid-ered equivocal.
The
phalangealcountinthemanus
is 2-3—4-7-3.
The
presenceofeitherfourorfive phalangesinthefourthdigitcannot beascertained unequivocally.Measurements
ofthehumerus
are giveninTable 4.Only
the proximal head ofthe left femur andthe distal head ofthe right femur are exposed.
Theseindicatea slenderbonewithtibialand
fib-ular articfib-ularheadslevel.
The
right fibulaisonly partiallyexposed.The
righttibiaisalittlesturdierthanthefibulabutitis stillaratherslenderbone
Table5. Measurementsofthelimbbonesof Dactylosaurusgracilis(holotype ofDactylosaurusschroederiNopcsa; BGR,uncatalogued).
with astraightmedial,
and
aslightlyconcave lat-eral,margin.Threeossifiedelementscanbe iden-tifiedinthetarsus(Fig. 20B), viz. theastragalus,calcaneum, andthe fourth distal tarsal.
The
pha-langealformulainthe pesis2-3-7-5—4,five pha-langes in the fourthdigit representingtheplesio-morphic condition.
Measurements
of the limb bones ofthe specimen described byNopcsa
are givenin Table 5.Discussion— The body
proportions of Dacty-losaurusgracilisfall squarely within therangeof variability oftheMonte
SanGiorgio pachypleu-rosaurs (Table 3).The
upper temporal fenestraeare distinctlysmallerthan theorbit,yet distinctly larger than the upper temporal fenestrae in Ser-pianosaurus
and
Neusticosaurus.As
inAnarosau-rus, the postfrontal, postorbital, and parietal broadly enter the margin ofthe upper temporal
fossa (unlike inSerpianosaurus
and
Neusticosau-rus), but the upper temporal fenestrae in Dacty-losaurusgracilisarerathernarrowand
elongated asopposedtothesubcircularcontoursoftheupper temporal fenestra in Anarosaurus.As
inAnaro-saurus, theupper temporalarchof Dactylosaurm
isnarrow
and
thecheekdeeply excavated, unlike Serpianosaurusand
Neusticosaurus, whichshow
abroadenedpostorbitaland
squamosal. Like oth-erpachypleurosaurs,Dactylosaurusshowsarather anterior positionofthefrontoparietal suture,with distinct posterolateralprocessesofthefrontals ex-tendingbackward between the parietal and post-frontal. Dactylosaurusgracilis differs from other pachypleurosaurs exceptKeichousaurusinthenar-row
interorbitalbridgeformed bythefrontals;the close approximation ofthe prefrontaland
post-frontalbonesalong thedorsalmargin ofthe orbitissharedwithundescribedAnarosaurusmaterial
from Winterswijk.
The
lattercharacteris associ-atedwith thedevelopment ofa longandslender anteriorprocessofthe postfrontal.The
cervical vertebral count in Dactylosaurus gracilis is lowcompared
to other pachypleuro-saurs, particularlythoseofthelowerMuschelkalk{Anarosaurus: 19-20[see above]; Keichousaurus: 26;Serpianosaurus:714-18[Rieppel,1989];
Neus-ticosaurus: 15-20[Carroll&.Gaskill, 1985;
Sand-er, 1989],but the significanceofthis characteris
difficult to assessbecause ofthe lack ofdata
on
intraspecificvariability.Vertebrae aredifficultto
countintheTubingen specimen ofDactylosaurus gracilis,butthere
may
beasmany
as 19cervicals (and 17 preserveddorsals).The
morphology ofthehumerus
of Dactylosau-rusisquite distinctive.Nopcsa
(1928;seealsoSues&
Carroll, 1985,p. 1607)foundit similar to iso-latedhumeri fromthelowerMuschelkalkreferred to "Macrotracheli"by H.v.Meyer
(1847-1855).Huene
(1942, Fig. 11) figured ahumerus
frommuch
youngerdeposits(lowermostLettenkohleof SchwabischHall,southernGermany), whichSuesand
Carroll(1985,p. 1607) againfoundsimilar to that ofDactylosaurus, but the Lettenkohlespeci-men
appearsmore
curved ratherthan angulated as in thehumerus
ofDactylosaurus, andthe ent-epicondyleissetoff"fromtheshaftbyacrackrather than a notch. Indeed, the specimen figured byHuene
(1942) closely resembles thehumerus
of Neusticosauruspusillus (Sander, 1989, Fig. 15c),and
doesnotindicatethe extensionofthe strati-graphic occurrence ofDactylosaurus beyond the lower Muschelkalk.The humerus
ofDactylosaurus gracilis (sexy
sensu Rieppel, 1989; Sander, 1989) is
plesio-morphicwithrespect toallother pachypleurosaurs
intheretentionofawell-differentiated entepicon-dyleandadistinctectepicondylar groove.
The
ju-venile specimen describedby Giirich (1884) in-dicates similar ontogenetic changes in the
mor-phologicaldifferentiationasthose
known
inother pachypleurosaurs(Rieppel, 1989; Sander, 1989), while aseriesofhumeri fromthelowerMuschelk-alk of Goglin,
Upper
Silesia(GPIT
1744/1-10) indicatesa similarsexualdimorphism
in Dacty-losaurusgracilisasisknown
forother pachypleu-rosaurs(Rieppel, 1989; Sander, 1989).As
inother pachypleurosaurs, the index of minimal width/ distalwidth ismost
informativewith respect to ontogeneticchangesandsexualdimorphism
inthe differentiationofthe humerus. Plottingall avail-ablematerial(seereferredmaterial above)results inanindexof0.53-0.67forsexx
and/orjuveniles,and0.40-0.57foradultspecimensofsexy.
Over-lapofthisratio(intherange of 0.54-0.57)between sexes isrestricted to three specimens (mb R. un-catalogued, gpit 1744/1-2), and
may
reflect nat-ural variability as well aswearoftheectepicondyle duringpostmortemtran