JOURNAL
OF
RANGEMANAGEMENT
Published bimonthly-January, March, May, July, September, November
Copyright 1980 by the Society for Range Management
Managing Editor FLOYD E. KINSINGER
2760 West Fifth Avenue Denver, Colo. 80204 Technical Editor REX D. PIEPER
Dept. of Animal 8 Range Sciences New Mexico State University Las Cruces, N.M. 88003 Book Review Edltor RICHARD E. FRANCIS
Rocky Mtn. Forest & Range Experiment Station 240 West Prospect Fort Collins, Colo. 80521 Production Editor PATRICIA G. SMITH
2760 West Fifth Avenue Denver, Colo. 80204 Editorial Board 1978-80
DONALD A. KLEBENOW, Reno, Nev. JAMES T. NICHOLS, North Platte, Neb. MICHAEL D. PITT, Vancouver, B.C. RONALD E. SOSEBEE, Lubbock, Tex. 1979-81
ROD BOVEY, College Station, Tex. NEIL FRISCHKNECHT, Provo, Utah MARTIN VAVRA, Union, Ore. HAROLD GRELEN, Pineville, La. 1980-82
HENRY MAYLAND, Kimberly, Ida. JOHN MENKE, Davis, Calif. STEPHEN SHARROW, Corvallis, Ore. PHILLIP URNESS, Logan, Utah
TABLE OF CONTENTS:
Vol. 33, No. 5, September 1980
ARTICLES
323328
331
337 340
34s 349
354 360
363
367
371
374
379
INDIVIDUAL SUBSCRIPTION is by membership in 381 the Society for Range Management.
LIBRARY or other INSTITUTIONAL SUBSCRIP- TIONS on a calendar year basis are $56.00 postpaid 385 to all countries. Payment from outside the United States should be remitted in US dollars by interna- tional money order or draft on a New York bank. 388 BUSINESS CORRESPONDENCE, concerning sub- scriptions, advertising, reprints, back issues, and 390
related matters, should be addressed to the Manag- ing Editor, 2760 West Fifth Avenue, Denver, Colo.
80204. 394
EDITORIAL CORRESPONDENCE, concerning manuscripts or other editorial matters, should be
Drought Effects on Adventitious Root Development in Blue Grama Seedlings by D.D. Briske and A.M. Wilson
White-Tailed Deer Densities and Brush Cover on the Rio Grande Plain by Allen A. Steuter and Henry A. Wright
The Effects of Subsurface Irrigation on Current and Subsequent Year’s Growth in Shadscale by Patricia S. Johnson and Brien E. Norton
Simulated Cattle Injury to Planted Slash Pine: Girdling by Clifford E. Lewis Simulated Cattle Injury to Planted Slash Pine: Combinations of Defoliation, Browsing, and Trampling by Clifford E. Lewis
Simulated Cattle Injury to Planted Slash Pine: Defoliation by Clifford E. Lewis Susceptibility of Selected Woody Plants to Pelleted Picloram by Lynn M. Kitchen, C.J. Scifres, and J.L. Mutz
Feral Horse Demography: A Preliminary Report by Michael L. Wolfe, Jr. Relating Fire Occurrence to Weather Conditions on the Great Basin Rangelands by Kenneth G. Hubbard
Effects of a Fall Wildfire on Herbaceous Vegetation on Xeric Sites in the Selway- Bitterroot Wilderness, Idaho by Evelyn H.L. Merrill, Henry F. Mayland, and James M. Peek
Diameter-Age Relationships of Two Species of Mountain Mahogany by Jack D. Brotherson, James N. Davis, and Larry Greenwood
Factors of Influencing Microhistological Analysis of Herbivore Diets by M. Vavra and J.L. Holechek
Seasonal Differences in the Element Content of Wyoming Big Sagebrush by Larry P. Gough and James A. Erdman
Herbage Yields from a Clayey Range Site 10 and 11 Years after Severe Renovation by Frank Rauzi
Strains of Blue Grams and Sideoats Grama Evaluated for the Southern Great Plains by William D. Pitman and Chester C. Jaynes
An Evaluation of Anti-Coyote Electric Fencing by Michael J. Dorrance and John Bourne
Quality, Yield, and Survival of Asiatic Bluestems and an Eastern Gamagrass in Southern Illinois by J.J. Faix, C.J. Kaiser, and F.C. Hinds
Nonstructural Carbohydrates in Roots of Gambel Oak Sprouts Following Herbi- cide Treatment by David M. Engle and Charles D. Bonham
Growth Patterns and Biomass Relations of Xanthocephalum sarothrae (Pursh) Shinners on Sandy Soils in Southern New Mexico by Samu Nadabo, Rex D. Pieper, and Reldon F. Beck
addressed to the Technical Editor, Dept. of Animal
and Range Sciences, New Mexico State University, TECHNICAL NOTES
Las Cruces, N.M. 88003. 398
INSTRUCTIONS FOR AUTHORS appear on the
A Fecal Collection Apparatus for Deer Nutrition Studies by John C. Malechek, inside back cover of each issue. Kenneth 0. Fulgham, and Michael A. Smith
THE JOURNAL OF RANGE MANAGEMENT (ISSN BOOK REVIEWS 0022-409X) is published six times yearly for $28.00 400
per year by the Society for Range Management,
Grasses of Louisiana by Charles M. Allen 2760 West Fifth Avenue, Denver, Colo. 80204.
Drought Effects on Adventitious Root Devel-
opment in Blue Grama Seedlings
D.D BRISKE AND A.M. WILSON
Abstract
Crowns of blue grama (Bouteloua gracilis) seedlings of three ages were exposed to drought treatments for 2 days, in constant humidity environments, and were then planted in moist soil for a lo-day growth test at 25°C. Percentage survival of crowns decreased with a decrease in water potential during the temporary drought treatment and with a decrease in crown age at time of treatment. The percentage survival rates of 21-, 28-, and 35-day-old crowns treated at -180 bars were 5, 54, and 83, respectively. Crowns exposed to the 2-day drought treatment subsequently produced shorter adventitious roots than untreated crowns. Thus, a drought-induced inhibition of adventitious root growth may reduce the probability of successful seedling establishment.
Establishment of blue grama (Bouteloua gracih) seed- lings begins with germination and extension of the seminal primary root, which originates from the lower portion of the embryo (Gould 1968). After approximately 2 weeks of favorable environmental conditions following emergence, seedlings develop the capacity for adventitious root devel- opment from crowns of the primary shoot and tillers located at or very near the soil surface (Hyder et al. 197 1; van der Sluijs and Hyder 1974). If seedlings do not initiate and develop adventitious roots within 6 to 10 weeks after emer- gence, they often die. Seedling death at this time does not result from an inherent limit to longevity of the seminal primary root; rather, seedlings expand leaf area to a maxi- mum that can be supported by water from the seminal root and then succumb to an increase in transpiration stress (Wilson et al. 1976). Adventitious roots are effective in seedling establishment because they are longer-lived and have a greater capacity for water uptake than the seminal primary root. Thus, successful adventitious root establish- ment provides seedlings with an effective means of drought avoidance.
In the more arid portions of the Central Plains, blue grama plantings often fail because the environmental requirements for development of adventitious roots are not met. These requirements include a moist soil profile at the time of planting, temperatures between 15 and 30°C, and two periods in which the soil surface remains continuously moist for 2 to 4 days-one period for emergence and one for development of adventitious roots (Briske and Wilson 1977, 1978; Fults 1944; Olmsted 1941, 1942; Riegel 1941; Wilson and Briske 1979).
The authors are, respectively, former graduate research assistant, Department of Range Science, Colorado State University, and plant physiologist, U.S. Department of Agriculture, Science and Education Administration, Agricultural Research, Crops Research Laboratory, Colorado State University, Fort Collins, Colorado 80523. Briske currently is assistant professor, Department of Range Science, Texas A&M University, College Station, Texas 77843.
The study involved cooperative investigations of the Agricultural Research, Science and Education Administration, U.S. Department of Agriculture, and the Colorado State University Experiment Station, Fort Collins 80523 (Scientific Series Paper NO. 2413). The authors thank Michael A. Orrino, former biological aid, for photographs of seedlings and assistance in plant measurements. We thank John A. Dickerson, of the Soil Conservation Service, for providing the blue grama seed used in the study.
Manuscript received February 23, 1979.
JOURNAL OF RANGE MANAGEMENT 33(5), September 1980
In addition to the direct effects of environment, morpho- logical stage and physiological condition may influence the
capacity of seedlings to initiate and extend adventitious roots. On the Central Plains, seedlings surviving on only the seminal root are often exposed to periods of drought, which may adversely affect their physiological condition. The objective of this research was to determine the capacity of seedlings to prod&e adventitious roots as affected by age and exposure to temporary drought. This capacity may determine whether or not seedlings will become successfully established once the rather restrictive environmental requirements for adventitious root initiation and elongation are met.
Methods
Blue grama accession PM-K-1483 is a synthetic blend of seed from 12 accessions originating in southern Kansas and Texas. Seed for this study was harvested in 1974 at the Plant Materials Center, Manhattan, Kansas.
Seeds were planted at a depth of 2 cm in plastic pots (15 cm diameter by 15 cm deep) filled with autoclaved sandy loam soil. Planting of seeds was scheduled so that seedling crowns of three age classes (2 1, 28, and 35 days) would be ready for temporary drought treatment on the same date. During initial seedling growth in the greenhouse, the pots were subirrigated to promote growth of the seminal primary root and to maintain a 1.5-cm layer of dry surface soil that would prevent the initiation of adventitious roots. After emergence, seedlings were thinned to 12 per pot.
At 2 1,28, and 35 days after planting, the seminal root system and subcoleoptile internode were removed just below the coleoptilar node. All leaf tissue was removed 1 cm above the coleoptilar node to simulate a drought-induced die-back of all exserted leaf tissue and to facilitate placement of crowns within the constant humidity tray. These procedures were accomplished to accurately control crown drought stress. Uniform stress generally is not achieved through drying soil in the entire rooting zone of the seminal root. Therefore, only the seedling crown (lowest l-cm section of stem base) was retained to evaluate drought tolerance.
Seedling crowns were exposed to temporary drought for 2 days in constant humidity trays (Wilson 1971). Water potentials of -30, -60, -90, - 120, - 150, and - 180 bars were maintained in the trays with various concentrations of NaCl solution (Lang 1967; Robin- son and Stokes 1949). Seedling crowns lost water vapor to the constant humidity air but did not touch the NaCl solutions. The capillary flow of NaCl solution above and below the constant humidity tray was sufficient to compensate for evaporation from the moist paper.
After 2 days of exposure to drought, seedling crowns were planted at a depth of about 0.7 cm in plastic pots (15 cm diameter by 15 cm deep) filled with autoclaved sandy loam soil. Thus, the upper portion of the crowns extended about 0.3 cm above the soil surface. The soil was covered with a 0.5-cm layer of fine gravel to reduce evaporation from the soil surface. The soil was irrigated daily by surface application to maintain favorable moisture condi- tions (about -0.3 bars) for the growth of leaves and adventitious
roots from treated crowns. After the planting of crowns, pots were placed in a growth chamber at a constant temperature of 25OC, daylength of 15 hours, relative humidity of SO%, and photosyn- thetic photon flux density of 480 microeinsteins mZ2 set-‘. The experiment also included a control consisting of 21-, 28, and 35-day-old crowns that had not been exposed to temporary drought (untreated). The seminal root system and leaves were removed just as in the treated crowns, but they were then imme- diately planted on the same date as the treated crowns.
After a IO-day growth performance test, all pots were removed from the growth chamber, soil was washed from the roots with a fine spray of water, and seedlings were placed in a 10% solution of acetic acid and stored at 5°C until measurements of growth could be made. The number of shoots (primary shoot and all tillers) that had developed before the drought treatment, the number of shoots that produced regrowth during the lo-day test, and the number of adventitious roots were counted. The lengths of all leaf blades and adventitious roots were also measured. New shoots, leaf blades, and adventitious roots were counted and measured if they had reached lengths of 0.5, 0.5, and 0.1 cm, or more respectively. Percentage survival of treated and untreated crowns was based on the number of crowns in which one or more shoots exhibited leaf growth of at least 0.5 cm during the lo-day growth test under favorable temperature and moisture conditions. The number of surviving 21-day-old crowns in the -180 bar treatment was inade- quate for an evaluation of drought effects on selected growth measurements. Consequently, only crown survival was compared for all three age classes at the -180 bar drought treatment.
A randomized complete block experimental design with six replications was used. The six replications were six sequential repetitions of the experiment. Each value (within a replication) for seedling survival was based on a sample of 15 treated or untreated crowns planted in an individual pot. Each value for the other growth criteria was based on the average of all surviving seedlings from one pot. Analysis of variance, linear regression, and poly- nomial regression were used to evaluate relationships among age classes, drought treatments, and growth responses. All observa- tions were included in statistical analyses, but only means were reported.
Additional seedlings (21,28, and 35 days of age) were grown for measurement of xylem water potential with a pressure chamber (Waring and Cleary 1967). The main shoot and large tillers were used for these measurements. The objective was to determine seedling water potential before harvest compared with crown water potential during the 2-day drought treatment.
Extra seedling crowns (35 days old) were exposed to water potentials of -30, -60, -90, -120, -180 bars in constant humidity trays to determine if they had reached equilibrium after 2 days of treatment. The water potential of these crowns was measured with a thermocouple psychrometer. Because standard procedures are not useful for measuring water potentials below about -60 bars, a modified procedure was used (Campbell and Wilson 1972). The thermocouple junction was cooled for 10 minutes over a sample of NaCl solution (-10 bars) to allow water condensation on the junction. The sample of seedling crowns was then quickly positi-
oned below the junction, and the thermocouple output (micro- volts) was recorded. A similar procedure was used with standard NaCl solutions for developing a calibration curve. Measurements of crown water potentials were made for three of the replications.
Results
Seedling and Crown Water Potential
Before harvest and treatment of crowns, 21-, 2%, and 35-day-old blue grama seedlings exhibited average xylem water potentials of -9.5, -14.5, and -17.0 bars, respectively. Thus, crown water potentials in the least severe drought treatment were about 13 to 20 bars lower than xylem water potentials of seedlings before harvest.
Water potential of soil in which the seminal root was
growing was kept at about -0.3 bars. Therefore, differences in seedling water potential were not due to differences in soil moisture conditions, but were probably due to greater leaf area and transpirational water loss in older seedlings than in younger seedlings. Wilson et al. (1976) observed that an increase in leaf area at a time when there was little or no increase in capacity for water uptake created a condition of seedling drought, even though the seminal root was growing in moist soil. Apparently, water uptake was restricted by a threadlike nature of the subcoleoptile internode connecting the seminal primary root and the shoot.
Thermocouple psychrometer measurements indicated that seedling crowns had reached equilibrium with constant humidity environments during the 2-day treatment (Table
1). Because water potentials represent average values for individual crowns, the possibility of minor water potential gradients within the crown has not been eliminated. Shoot apices and adventitious root primordia of large 35-day-old crowns might have been protected from dehydration more than the apices and primordia of comparatively small 21- day-old crowns. However, the close agreement between solution osmotic potential and average crown water potential suggests that differences among age classes in crown injury and survival resulted mainly from differences in drought tolerance rather than drought avoidance (Levitt 1964).
Table 1. Osmotic potentials of controlling solutions (bars) and water potentials for 35-day-old crowns for each of the six drought treatments after a 2-day equilibration period in constant humidity trays.
Solution osmotic potentials Crown water potentials -30
-60 -90 -120 -150 -180
l&indicates standard deviation of the sample.
-3Of2.1’ -62f 1.5 -87f7.9 -116f7.5 -152f7.5 -1801t9.2
20 DAYS --o- 35 DAYS --A- -
-?*o -150 I -120 I -90 1 -60 1 -30 1 0 I
DROUGHT TREATMENT (bars)
Fig. 1. Effects of age and temporary drought treatments on thepercentage of blue grama crowns that produced regrowth during a IO-day growth test underfavorable soil moisture conditions. Correlation coefficients (r) labeled with two asterisks p*) are significant at the 0.01 level. SEE = standard error of estimate.
Table 2. Number of shoots per crown of 35-, 2%, and 21-day-old blue grama seedlings, before temporary drought treatments; and number of shoots per crown producing regrowth during a lo-day growth test following temporary drought treatments.
Growth and criteria and
crown age class -180 -150 -120
Temporary drought treatments (bars)
-90 -60 -30 0 Mean
Number of shoots/crown before treatment
35 days 28 days 21 days Treatment Mean
Number of shoots/crown producing regrowth
35 days 28 days 21 days Treatment Mean
6.3’ 6.1 6.3 6.6 6.9 6.4 6.6 6.5 c
3.9 4.5 4.2 3.9 4.5 4.2 4.4 4.2 b
2.6 3.1 2.7 2.8 2.8 2.8 2.7 2.8 a
4.3 a 4.6 a 4.4 a 4.4 a 4.7 a 4.5 a 4.6 a
3.5 4.0 4.2 4.8 5.0 4.8 5.5 4.5 c
2.1 2.6 2.5 2.8 3.1 3.0 3.3 2.8 b
1.0 1.6 1.2 1.6 1.7 2.0 2.0 1.6 a
2.2 a 2.7 c 2.6 b 3.1 cde 3.3 de 3.3 se 3.6 e
IValues represent an average of six replications. Means in columns or rows, within each growth criterion, labeled with the same letter are not significantly different at the 0.0 I
level.
Effects of Temporary Drought on Survival
Percentage survival of crowns exhibited a significant (P<O.Ol) age times drought interaction, indicating that crowns of younger seedlings were more susceptible to drought injury than crowns of older seedlings (Fig. 1). Per- centage survival rates of treated (-180 bars) and untreated crowns were 5 and 86 for 2lday-old crowns, 54 and 95 for 28-day-old crowns, and 83 and 99 for 35day-old crowns, respectively. Drought tolerance of 28day-old crowns approached that of 35day-old crowns but had not yet deve- loped sufficiently for a high percentage survival in the severe drought treatments. Twenty-one-day-old crowns had deve- loped neither the capacity for regrowth (control treatment) nor the drought tolerance exhibited by the 28- and 35-day- old crowns.
Seedlings randomly assigned to various treatments did not differ in number of shoots per crown before harvest and treatment (Table 2). The number of shoots that survived the drought treatments and produced regrowth during the lo- day test decreased with a decrease in water potential during temporary drought treatments.
Effects of Temporary Drought on Growth
Leaf Development and Growth
Leaf growth resulted mainly from the expansion of imma- ture leaf blades that had been clipped at the time crowns were harvested; it resulted, to a lesser degree, from the initiation of new tillers from axillary buds. Twenty-six per- cent of the crowns produced one new tiller, and only rarely did crowns produce two new tillers. Data indicated that both leaf intercalary meristems and axillary buds possessed a high degree of drought tolerance.
Total leaf length produced during the IO-day test decreased with a decrease in water potential during the 2-day drought treatment (Fig. 2). Total leaf length produced bY 21-, 28-, and 35-day-old-crowns was reduced 56,35, and 30%, respectively, as a result of treatment at -150 bars. Number of surviving shoots per crowns was positively asso- ciated with total leaf length produced per crown (r = 0.96). Adventitious Root Development and Growth
Adventitious roots were not initiated in crowns of any age class until several centimeters of leaf had developed. Thus, leaf tissue was the first regrowth observed. These results agree with those of Hyder et al. (1976), who found that the
capacity for root growth in mature blue grama crowns was more sensitive to drying than the capacity for shoot growth. Only 3% of the crowns produced leaf growth but no root growth in this study.
The number of roots per crown produced during the IO-day test decreased with a decrease in age of crowns at time of treatment (Fig. 3). The large number of roots pro- duced by older crowns was mainly a result of the large number of surviving shoots from which adventitious roots could develop.
Number of roots per crown also tended to decrease with a decrease in water potential during the temporary drought treatments. In relation to the control treatment, the number of roots in 21-, 28-, and 35-day-old crowns was reduced 24, 8, and S%, respectively, as a result of the - 150 bar treatment. Only in the 21day-old crowns was the reduction in root numbers significant (KO.05).
Total length of adventitious roots per crown decreased with a decrease in age of crowns at the time of drought treatment (Fig. 4 and 5). Greater total length of roots in the older crowns can be attributed both to a greater number of
50
r
s
I
O- I 1 I 1 35, WIYS --o-- -180 -150 -120 -90 -60 -30 0
DROUGHT TREATMENT (bars)
Fig. 2. Effects of age and temporary drought treatments on total leafblade length produced by the surviving blue grama crowns during a IO-day growth test under favorable soil moisture conditions. Correlation co- efficients (r) labeled with two asterisks p*) are significant at the 0.01 level. SEE = error of estimate.
IO -
26 A ________--
g -________-&.--- ~ ~
‘1
5 p 6-
E , = 0.16 SEE = *_ 1.0 ro;t 0
k 4’31 - .__~____-__- 0
:
3
r= 0.40* SEE’ ? 0.5 o rOOt
0
z 2- 0 AGE CLASS
‘21 DAYS + 20 DAYS -m- 35 DAYS --A- -
21 days
-180 bars
21 days
Untreated
-7eo
-150 -120 -90 -60 -30 0DROUGHT TREATMENT (bars)
shoots from which adventitious roots could be produced and to a greater length per root.
Total length of adventitious roots per crown also decreased with a decrease in water potential during the temporary drought treatments (Fig. 4). Total root length produced from 21., 2X-, and 35.day-old crowns was reduced 52, 34, and 27%, respectively, by the -150 bar treatment. Number of live shoots per crown was positively associated with total length of roots per CTOW~ (I = 0.94).
The length of the longest root per crown decreased wifh a decrease in age of crowns at the time of treatment (Fig 6). The maximum rate of root elongation (averaged over the entire IO-day growth test) was about 1.0 cm per day and occurred in 35-day-old, untreated crowns. This is a low rate
2, DAYS - 28 DAYS -m- 35 DAYS --A--
_qKl
I
-I 50 -120 -90 -60 -30 0 DROUGHT TREATMENT (bars1\ \
28 days
‘\ 28 days
-180 bars
,ptreoted
of elongation, compared with an elongation rate of 2.3 cm per day found in a previous experiment (Briske and Wilson
1977). The low rate of root elongation was probably a result of the long period (2 to 6 days) required by seedling crowns to produce photosynthetic tissue. Crowns required several centimeters of leaf length before the initiation and develop- ment of adventitious roots.
Ih roots because of rapid drying of the soil surface. Even under conditions in which seedling-drought is not severe, the initial rate of root extension is sometimes just sufficient to keep the root tip ahead of the drying soil front (Wilson and Briske 1979). Thus, a drought-induced delay of root initiation and an inhibition of root growth may reduce the probability of
successful seedling establishment. Injury and death of seed- lings might be reduced by conserving soil moisture and by planting when the soil profile is moist and the probability of precipitation and favorable temperatures is high.
On the basis of leaf and root growth following temporary drought, the results indicate that drought tolerance of blue grama seedlings increases with increasing age. However, both the mechanism by which drought tolerance increases
1
:j , , ijiii~
, ,
furtherinvestigation. and the stage of seedling tolerance reaches a maximum development remain unknown at which drought and warrant-180 -I 50 -120 -90 -60 -30 0
DROUGHT TREATMENT (bars) Literature Cited
Fig. 6. Effects of age and temporary drought treatments on the longest Briske, D.D., and A.M. Wilson. 1977. Temperature effects on adventitious adventitious root produced by blue grama crowns during a IO-day root development in blue grama seedlings. J. Range Manage. 30. 276- growth test under favorable soil moisture conditions. Correlation co- 280.
efficients (r) labeled with two asterisks (**) are significant at the 0.01 Briske, D.D., and A.M. Wilson. 1978. Moisture and temperature require- level. SEE = standard error of estimate. ments for adventitious root development in blue grama seedlings. J.
Range Manage. 31: 174-178.
Campbell, G.S., and A.M. Wilson. 1972. Water potential measurements of of the -150 bar drought treatment. The relatively small soil samples, p. 142-149. In: R.W. Brown and B.P. Van Haveren (ed.) percentage decrease iniength of roots produced by %-day- Psychrometryin Water Relations Research. Utah Agr. Exp. Sta., Logan. old crowns is similar to the results of Hyder et al. (1976), Fults, J.L. 1944. Some factors affecting the establishment of perennial grass who found that root lengths of mature blue grama crowns for erosion 615-625. control in eastern Colorado. J. Amer. Sot. Agron. 36: were not affected by drying treatments. The crowns of Gould, F.W. 1968. Grass Systematics. McGraw-Hill Book Co., New York. mature blue grama plants-and of older seedlings apparently 382 p.
are more drought tolerant than crowns of younger seedlings. Hyder, D.N., A.C. Everson, and R.E. Bement. 1971. Seedling morphology and seeding failures with blue grama. J. Range Manage. 24: 287-292.
Discussion Hyder, D.N., W.R. Houston, and J.B. Burwell. 1976. Drought blue grama as affected by atrazine and N fertilizer. J. Range Mnage. 29: resistance of
Drought may affect the capacity for root development from blue grama seedling crowns in two ways: (1) by reduc- tion of seedling leaf area, and (2) by direct injury of root primordia and other crown tissues. This study simulated the conditions in which drought causes die-back of exserted leaf tissue and, in addition, causes desiccation of crowns.
similar to the effects observed in this study:
In relation to soil moisture requirements for development of adventitious roots, the test conditions following tempor- ary drought were less severe than conditions generally found The combined clipping and mild drought treatments probably injured seedlings more than a similar degree of drought (without clipping) in the field. For example, one
in the field. Favorable moisture conditions during the lo- would expect that only portions of leaves would die back at a water potential of -30 to -60 bars (Wilson and Sarles
day growth test were adequate for development of leaves 1978). The retention of leaves by seedlings at such water potentials would probably result in a greater development of
and roots from many of the crowns, in spite of drought roots than in the present experiment, in which all exserted leaf tissue was removed. However, under severe drought conditions in the field, all exserted portions of leaves would
injury. On the Central Plains, periods of 2 or more days with die back; the effects of drought in that respect, would be
214-216.
Lang, A.R.G. 1967. Osmotic coefficients and water potentials of sodium chloride solutions from 0 to 4O’C. Aust. J. Chem. 20: 2017-2023. Levitt, J. 1964. Drought p. 57-66. In: M. Stelley and H. Hamilton (ed.).
Forage plant physiology and soil-range relationships. Amer. Sot. Agron., Madison, Wisconsin.
Olmsted, C.E. 1941. Growth and development in range grasses. I. Early development of Bouteloua curtipendula in relation to water supply. Bot. Gaz. 102: 499-519.
Olmsted, C.E. 1942. Growth and development in range grasses. II. Early development of Bouteloua curtipendula as affected by drought periods. Bot. Gaz. 103: 531-542.
Riegel, A. 1941. Life history and habits of blue grama. Trans. Kans. Acad of Sci. 48: 119-127.
Robinson, R.A., and R.H. Stokes. 1949. Tables of osmotic and activity coefficients of electrolytes in aqueous solution at 25OC. Trans. Faraday Sot. 45: 612-624.
van der Sluijs, D.H., and D.N. Hyder. 1974. Growth and longevity of blue grama seedlings restricted to seminal roots. J. Range Manage. 27: 117-
119.
pressure bomb. Science 155: 1248-I 254.
Wilson, A.M. 1971. Amylase synthesis and stability in crested wheatgrass seeds at low water potentials. Plant Physiol. 48: 541-546.
Wilson, A.M., D.N. Hyder, and D.D. Briske. 1976. Drought resistance characteristics of blue grama seedlings. Agron. J. 68: 479-484. Wilson, A.M., and J.A. Sarles. 1978. Quantification of growth drought
tolerance and avoidance of blue grama seedlings. Agron. J. 70: 231-237. Wilson, A.M., and D.D. Briske. 1979. Seminal and adventitious root growth of blue grama seedlings on the Central Plains. J. Range Manage. 32: 209-213.
Waring, R.H., and B.D. Cleary. 1967. Plant water stress: evaluation by
a continuously moist soil surface are rare (Briske and Wil- son 1977), and seedlings often fail to develop adventitious
White-Tailed
Deer Densities and Brush
Cover on the Rio Grande Plain
ALLEN A. STEUTER AND HENRY A. WRIGHT
Abstract
Rio Grande Plain habitats with a range in total brush cover from 10 to 97% were selected from three brush control treatments and native brush types. Deer density in each habitat was determined from helicopter census and observation towers. Three brush cover classes resulted in three levels of white-tailed deer use during summer. Areas with less than 43% total brush cover had a maxi- mum density of 1.4 deer/405 ha. Brush cover from 43 to 60% had a maximum density of 3.25 deer/405 ha. Righest summer deer use occurred on areas with 60 to 97% total brush cover (7.5 deer/40.5 ha).
The association of white-tailed deer (Odocoileus virginia- nus) with brush habitats is a subject of considerable interest
to Texas range managers. The Rio Grande Plain has exten- sive areas of dense mixed brush vegetation (Davis and Spicer 1965) and some of the highest deer densities in the United States.
Mechanical, chemical, and prescribed burning methods have been used to reduce brush cover, thereby increasing forage production and livestock handling efficiency. Often this is accomplished at the expense of deer habitat (Box 1964). At the same time, ranchers have discovered the income potential of selling hunting rights on their land (Teer and Forrest 1968). Only recently has the attempt been made to evaluate the economic returns from livestock and white- tailed deer which result from brush control treatments (Whitson et al. 1977).
Herbicidal brush control applied in strips that treat 80% of a pasture have little effect on deer numbers; however, 100% block spraying, which removes 86% of the woody canopy, has been shown to reduce deer numbers for as long as a year (Beasom and Scifres 1977). Herbicide treatments, that removed 75a/c of the living brush canopy, had no effect on “screening cover” at deer height (Tanner et al. 1978).
Brush functions as an important source of both food and cover for white-tailed deer in South Texas (Davis 1952, McMahan 1964, McMahan and Inglis 1974). Since brush provides part or all of two major components of deer habi- tat, the intensity of deer use on an area may be related to brush cover or distribution. The purpose of this study was to determine the relationship between deer density and various levels of brush cover in native brush habitats and brush control areas, which had not been commercially hunted since 1948.
Study Areas and Methods
The study was conducted on the Piloncillo Ranch, 6.4 km south of Catarina, Texas (Fig. 1). The ranch lies in the west-central
The authors are research assistant and Horn professor, Department of Range and Wildlife Management, Texas Tech University, Lubbock 79409. This research was supported in part by U.S.D.A. Forest Service, Rocky Mountain Forest and Range Experiment Station, Southern Great Plains Wildlife Habitat Laboratory, and is published as Texas Tech University, College of Agricultural Sciences Publication No. T-9-216. We thank Dan Harrison, Jr., who let us conduct this study on his ranch.
Manuscript received May 14, 1979.
328
portion of the Rio Grande Plain vegetation zone as described by Correll and Johnston (1970). Approximately 40 species of woody plants are present in the area which Muller (1947) termed “thorn scrub.” Mesquite (Prosopis glandulosa var. glandulosa) is well represented on all sites. However, blackbrush (Acacia rigidula), whitebrush (Aloysia lycioides), twisted acacia (Acacia tortuosa), lotebush (Ziziphus obtusfolia), guayacan (Porlieria angustifolia), squawbush (Condalia spathulata), Texas colubrina (Colubrina
texensis), guajillo (Acacia berlandieri), and pricklypear (Opuntia
lindheimeri) can also assume a dominant or co-dominant position, depending on local conditions.
Primary soil associations are Brundage, Maverick, Pryor (Ari- disols), and Duval and Brystal (Alfisols). Study areas included a mixture of Hardland, Rolling Hardland, and Gray Sandy Loam (CN, CS, HN, HS), mixtures of Hardland, Clay Loam, and Sandy Loam (SN, SS), Sandy Loam (PN, PS), and Clay Loam (BN, BS, LN, LS) range sites (unpublished, Dimmit Co. soil survey). Soil textures varied from sandy loam to clay.
Fig. 1. Geographical location of Piloncillo Ranch in Texas (insert) and distribution of study areas on the western harfof the Pilocillo Ranch: PN, PS = controls; SN, SS = sprayed treatments; CN, CS = controls; HN, HS = rootplowed treatments: LN, LS= controls; BN, BS= sprayedand burned treatments.
The study was conducted from May 1977 through July 1978. Research was conducted on 12 study areas of 90 to 365 ha (Fig. 1). Study areas were selected to encompass wide variation in brush cover between areas and minimal variation within study areas. Total shrub cover ranged from 10 to 97% and grass cover varied from 4 to 110%. Bare ground ranged from 0 to 37%. Sampling within study areas was confined to permanent 20.3-ha circular study sites. Thirty study sites were used.
Study areas SN and SS (Fig. 1) were aerial sprayed with Tordon 225 (1: 1, 2,4,5-T and Picloram) in June 1975 at 0.56 kg/ ha. Study areas BN and BS were sprayed with Tordon 225 in June 1975 and burned in February 1976. Study areas HN and HS were root- plowed in 1956. The remaining six study areas (PN, PS, CN, CS, LN, LS) received no brush control treatment.
Helicopter censuses by study area were conducted from 7:00 to 9:00 a.m. in January and July 1978 to determine deer density. Also, continuous 2-hr counts were made from the center of study sites using portable hunting towers 5.2 m tall. Four tower counts were logged at each study site during June, July, and August and two counts were logged on 18 selected sites during November 1977 and January 1978. Tower counts were conducted either from sunrise plus 2 hr, or sunset minus 2 hr. The timing of helicopter and tower counts corresponded to the daily peaks in deer activity (Michael
1970, Montgomery 1963).
Variability in brush cover around towers resulted in a wide range of distances at which deer could be seen. Data were analyzed on deer-per-hectare-visible basis to account for this variation in vis- ible distance. The visible area around each tower site was mapped on grid paper by two observers before deer counts were made. The number of visible hectares was determined with a planimeter. Area estimates of the two observers were usually within 10% of the mean. The visible area was defined as that area in which an active unalarmed deer could be seen at some time while moving through the site.
Brush and grass cover was recorded by species using three 30.5-m line intercepts per study site. The dead standing brush in the sprayed and burned areas added to the screening cover. Therefore, both living and dead brush were used in relating cover to deer use. Cover values for the study areas were obtained by taking the average of the study sites within an area. The horizontal distribu- tion of screening cover on study sites was described using a vegeta-
9.0 --
z 8.0 .-
a 0
3 7.0 -- Y = 0.921,*.303x
0” 3 6.0 .-
r*= 0195
L
ZZ 5.0 --
2 4.0 .-
1 I I 1 I 1 I I I
0 81 .2 .3 .4 .5 .6 .7 .8 .9 1.0
Deer Observed I.405 ha (Tower Data - 2 hr Obsn. )
Fig. 2. Relationship between deer density determined from aerial census and deer activity determined from tower observations for eight study areas. Tower data are the average of two study sites per study area.
JOURNAL OF RANGE MANAGEMENT 33(5), September 1980
tion profile board (Nudds 1977). This data provided a measure of cover stratification due to differences in canopy height and density. Herbaceous composition was determined from a minimum of 33, 0.25-m* frequency plots per study site.
Results and Discussion
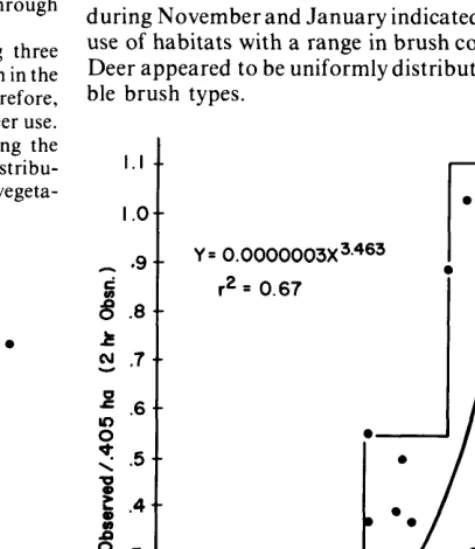
Cover of shrubs was a good predictor of habitat prefer- ence for white-tailed deer in summer. As brush cover decreased, white-tailed deer numbers declined (Fig. 3 and 4). No apparent relationship existed between shrub cover and deer use during November and January. Composition of shrubs, grasses, and forbs was not correlated to habitat preference of white-tailed deer, although when shrub cover was below SO%, a large variety of shrub species seemed more desirable than less diverse brush stands.
White-tailed Deer Census
Most data collected on white-tailed deer were obtained from systematic observations from towers on 30 study sites. To evaluate whether these data were a measure of deer density or deer activity, data taken from towers were com- pared to aerial census data for the eight study areas sampled during the summer 1978. The average density of deer on the study sites, within a study area, and the density obtained by aerial census were highly correlated (P<O.Ol) (Fig. 2). This correlation indicated that tower data could be used validly to estimate deer density on study sites, assuming that heli- copter censuses were unbiased and accurate.
Deer Density and Brush Cover
Both aerial censuses and tower counts of six study areas during November and January indicated no differential deer use of habitats with a range in brush cover from 15 to 75%. Deer appeared to be uniformly distributed across the availa- ble brush types.
r2 = 0.67
IO 20 30 40 50 60 70 80 Qb 100
Pucent Cover
of
T&al Brush SpeciesFig. 3. Relationship between the number of deer observedper 0.405 hafrom towers and percent cover of total brush species for 30 study sites during the summer. Deer data are the average offour, 2-hr observations. Circled sites not included in regression.
Tower observations suggested that deer spent a large portion of the early morning and evening hours involved in rut activities. Bucks associated with does were a common sight during this season, whereas bucks and does were segre- gated during the summer and spent most of their active time feeding. Michael (1965) reported that bucks and, to a lesser extent does, tended to have larger home ranges during the winter. The preoccupation with mating and increased mobility of deer made it difficult to determine their response to habitats of varying brush cover.
Summer deer activity on 25 study sites was positively correlated (P<O.Ol) with total brush cover in the range of 10 to 97% (Fig. 3). Similarly, aerial census of summer densities for the 12 study areas showed a positive correlation (P<O.Ol) with brush cover (Fig. 4).
Total brush cover of 43% and 60% seemed to be critical levels for white-tailed deer summer habitat (Fig. 3). Below 43% cover, deer use was low, with a maximum density of 1.4 deer/ 40.5 ha. Total brush cover from 43 to 6070 resulted in a maximum of 3.25 deer/40.5 ha. Total brush cover from 43 to 60% resulted in a maximum of 3.25 deer/40.5 ha. The highest deer use, during the summer, occurred on sites with 60 to 97% total brush cover. On these sites the maximum density exceeded 7.5 deer/40.5 ha. The maximum densities were projected using the model in Figure 2. Variability below the maximum line was probably due to site features other than cover which made some sites less suitable for deer.
Some inferences might be drawn from five sites, appar- ently unsuitable for white-tailed deer and excluded from the regression model (Fig. 3). Sites B6 and C5 had total brush cover of 60 and 63%, respectively; however, the horizontal vegetation density on these sites was relatively low. The vegetation profile indicated that height of most cover was above 1.5 m on C5 and below 1 .O m on B6. Study sites S2 and S5 were dense whitebrush bottoms. Site Ll was a dense mesquite type (86% cover) with less than 5% herbaceous cover in the understory. All five of these sites were substan- tially under used by deer relative to other sites in the same cover class (Fig. 3).
Good summer deer habitat had total brush cover about
8.0 0
6.0 --
2 5.0 --
2 z 4.0 --
j 3.0 .-
r* = 0.66
10 20 30 40 50 60 70 80 90 100
Percent Cover of Total Brush Species
Fig. 4. Relationship between deer density determined from aerial census and percent cover of total brush species for 12 study areas during the summer.
60%. Mixed brush types which resulted in understory shrub growth also characterized these sites.
The minimum grass cover for sites receiving highest deer use was 10%. Forb composition and frequency varied greatly between sites and years; however, no consistent rela- tionship could be determined with deer use of an area. Although forbs are a preferred food item, browse is the staple in deer diets on the Rio Grande Plain (Davis and Winkler 1968). Therefore, sites with a wide selection of browse species may provide a more nutritious food supply. NO relationship was apparent between brush diversity and deer use on sites with more than 5 1% total brush cover. Sites with less than 50% brush cover tended to receive heavier deer use if brush composition was more diverse.
On untreated study sites the brush cover and composition were broadly determined by range site. Range sites with lowest deer use were hardland and rolling hardland; how- ever, two of these range sites, characterized by a diverse brush composition and grass cover greater than 55%, had high deer use. Deer use of clay loam range sites varied from low to high, whereas sandy loam range sites had moderate to high deer use. McMahan and Inglis (1974) also noted that deer were attracted to sandy loam range sites.
Neither brush composition nor range site differences ade- quately explained the variability in deer use of sites with approximately equal brush cover. Deer use of sites below the maximum levels observed for the three broad cover classes (O-43%, 43-60%, 60-97%), was apparently due to habitat factors not measured during this study.
The study areas were relatively large blocks of low, medium, and high brush cover. Deer use on smaller areas of low brush cover may be substantially different than reported here. Davis and Winkler (1968) suggested that an intersper- sion of small openings in dense brush habitats may be advisable for deer.
Conclusions
The results of this study can be used to define more accurately the trade-offs between white-tailed deer and livestock production in the Rio Grande Plain. Maintenance of a given deer density requires that adquate habitat be available on a year-round basis. Total brush cover accounted for 67ajo of the variability in deer density as measured by two census methods during the summer months. At this time of year brush appears to be an impor- tant source of both cover and food. Brush control practices are obviously directed at altering this habitat component and the ranch manager should be aware that decreases in brush cover over large areas will result in lower deer use.
Literature Cited
Beasom, S.L., and C.J. Scifres. 1977. Population reaction of selected game species to aerial herbicide applications in South Texas. J. Range Man- age. 30: 138-142.
Box, T.W. 1964. Changes in wildlife habitat composition following brush control practices. in South Texas. Trans. N. Amer. Wildl. Conf. 29: 432-438.
Correl, D.S., and M.C. Johnston. 1970. Manual of vascular plants of Texas. Texas Research Foundation. Renner, Texas. 1881 p.
Davis, R.B. 1952. The use of rumen contents data in a study of deer-cattle competition and “animal equivalence.“Trans. N. Amer. Wildl. Conf. 17: 448-458.
Davis, R.B., and R.L. Spicer. 1965. Status of the practice of brush control in the Rio Grande Plain. Texas Parks and Wildlife Bull. No. 46. 40 p.
Davis, R.B., and C.K. Winkler. 1968. Brush vs cleared range as deer habitat in South Texas. J. Wildl. Manage. 32: 321-329.
McMahan, C.A. 1964. Comparative food habits of deer and three classes of livestock. J. Wildl. Manage. 28: 798-808.
McMahan, C.A., and J.M. Inglis. 1974. Use of Rio Grande Plain brush types by white-tailed deer. J. Range Manage. 27: 369-374.
Michael, E.D. 1965. Movements of white-tailed deer on the Welder Wild- life Refuge. J. Wild\. Manage. 29: 44-52.
Michael, E.D. 1970. Activity patterns of white-tailed deer in South Texas. Texas J. Sci. 21: 417-428.
Montgomery, G.G. 1963. Nocturnal movements and activity rhythms of white-tailed deer. J. Wildl. Manage. 27: 422-427.
Muller, C.H. 1947. Vegetation and climate of Coahuila, Mexico. Madrono 9 :33-57.
Nudds, T.D. 1977. Quantifying the vegetative structure of wildlife cover. Wild. Sot. Bull. 5: 113-117.
Tanner, G.W., J.M. Inglis, and L.H. Blankenship. 1978. Acute impact of herbicide strip treatment on mixed brush white-tailed deer habitat on the Northern Rio Grande Plain. J. Range Manage. 31: 386-391.
Teer, J.G., and N.K. Forrest. 1968. Bionomic and ethical implication of commercial game harvest programs. Trans. N. Am. Wildl. Conf. 33:
192-204.
The Effects of Subsurface Irrigation on
Current and Subsequent Year’s Growth in
Shadscale
PATRICIA S. JOHNSON AND BRIEN E. NORTON
Abstract
Sursurface irrigation of individual A triplex confertifofiu (shad- scale) plants was implemented in the field during the summer of 1976 through the use of vertical access tubes to a depth of 50 cm. Shoots were marked on control and watered plants and examined periodically by enumerating every leaf, bud, flower, fruit, and second-order stem. Plant response to subsurface irrigation as determined in the fall enumeration revealed a modest increase in stem length and leaf weight and summer production of lateral branches. The carryover effect of summer irrigation was reflected in new growth on shoots of watered plants in spring 1977 being more than twice the production of shoots on controls. The 1976 response to subsurface irrigation is thought to be carbohydrate storage and/or root development. Watering did not enhance bud or shoot survival overwinter.
The inhibition of cambial activity is commonly
attributed
to the action of hormones (Rubenstein and Nagao 1976, McKell and Willard 1974). Other factors that have been shown to inhibit cambial activity include water stress (Rubenstein and Nagao 1976, McIntyre 1976, Avila et al. 1975, Waisel 1970, Wardlaw 1969, Fahn et al. 1968), light intensity, and nutrition (Rubenstein and Nagao 1976). Water and available nutrients are usually limiting for extended periods in regions of the arid western part of North America. Among these constraints, water is generally regarded as the most limiting resource.Effective utilization of random summer rainfall events is dependent on timing of responses so as to optimize growth and survival under a generally arid regime (Noy-Meir 1973). Stem elongation in shadscale (Atriplex confertzfolia (Tor. & Frem.) S. Wats.), a halophytic shrub of the Great Basin desert, is largely confined to spring months under natural
Authors are research technician and associate professor, Department of Range Science, Utah State University, Logan 84322.
This research was supported through the US/IBP Desert Biome Program; addi- tional funding was also provided by the Utah State University.
Manuscript received March 3, 1979.
JOURNAL OF RANGE MANAGEMENT 33(5), September 1980
conditions. At this time soil water potential is relatively high due to moisture recharge of the soil profile from snowmelt (Caldwell et al. 1977). Summer precipitation in semiarid areas results in an increased soil water potential only in the surface layers, which dry quickly if further rain is not imme- diately forthcoming. Summer storms do not stimulate addi- tional shoot growth of shadscale, even when these storms are augmented by surface irrigation (J.A. MacMahon unpublished data, Hodgkinson et al. 1978). Root growth, however, continues through the summer deep in the soil profile (Fernandez and Caldwell 1975, Holthausen 1977). The inherent quiescence of the shoot system in summer can be interpreted as an adaptive mechanism to the aridity of the season, thereby avoiding the cost of responding to erratic and short-lived soil water impulses. If, however, water stress is responsible for the inhibition of shoot growth and bud elongation, relief of that stress should induce growth. Since shadscale is better able to use water from deep in the soil profile than from near the surface during the summer (Johnson and Norton 1979), surface watering is probably not effective as an ameliorating treatment.
In this study an attempt was made to experimentally disturb the quiescent status of the shrub by subsurface irri- gation and to record subsequent response under field condi- tions. We hypothesize that reducing summer water stress of shadscale will (1) stimulate above-ground growth during the season of treatment, (2) favor the retention and develop- ment of vegetative and reproductive parts, and (3) increase production the following spring.
Site and Methods
The study was conducted at the U.S. International Biological Program (US/IBP) Desert Biome intensive study site in Curlew Valley, near Snowville, Utah. The area has been described in detail by Balph (1973). The study site is co-dominated by shadscale and big sagebrush (Artemisia tridentata Nutt.); squirreltail (Sitanion
hystrix
(N
utt.) J.B. Smith) is the major grass species associated with these shrubs. Crucifers and chenopods contribute the bulk of a sparse annual plant component. Mean elevation at the site is 1,320 m and soils are primarily silty loams. Mean annua precipita- tion is 230 mm, 60 to 70% of which comes during winter.Shoot Enumeration Technique
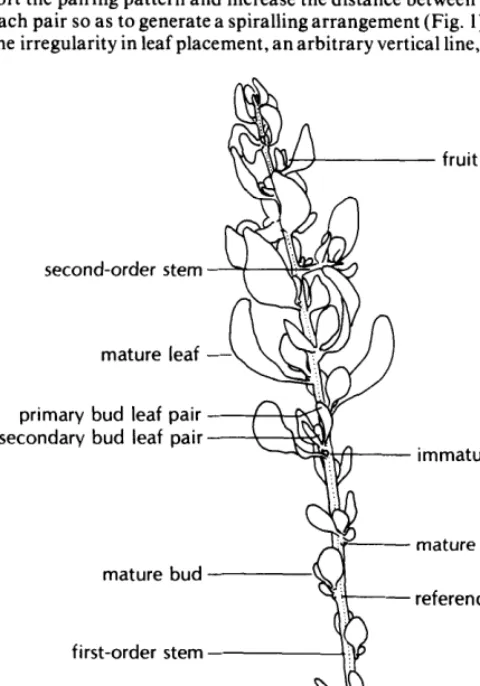
A method similar to that used by West and Fareed (1973) was developed to describe individual marked stems by identifying and recording all buds, leaves, flowers, fruits, and second-order stems and their relative locations on the shoot. The following terms describing shoot parts will be used throughout this paper-as they are defined below and illustrated in Figure 1.
Immature buds-buds with leaves up to 1 mm long. Mature buds-buds with leaves more than 1 mm long but with no internodal elongation.
Mature leaves-leaves which subtend an axillary bud and are found on elongated stems.
Bud leaf pairs--pairs of leaves of similar size, apparently opposite each other in the mature bud but upon stem elongation separated by an internode. Since several bud leaf pairs may occur in one mature bud, the individual pairs were designated as primary, secondary, etc., according to their vertical position in the bud.
First-order stem-the main stem of a shoot, originating from a bud on the previous year’s stem.
Second-order stem-a stem originating from buds on the first-order stem of the current year’s shoot growth.
The primoidial leaves appear to be opposite; during shoot elon- gation, however, differential growth and twisting of the stem dis- tort the pairing pattern and increase the distance between leaves of each pair so astb generate a spiralling arrangement (Fig. 1). Due to the irregularity in leaf placement, an arbitrary vertical line, running
second-order stem
mature leaf
primary bud leaf pair secondary bud leaf pair
bud
mature bud
mature leaf scar
- reference line
first-order stem -
two buds at one node-
basal node
Fig. 1. Diagram of a current year’s shadscale shoot.
from the basal node to the tip of the stem, was used as a reference line in the enumeration process. The nodes were grouped into sets, each set beginning with a node closest to the reference line and including all successive nodes (moving toward the top) between it and the next node near the line. The enumeration began at the base of the stem and ended at the apex, and each node was enumerated in order within its set. The enumeration of second-order stems was separate from that of the first-order stem, and was done in the same fashion.
Watering Treatment
During the spring of 1976 a field population of shadscale plants was located, all of approximately the same size within a l-hectare portion of the study site. Pairs of plants of similar size, in the same shoot length class (short, medium, and long) and of identical sex were identified in the field; one member of the pair was randomly assigned to a watering treatment and the other was designated as control, until there were ten plants in a group. Male and female plants of each shoot length class were represented in the two groups. This procedure of plant selection which provided matching individuals in the control and treated experimental populations, was necessary in order to analyze the data statistically using pair- wise methods.
Three white polyurethan pipes (diameter 2.5 cm) were installed around each of the watered plants but not the controls. The pipes were buried vertically to depths of 15, 30, and 50 cm and were located 10 cm to the north, west, and south of the plant, respec- tively. Gravel was poured into the pipes to a depth of about 2 cm to reduce the erosive impact of the watering; the pipes were capped to prevent direct evaporation. Irrigation was confined to these pipes and usually occurred once a week from May 26 to September 13; up to 500 ml of water were poured into each pipe on each water day. The total amendment averaged 21.8 liters per plant. Assuming a soil water utilization zone of 25 cm radius from the taproot, if the irrigation water had fallen as rainfall it would be equivalent to approximately I10 mm, which is more than the average precipita- tion over the same summer period.
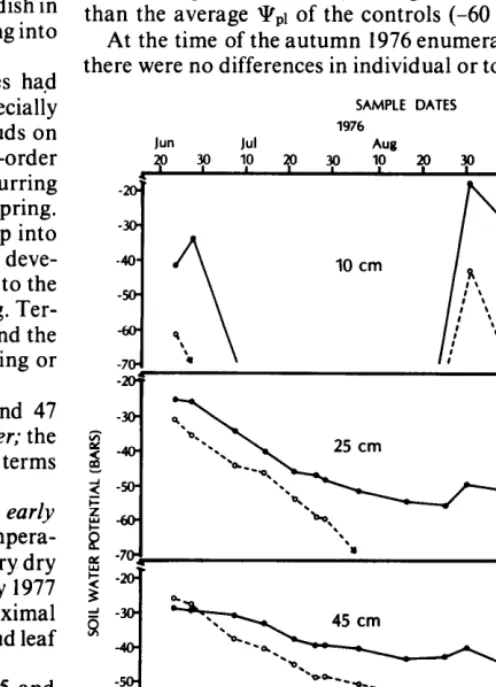
Ceramic-cup thermocouple psychrometers (manufactured by Wescor, Inc.) were buried in a vertical line 10 cm from the east side of two plants per treatment at depths of 10, 25, 45, and 60 cm. These were read at dawn with a Wescor MJ 55 microvoltmeter periodically through the summer, immeditely before irrigation. The psychrometers were also read twice in 1977, on April 29 and June 28. Prior to installation, each psychrometer was calibrated with NaCl solutions (Brown 1970).
Pressure bomb readings of plant water stress for both control and watered plants were taken at dawn approximately every two weeks (Scholander et al. 1965, Waring and Cleary 1967). These measurements occurred approximately 1 week after the last appli- cation of water and immediately before the next irrigation, Read- ings were taken on three to four stems for both control and watered treatments at each sampling date.
Four stems on each control and watered plant were chosen and marked with lengths of colored thread. A complete enumeration of the marked stems occurred four times: at the end of the spring growth period in June 1976; in October 1976; prior to spring growth in April 1977; the fourth occasion was the end of the spring growth period 1977, but did not include an enumeration of the new 1977 stems. One unmarked stem (representing less than 2% of the current year’s shoot growth) was harvested from each of the 10 control plants for leaf, fruit, and stem analysis at the time of the first enumeration in 1976. Stem lengths were measured and leaves, and fruits, and stems were dried for two days at 800 C, and weighed. Half the marked stems were harvested from every plant at the end of the second enumeration in October 1976. Leaves and fruits were separated from the stems, leaves were sorted according to their leaf classification, and all stems were measured for length. All leaves, fruits, and stems were dried at 800 C for two days and weighed. The remaining marked stems were harvested after the final enumera- tion in June 1977. Leaves and flowers were separated from the