Copyright 0 1994 by the Genetics Society of America
Genetic Analysis of 63 Mutations Affecting Maize Kernel Development
Isolated From
Mutator
Stocks
Michael J. Scanlon,*9' Philip
S.Stinard,+ Martha G. James,* Alan M. Myers*92 and
Donald S. Robertson*
Departments of *Biochemistry and Biophysics, tAgronomy, and zoology and Genetics, Iowa State University, Ames, Iowa 5001 1 Manuscript received May 7 , 1993
Accepted for publication September 28, 1993
ABSTRACT
Sixty-three mutations affecting development of the maize kernel were isolated from active Robert- son's Mutator ( M u ) stocks. At least 14 previously undescribed maize gene loci were defined by mutations in this collection. Genetic mapping located 53 of these defective kernel (dek) mutations to particular chromosome arms, and more precise map determinations were made for 21 of the mutations. Genetic analyses identified 20 instances of allelism between one of the novel mutations and a previously described dek mutation, or between new dek mutations identified in this study; phenotypic variability was observed in three of the allelic series. Viability testing of homozygous mutant kernels identified numerous dek mutations with various pleiotropic effects on seedling and plant development. The mutations described here presumably arose by insertion of a Mu transposon within a dek gene; thus, many of the affected loci are expected to be accessible to molecular cloning via transposon-tagging.
T
he maize kernel provides a particularly tractable genetic system for analysis of tissue develop- ment and differentiation in higher plants (NEUFFER etal. 1986; SHERIDAN 1988). Morphological develop- ment of the triploid endosperm tissue and the con- spicuous embryo in maize kernels can be monitored easily, and several hundred kernels are available for study on a single ear. Extensive efforts have utilized mutations that affect anthocyanin accumulation in the aleurone, the single-celled outer layer of the endo- sperm, to analyze tissue-specific gene expression, tran- sposon structure and function, and other biological activities. Another large body of work has centered on the numerous genetic loci that affect composition of starch or protein in the endosperm. Kernel muta- tions also have been useful in characterizing develop- ment of the maize embryo (CLARK and SHERIDAN
1986, 1988, 1991; MCCARTY et al. 1989a, 1991;
ROBERTSON 1952; SHERIDAN and CLARK 1987; SHER-
An abundant class of maize mutations, termed de- fective kernel (dek) mutations, were first characterized
by JONES (1920) and MANGLESDORF (1923, 1926) based on kernel, phenotypes involving profound ef- fects on both endosperm and embryo development. Typically, dek mutations are recessive and in the homozygous state frequently cause development of inviable kernels. Probably because of difficulties as- sociated with genetic analyses of recessive-lethal mu-
IDAN 1988).
'
Present address: Department of Plant Biology, University of California,'
To whom correspondence should be addressed.Berkeley, California 94720.
Genetics 1 3 6 281-294 (January, 1994)
tations, the dek genes as a class were largely neglected for the more than 50 years spanning the early studies of JONES and MANCLESDORF and the later work of
NEUFFER and SHERIDAN (1980) and SHERIDAN and
NEUFFER (1980). These latter reports described ge- netic, morphological, lethality, and embryo rescue studies of numerous dek mutations induced by the mutagen ethylmethane sulfonate (EMS). Genetic map- ping located 105 dek mutations to 17 of the 20 maize chromosome arms, and a diverse array of kernel mu- tant phenotypes was described. Subsequent analyses of EMS-induced dek mutations identified 33 unique gene loci (NEUFFER et al. 1986). Of 855 recessive mutations recovered in kernels in the homozygous state, only 147 yielded viable plants, although many produced inviable seedlings with altered pigmenta- tion. Histological examination showed dek mutant em- bryos typically were affected more severely than were the endosperms of the same kernels. Although im- mature dek mutant embryos were blocked in devel- opment over a wide range of stages, the majority in the study (80%) were able to germinate in tissue culture (SHERIDAN and NEUFFER 1980), indicating that embryo lethality often occurs late in kernel de- velopment.
Further investigations utilizing dek mutations as a
genetic investigations also will be useful, for example analyses of background effects, double mutants and phenotypic variation resulting from allelic series of
dek mutations.
AS a step toward the molecular analysis of dek gene function, this report describes the genetic character- ization of 63 kernel mutant isolates from ROBERTSON'S
Mutator
(Mu)
stocks (ROBERTSON 1978). One of the goals of this mutagenesis project was to generate dek mutations by insertion ofMu
transposons, for use in molecular cloning studies of genes required for kernel development. Viability studies of these putative Mu- induced dek mutations are presented, and chromo- some arm placements are reported for 53 mutations. More precise genetic mapping information obtained via linkage determinations is reported for 21 muta- tions. Fourteen previously undescribed dek gene loci were identified, and the pleiotropic effects of several of these dek mutations are discussed. The accumulated data presented here will be useful in future genetic, biochemical and molecular analyses of these various genes that affect development of the maize kernel.MATERIALS AND METHODS
Stocks: Mutator lines are maize stocks that exhibit Mutator activity as defined by ROBERTSON (1978). Standard lines, defined as stocks that have never been crossed to Mutator lines, are the F1 hybrids Q66/Q67 (inbred Q66 X inbred Q67), or B77/B79 (inbred B77 X inbred B79). T h e com- plete set of B-A translocations and the waxy-marked trans- location series were propagated at Iowa State University by D. S. ROBERTSON. A series of 31 EMS-induced mutations, dekl through dek3l (NEUFFER et al. 1986), used to test for allelism with the putative Mu-induced dek mutations were obtained from W. F. SHERIDAN. All stocks of marker genes used in linkage determinations were supplied by the Maize Genetics Stock Center, University of Illinois, Urbana, Ill., except the brnl-R Lg3 stock, which was developed by D. S. ROBERTSON and P. S. STINARD.
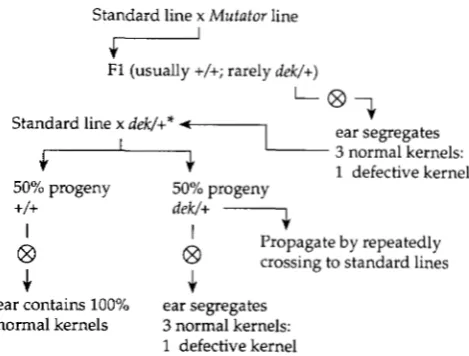
Isolation and propagation of maize kernel mutants: The strategy for the isolation of dek mutations from a Mutator stock is presented in Figure 1. Active Mutator plants were outcrossed as males onto standard lines, and F1 plants grown from the resulting kernels were self-pollinated. The self- pollinated ears were then scored for the presence of defec- tive kernels, with the expectation that onequarter of the kernels on an ear would be mutant if a recessive dek mutation was carried by the F1 plant. Most of the dek alleles in this study are recessive-lethal mutations; thus, they were propa- gated by outcrossing dek/Dek heterozygotes to DeklDek standard lines. T o do so, plants were grown from normal sibling kernels obtained from self-pollinated ears displaying onequarter defective kernels. These plants were self-polli- nated to determine their genotype, and also outcrossed as males onto standard lines. Genotype was determined by observing the self-pollinated ears; the presence of one- quarter defective kernels identified dek/Dek plants, whereas all normal kernels indicated DeklDek plants. Approximately 10 progeny kernels from the outcrosses of dek/Dek plants were grown to maturity, and these were both self-pollinated and outcrossed as males to standard lines in a continuation of this self-pollination/outcross protocol. As expected, in
Standard line x Mutator line
r"
F1 (usually +/+; rarely dek/+)
Standard line x dek/+* ?
+
7
Y ~ ~ ~ Z X Z e l s :50% progeny 50% progeny
+/+
1 defective kernel
I dek/+ I
7
€3
@ Propagate by repeatedly crossing to standard linesJ.
J.
ear contains 100% ear segregates normal kernels 3 normal kernels:
1 defective kernel
FIGURE 1.-Strategy for induction and propagation of maize kernel mutants using Robertson's Mutator. See MATERIALS AND METHODS for details. The genotype of the plant denoted with an asterisk was distinguished from its Dek/Dek siblings by observation of one-quarter defective kernels on self-pollinated ears.
most instances approximately one-half of the progeny plants of each outcross were found to be heterozygous for the dek mutation.
Viability tests and classification of dekldek seedling phenotypes: Viability tests were performed on dekldek de- fective kernels that upon inspection at maturity contained some visible embryonic structure, by planting 20 mutant kernels in the field or in pots in the greenhouse. Those dek mutations that resulted in kernels with no visible embryonic structure were classified as germless-lethal without further analysis. Mutations causing a 5% or lower germination rate were classified as lethal. Significant germination rates of dekldek mutant kernels did occur in some instances, as described below.
The germination products of defective kernels were noted for seedling and/or plant phenotypes. In rare in- stances, dekldek kernels produced plants with fertile ears and tassels; these were self-pollinated, and ears were har- vested and scored for the corresponding kernel phenotype to verify the homozygous genotype of the parental plants. Whenever putative dekldek homozygous plants produced functional ears but no pollen, they were pollinated by stand- ard Q66/Q67 plants. Kernels from the resulting crosses were planted, and the plants were self-pollinated and scored for kernel mutant phenotype to verify the homozygous genotype of the parental plants. All plants in this outcross are expected to be dek/Dek heterozygotes and produce one- quarter defective kernels upon self-pollination.
Genetic mapping: Three different techniques were used for mapping dek mutations to a specific chromosome arm. Placement was most often accomplished using the B-A trans- location method (ROMAN and ULSTRUP 1951) as described by BECKETT (1978). In other instances, mutants were placed using either the waxy-marked translocation series as de- scribed by ANDERSON (1956) and ROBERTSON (1955), or demonstration of allelism to a previously mapped dek mu- tation (described in the following section). Analyses of link- age of kernel mutants to marker genes of known map location were performed according to standard techniques.
Maize Kernel Development Mutants 283 both ears of this cross produced 100% mutant kernels.
When dek mutations to be tested were lethal in the homo- zygous state, or produced sterile plants, allelism tests were conducted using 20 plants grown from progeny kernels of each dek/Dek isolate outcrossed to standard. One-half of these kernels were expected to be heterozygous for the particular dek mutation. In some instances, 20 normal ker- nels were planted from a self-pollinated ear of a heterozy- gous mutant plant, with the expectation that two-thirds of the plants would be heterozygous for the particular dek mutation. Allelism tests were performed by outcrossing pollen of one mutant family (mutant A) to the first ear of a second mutant family (mutant B), and self-pollinating the second ear of mutant A; this procedure was carried out in reciprocal fashion. Whenever a mutant plant did not pro- duce a second ear, that plant was self-pollinated and the pollen was used in an outcross to the first ear of a second mutant family. Allelism was assigned whenever the out- crossed ears of two dek/Dek heterozygous plants from differ- ent mutant families (genotype determined by self-pollina- tion) segregated at a 3: 1 ratio for normal kernels relative to mutant kernels. Independent dek mutations were deter- mined to be nonallelic whenever the outcross ear of such a cross produced all wild-type kernels.
RESULTS
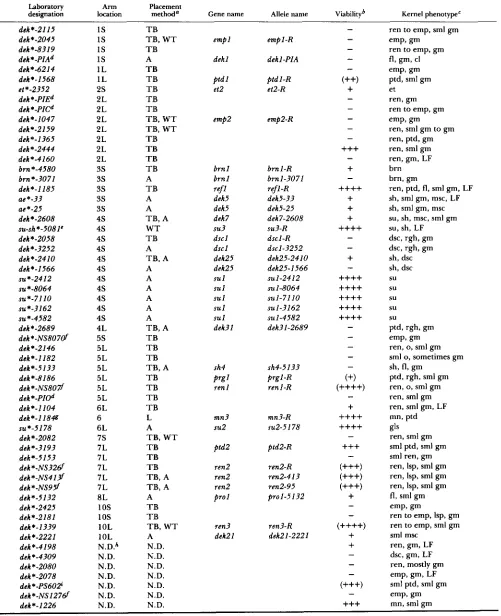
Descriptions and inheritance of the mutant phe- notypes: Sixty-three dek mutations were isolated from active Mutator lines as described in MATERIALS AND METHODS. Table 1 lists for each mutation the initial laboratory designation, the chromosome arm location, a summary description of the kernel phenotype, and an assessment of the viability of dekldek plants. Table 1 also indicates those mutations that could be assigned as alleles of previously known genes or novel genes identified in this study (see below). Of the mutations listed in Table 1, all are recessive regarding the defec- tive kernel phenotype. These mutations all were iden- tified by self-pollinating dek/Dek heterozygous plants and propagated by outcrossing such plants onto stand- ard lines as shown in Figure 1. All have been shown to be heritable.
T h e mutations listed in Table 1 display a wide range of phenotypes (described in Table
2);
the two most common types are reduced endosperm size (ren) and empty pericarp or papery kernel (emp). Single ears segregating for the ren or emp phenotype often dis- played significant variation in the degree of mutant kernel filling. Although the emp mutant kernels are almost always germless, the ren mutant kernels often contained a small embryo, and larger ren mutant kernels usually contained larger embryos than their smaller sibling mutants. Other commonly observed kernel mutant phenotypes included sugary-shrunken, miniature seeds, etched-pitted kernels and aleurone mosaics (Table 1).Most of the mutant phenotypes were inherited in the normal Mendelian ratios expected for single fac- tor-controlled traits. Eight mutations, however, showed a low frequency of expression (LF); in these
instances self-pollinated ears of heterozygous plants displayed fewer than the expected 25% defective ker- nels.
Genetic mapping of deh mutations: Classical ge- netic mapping techniques were used to locate 53 dek mutations on specific chromosome arms (Table 1). Mutants were placed to 15 of the 20 chromosome arms of maize. Subsequent genetic tests, including allelism determination and linkage analyses, were used to confirm the placement of 33 mutations. In most instances mutants were placed using the B-A translo- cation series (ROMAN and ULSTRUP 195 1 ; BECKETT 1978) or the waxy-marked translocation series (AN-
DERSON 1956; ROBERTSON 1955); some mutations that
caused phenotypes reminiscent of those resulting from previously mapped mutations were placed by positive tests of allelism. Chromosome arm locations were not determined for 11 of the mutations. Because most of the mutations are lethal in the homozygous condition and were tested as heterozygotes, and some B-A trans- location stocks shed little pollen, some dek mutations were not adequately tested by all B-A translocation stocks. Also, some of the unlocated mutant genes may be proximal to the breakpoints of the corresponding B-A translocation or too far removed from waxy marked translocation breakpoints to detect linkage (ROMAN and ULSTRUP 1951; BECKETT 1798; ANDER-
SON 1956; ROBERTSON 1955). Finally, placement of
certain kernel mutations can be difficult because some B-A translocations produce small, opaque hypoploid endosperms, a phenotype resembling that of many of the kernel mutants being tested (ROMAN and ULSTRUP
195 1).
Assignment of dek mutations as alleles of known or novel genetic loci: Allelism tests were performed as an aid in genetic mapping efforts and in order to distinguish novel kernel mutations from alleles of previously described loci. After genetic mapping to a specific chromosome arm, dek mutations were tested for allelism with previously described mutations lo- cated on the same chromosome arm that also affect kernel development. T h e results of the allelism tests are given in Table 1, which lists 20 identified cases of allelism among the 63 mutants studied. This list in- cludes nine putative Mu-induced alleles of dek loci identified by NEUFFER and SHERIDAN (1980), five alleles of the sul locus and three pairs of alleles of novel dek loci first identified in this study.
TABLE 1
Characterization of dek mutations
Laboratory A m
designation location methoda Gene name Allele name Viabilityb Kernel phenotype' Placement dek*-2115 dek*-2045 dek*-8319 dek*-PIAd dek*-6214 dek*-1568 et*-2352 dek *-PIEd dek *-PICd dek*-1047 dek*-2159 dek*-1365 dek*-2444 dek*-4160 brn*-4580 bm*-3071 dek*-1185 ae*-33 ae*-25 dek*-2608 dek*-2058 dek*-3252 dck*-2410 dck*-1566 su*-2412 su-sh*-5081C
su *-8064 ~ ~ * - 7 1 1 0 ~ ~ * - 3 1 6 2 su *-4582 dek*-2689 dek*-NS8070f dek*-2146 dek*-1182 dek*-5133 dek*-8186 dek*-NS807f dek*-PIOd dek*-1104 dek*-1184
su *-5178 dek*-2082 dek*-3193 dek*-5153 dek*-NS326f dek*-NS413f dek*-NS95f dek*-J132 dek*-2425 dek*-2181 dek*-1339 dek*-2221 dek*-4198 dek*-4309 dek*-2080 dek*-2078 deh*-PS60P dek*-NS12761 dek*-1226
1s
1s
1s
1s
1L 1L 2 s 2L 2L 2L 2L 2L 2L 2L3s
3s
3s
3s3s
4 s 4s 4s 4 s 4 s 4 s 4s 4 s 4 s 4 s 4 s 4L 5 s 5L 5L 5L 5L 5L 5L 6L 6 6L 7 s 7L 7L 7L 7L 7L 8L 1os
1os
1 OL 1 OL N.D.* N.D. N.D. N.D. N.D. N.D. T B T B , W T T B A T B T B T B T B T B T B , W T T B , W T T B T B T B T B A T B A ATB , A
W T T B
A
T B , A A A A A A A
TB, A
T B T B T B TB , A
T B T B T B T B L A
T B , W T T B T B T B TB, A
T B , A A
T B T B T B , W T
A N.D. N.D. N.D. N.D. N.D. N.D. N.D. N.D.
empl
dekl
ptd 1 et2
empa
brn 1 b m 1
dek5 dek5 dek7 st43 dscl dscl dek25 dek25 su 1 sul s u l su 1
su 1 dek3l ref1
sh4
ren 1
Prgl
m n 3
su2
p t d 2
ren2 ren2 ren 2 pro1 ren3 deli21 empl-R dekl-PIA
p t d l - R etZ-R
emp2-R
brn 1 -R
brnl-3071 refl-R dek5-33 dek5-25 dek7-2608 dscl-R dek25-2410 dek25-1566 su3-R
d ~ l - 3 2 5 2
SUI-2412 su 1-8064 SUI-7110 SUI-3162 su 1-4582 dek31-2689 sh4-5133 prg1-R renl-R mn3-R SUZ-51 7 8
ptd2-R
r e d - R ren2-4 1 3 ren2-95 prol-5132 ren3-R dek21-2221
-
-
-
-
(++)-
+
-
-
-
-+++
--
+
-
++++
+
+
+
++++
-
-
+
++++
++++
++++
++++
++++
-
-
-
--
- (+) (++++)-
+
++++
++++
-+++
-
(+++) (+++) (+++)+
-
-
(++++)+
+
--
-
(+++) -+++
ren to emp, srnl gm emp, gm
ren to emp, gm
fl, gm, cl emp, gm ptd, srnl gm et
ren, gm ren to emp, gm emp, gm
ren, srnl gm to gm ren, ptd, gm ren, srnl gm ren, gm, LF
brn brn, gm
ren, ptd, fl, srnl gm, LF
sh, srnl gm, msc, LF
sh, srnl gm, msc
su, sh, msc, srnl gm
su, sh, LF
dsc, rgh, gm dsc, rgh, gm sh, dsc sh, dsc su su su su su
ptd, rgh, gm emp, gm ren, 0, srnl gm
srnl 0, sometimes gm
sh, fl, gm ptd, rgh, srnl gm ren, 0, srnl gm
ren, srnl gm ren, srnl gm, LF
mn, ptd
ren, srnl gm srnl ptd, srnl gm srnl ren, gm ren, Isp, srnl gm ren, Isp, srnl gm ren, Isp, srnl gm fl, srnl gm emp, gm
ren to emp, Isp, gm ren to emp, srnl gm srnl msc
ren, gm, LF
dsc, gm, LF
ren, mostly gm emp, gm, LF
Maize Kernel Development Mutants
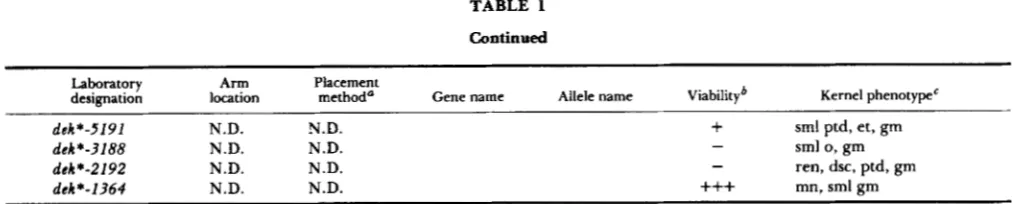
TABLE 1
Continued
285
Laboratory Arm Placement
designation location method" Gene name Allele name Viabilityb Kernel phenotype'
I
dek+-5191 N.D. N.D.
+
sml ptd, et, gmdek*-3188 N.D. N.D.
dcb*-2192 N.D. N.D.
dcC*-1364 N.D. N.D.
+++
mn, sml gm- sml 0, gm
-
ren, dsc, ptd, gm* TB, placement determined using appropriate B-A translocation; A, placement determined using allelism test; WT, placement determined using appropriate waxy-marked translocation (see Table 4 for specific translocation used); L, placement determined via linkage to a marker on the same chromosome.
Results of lethality tests are for homozygous mutants, -, inviable kernels, no germination;
+,
seedlings lethal, die before five leaf stage;++,
plants develop beyond five-leaf stage but die before producing flower inflorescence;+++,
plants produce male and/or female flower(s), but one (or both) is sterile;++++,
fertile plants, produce functional male and female flowers; ( ), less than 5% of mutant seed germinate to display indicated phenotype.See Table 2 for explantation of symbols used to describe kernel phenotypes. Mutants provided by STEVE BRIGGS, Pioneer Hi-bred International, Inc. May involve duplicate factors, with one factor located on chromosome 4 . f Mutants provided by NANCY SHEPHERD, DuPont de Nemours, Inc.
gdek*-Mu1184 is linked to y l on chromosome arm 6L, but is not uncovered by either TB-GLc, nor TB-6Sa. Therefore, it is located proximallv to the breakpoints of these B-A translocations.
*
N.D.,'not determin'ed.Mutant provided by PATRICK SCHNABLE, Iowa State University.
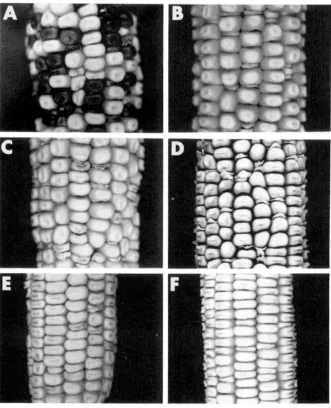
mutations in six of these novel loci are presented in Figure 2.
Phenotypic variation caused
by
allelic mutations:In most cases the kernel and seedling phenotypes (where applicable) caused by ailelic mutations were identical, although notable exceptions exist. For ex- ample, kernels homozygous for b m l - R are of normal size, and have dark brown endosperms and light brown embryos. In contrast, kernels homozygous for the allelic mutation bml-3071 are relatively small, the brown coloration in the endosperm is much less se- vere, and most kernels are germless; the rare germs that do form are lightly pigmented. Kernels homozy- gous for bml-R produce pale-green, inviable seed- lings, whereas those homozygous for bml-3071 do not germinate. Heterozygous bml-R/bml-3071 ker- nels display yet a third distinct phenotype; these ker- nels are pale brown, of normal size and produce pale- green, inviable seedlings.
Phenotypic heterogeneity also was observed for two novel alleles of dek25 (NEUFFER and SHERIDAN 1980) identified in this study. Although both the reference allele dek25-R and dek25-1566 are lethal mutations, more than 10% of homozygous dek25-2410 kernels germinated to produce striated, inviable seedlings (Table 1). Homozygous mutations in any of these three dek25 alleles produced no obvious differences in kernel phenotype. Kernels homozygous for two novel alleles of dek5 (NEUFFER and SHERIDAN 1980) identified in this study are phenotypically identical, although the mutations display dissimilar inheritance patterns. Specifically, dek5-33 is inherited in extremely low frequency as compared with both dek5-25 (Table 1) and the EMS-induced reference allele dek5-R
(NEUFFER and SHERIDAN 1980). The last incidence of
phenotypic heterogeneity observed was caused by dif- ferent mutations of the SUI locus. Kernels homozy- gous for the reference allele sul-R displayed much more kernel wrinkling and translucence than did het- erozygous kernels containing one copy of sul-R and one copy of the allelic mutation sul-2412.
Effects of specific deR mutations on plant pheno- type: Kernels homozygous for each dek mutation were tested for the ability to germinate (Table 1). Results of these lethality screens are reported on a scale from
"-," for lethal kernels exhibiting less than 5 % germi- nation, up to
"++++"
for mutant kernels that yield fertile plants, as detailed in the legend of Table 2. Of the 63 putative Mu-induced kernel mutations tested, 39 were embryo lethal in the homozygous condition, and 11 others caused seedling lethality in which plants died before reaching the five leaf stage of vegetative development. Four kernel mutations resulted in plants that reached the reproductive stage of development, but the male or female flower, or both, were non- functional. Nine other mutations were classified as viable and produced fertile plants from which self- pollinated ears were obtained.M. J. Scanlon
TABLE 2
Kernel phenotypes
Symbol Phenotype
TABLE 3
Novel genes affecting kernel development
Gene name Phenotypic attributes Brown endosperm-brown color in endosperm cells
Colorless aleurone-absence of anthocyanin pig-
Defective kernel
Discolored-some variation of normal clear yellow and the embryo
ments from aleurone cells
or white color, as though stained reddish or brownish
Empty pericarp-kernels with little or no endo- sperm tissue, flattened and papery
Etched-surface of kernel with irregular network of pits and fissures in underlying endosperm Floury-endosperm has a soft, chalk-like texture, a
generally reduced yellow color and an opaque a p pearance
Glassy-endosperm translucent with a hard flinty appearance, like su2
Germless-embryo very poorly developed or en- tirely absent
Low frequency of mutant kernels segregating on an ear, and/or low frequency of dek/Dek ears in a
family; non-Mendelian ratios of wild-type to mu- tant phenotype
from the aleurone, producing a grayish appear- ance over part of the kernel
Miniature-kernel typical in shape, form and tex- ture, but slpaller in size and with slightly loose pericarp
Mosaic-irregular-shaped patches of color and non- color in the aleurone tissue when appropriate genes for anthocyanin are present; the borders are distinct, as opposed to the mottled effect
chalky and waxy
indentations
size, often associated with loose pericarp and small embryo
Rough-surface of kernel covered with many small irregularities, giving it a rough texture
Shrunken-two types: (1) a moderately smooth col- lapse of the crown and/or both sides of the ker- nel, like s h l , and (2) an extreme collapse of endo- sperm tissue, often associated with a brittle and/ or translucent appearance, like sh2
Small-used in conjunction with other descriptive terms to designate smaller size
Sugary-kernel is shriveled, hard, and translucent; typical of sul
Loose pericarp-a space separates the pericarp
Opaque-endosperm is opaque and firm, not
Pitted-surface of kernel strewn with small pits or
Reduced endosperm-endosperm much reduced in
Adapted from SHERIDAN and NEUFFER (1980).
~
germination rate and produced small, infertile plants with narrow leaves and frayed, necrotic leaf margins. Other pleiotropic effects of this mutation include ru-
dimentary ear and tassel formation. Although more than 50% of the small seeded mutant kernels homo- zygous for dek*-2444 germinated, small narrow-leafed plants were produced that formed sterile ears and tassels. Numerous aberrations of shoot development
ptd I
p t d 2
ren 1
ren2
ren3
emp 1
emp2
dscl
et2
b m 1
mn3
su3
P%1
ref 1
Small kernels with pitted, scarred endosperm and small germ; usually lethal, although kernels with larger embryos will germinate to produce, small, nonflowering plants with large, necrotic, mottled sectors on leaves.
Pitted, cracked endosperm and small germ; a p proximately 10% of kernels will germinate to produce plants with narrow, frayed leaves with necrotic margins; forms sterile, rudimentary ear with very short silks and sterile unbranched tassel with small branch nodes.
Small kernels with reduced, opaque endosperm and small germ; usually inviable but kernels with larger embryos will germinate to produce fertile plants from which self-pollinated, homo- zygous ears have been obtained.
Endosperm variably reduced in size, often with loose pericarp and small germ; lethal, although larger kernels may produce small plants with rudimentary, sterile tassel.
Reduced endosperm, varying from partially filled to empty pericarp; may have small germ, or
germless; often lethal, but larger kernels via- ble, will produce fertile plants; homozygous ear has been obtained.
material and flattened by compression of sur-
rounding normal seed; germless, lethal. Papery, empty pericarp with very little or no en-
dosperm material; germless at maturity, lethal. Discolored, opaque or brownish seed, with dis-
torted shape and an irregular, rough texture; germless at maturity, lethal.
Etched endosperm; small germ, yields mutable, inviable albino seedlings with occasional green- ing of leaf tips.
( b m l - R ) Brown endosperm and embryo; kernels produce pale-green, inviable seedlings. ( b m l - 3 0 7 1 ) Light brown endosperm; usually germ- less, germs that do form are lightly pigmented; lethal.
Minature seed; small kernel, etchedlpitted endo- sperm; viable.
Sugary endosperm; endosperm glassy, smoother than s u l , and shrunken; viable. May involve duplicate factors.
Small, pitted, rough endosperm; usually germ- less, although kernels with larger embryos will germinate to produce small, striated seedlings that die at the two- to three-leaf stage. Reduced, pitted endosperm with dull, floury a p
pearance; small germs often present; very low frequency of expression on segregating ears; approximately 20% will germinate and pro- duce plants with well-developed ears, and tas- sels that shed little pollen.
Papery, empty pericarp, devoid of endosperm
also were exhibited by mutant plants homozygous for
Maize Kernel Development Mutants 287
J
r*
. "
-
ri. 1
FIGURE 2.-Segregation of novel deh mu- tations. Ears segregating mutant and wild- type kernels were obtained by self-pollination of plants heterozygous for the indicated deh mutation. (A) p i d l - R . (B) renl-R. ( C ) dscl-R.
(D) cmp2-R. (E) prgl-R. (F) rcfl-R.
brachytic (short internode) and produce almost twice as many leaves per linear foot of plant height as do
dek*-1364 heterozygote sibling kernels. The ligule and auricle domains of dek*-1364 homozygous plants are disrupted, and ligular tissue is ectopically ex- pressed in the leaf midrib. One result of the ligular- auricular disruption is leaves that droop severely. Lastly, the tassels of dek*-1364 homozygous plants were barren, although 5 of the 10 plants obtained produced ears that set seed when pollinated by stand- ard Q66/Q67. Thus, dek*-1364 does not block nor- mal development of the female gametophyte.
Several mutations in this study were classified as viable, with germination rates of 90% or greater. These include mn3-R, which causes the pitted and miniature phenotypes, su3-R, which causes the sugary- shrunken phenotype, and five other sugary-shrunken mutations found to be alleles of the sul locus.
Lethality arising from most of the mutations was uniform, in that no exceptional germinating kernels were observed. Kernels homozygous for eight muta-
tions, however, germinated at frequencies of up to
5 % (denoted by parentheses in Table 1). Various
phenotypes were observed in the plants resulting from such rare germination events. Examples include three small, pitted kernel mutations in which plant devel- opment was blocked at different stages; prgl-R caused seedling lethality prior to the five-leaf stage, Ptdl-R
blocked development in plants prior to the appearance of the male or female inflorescence, and dek*-PS602
resulted in narrow-leafed, infertile plants. Three mu- tations that caused the ren phenotype, renl-R, ren3- R , and ren2-413, also germinated at rates of less than
5 % . In those rare instances when germination oc- curred the renl-R and ren3-R mutants produced rel- atively normal plants; these were fertile, and upon self-pollination produced 100% mutant kernels. Homozygous ren2-413 kernels, in contrast, produced small plants with rudimentary tassel formation and no ears.
and aleurone mosaic frequently produced seedlings with reduced chlorophyll content (SHERIDAN and
NEUFFER 1980; COE, NEUFFER and HOISINGTON
1987). For example, five putative Mu-induced muta- tions found to be alleles of the dek7, dek5, dek25, and dek21 loci (Table l ) , produced striated or albino seed- lings. Other mutations found to affect seedling green- ing include p r g l - R and dek*-5191 (striated leaves), et2- R (albino seedlings), b r n l - R (pale green seedlings) and p t d l - R (brown, mottled leaves). Furthermore, altera- tions in seedling pigmentation are not limited to seed- lings grown from kernels with obvious aleurone dis- ruption. An allele of the opaque mutation p r o l ( p r o l - 5 1 3 2 ) produced striated seedlings, and dek*-4198, which causes the small seeded phenotype, produced albino seedlings.
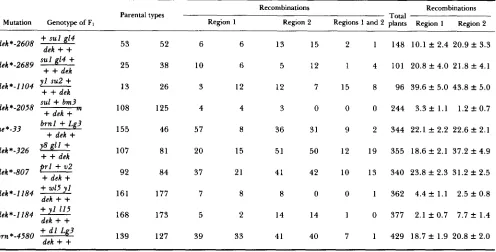
Linkage determinations: Genetic linkage analysis was used to map more precisely the location of several dek loci. Tables 4-6 present the results of two-point, three-point and four-point linkage studies involving 21 different dek mutations; these data are summarized in Table 7. Linkage tests involved marker genes on the particular chromosome arm containing the dek mutation.
In some instances, linkage mapping analyses pro- duced ambiguous results. For example, although mul- tiple positive T B tests located p t d 2 to chromosome 7L, the ptd2-R allele was found to be unlinked to the 7L marker 91, This result might be explained by p t d 2 being located on the extreme distal end of chromo- some 7L. T h e B-A translocation data seemingly are reliable, because the unique phenotype of p t d 2 mu- tants is not readily misclassified and does not resemble the small kernel phenotype often expressed in 7L hypoploid endosperms (ROMAN and ULSTRUP 1951).
A similar situation exists for linkage mapping of dek*-llO4, which was found to be unlinked to both y l (6L: 17) and su2 (6L:58). T h e dek*-llO4 mutation is closely linked to a mutation producing luteus seed- lings, and initial placement was assigned because crosses to TB 6L plants produced both small seeded kernels yielding green seedlings (putative endosperm hypoploids) and large seeded kernels segregating for luteus seedlings (putative embryo hypoploids). If in- deed the placement of this mutant to 6L is correct, the locus must be very distal to the centromere. Small sample size (96 individuals) is unlikely to have allowed inaccurate calculation of the actual percent recombi- nation value, because the linkage observed between su2 and y l is very close to that reported on the 1992 linkage map (41 cM).
Linkage mapping of ref1 was hampered by both the small sample size and large deficiencies in the defec- tive kernel phenotypic classes due to the low fre- quency of mutant expression. Finally, although su3
was placed to chromosome 4 after observing close
linkage to the waxy-marked translocation 4-9g (Table 4), linkage was not detected between su3 and either s u l (43347) or g 1 4 (4L:62) (Table 5), nor was su3-R uncovered by TB-4Sa or TB-4Lf. Mutant phenotypes that result from homozygosity for two, unlinked mu- tations (duplicate factors) are expected to segregate in 15:l (wild type:mutant) ratios on self-pollinated ears of plants heterozygous for each kernel developmental mutation. Since the su3 mutant phenotype is ex- pressed in low frequency approaching 15:1, the mu- tant phenotype may be the result of concomitant homozygosity for duplicate factors, one of which is located on chromosome 4 .
DISCUSSION
This study describes kernel phenotypes and associ- ated pleiotropic effects conditioned by 63 dek muta- tions that arose in a Mutator background. From these mutations we identified 14 novel dek gene loci, as well as new alleles of 10 known dek genes. An important aspect of this work is the potential to study kernel development by a molecular cloning approach, be- cause many of the mutations probably arose by Mu-
tator transposition.
Several of the mutant phenotypes described in this report are of the empty pericarp class, i.e., flattened hulls devoid of both endosperm and embryo tissue. Many of these empty pericarp mutants are likely to have mutations in genes required for viability of all cells types, which are not expressed specifically in kernel tissue. Our phenotypic analysis suggests, how- ever, that other dek mutations described in this study may identify genes that function specifically in the kernel, and are required for its normal development.
Mutator-induced dek mutations are likely to be useful in molecular analyses of kernel development:
Because the new dek mutations were derived from active Mutator stocks, it is likely that many arose from insertion of a Mu element. Thus, the mutations rep- resent a large pool of gene loci required for kernel development that are potentially accessible to molec- ular cloning via transposon tagging (WIENAND et al. 1982). Although the Mutator transposable element system is a large family of at least 13 distinct elements
(BARKER et al. 1984; CHANDLER, RIVIN and WALBOT
1986; TAYLOR and WALBOT 1987; OISHI and FREEL- ING 1987; VARAGONA, FLEENOR and WESSLER 1987; TALBERT and CHANDLER 1988; TALBERT, PATTERSON
and CHANDLER 1989; SCHNABLE, PETERSON and SAE-
DLER 1989; FLEENOR et al. 1990; QIN and ELLINGBOE 1990; HERSHBERGER, WARREN and WALBOT 1991 ; QIN, ROBERTSON and ELLINGBOE 199 1 ; CHOMET et al. 199 1; JAMES et al. 1993), most of the Mu-induced mutations that have been analyzed at the molecular level contain insertions of the 1.4-kb transposon Mu1
Maize Kernel Development Mutants 289 TABLE 4
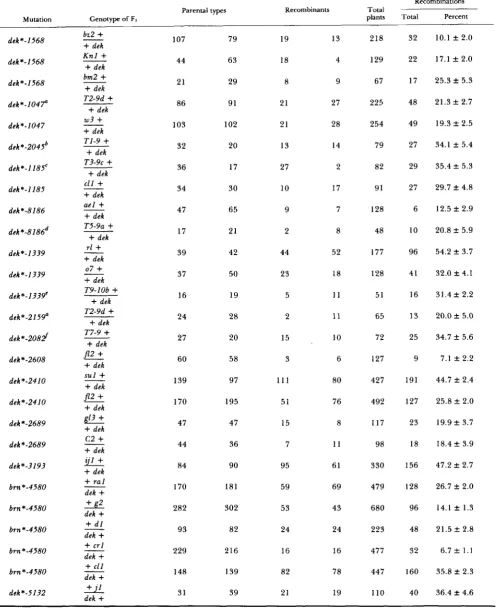
Two-point linkage data
Recombinations Parental types Recombinants Total
Mutation Genotype of FI plants Total Percent
d e k * - l 5 6 8
dek*-1568
dek*-1568
d e k * - l 0 4 P
dek*-1047
dek*-2045'
d e k * - 1 l 8 Z C
dek*-1185
dek*-8186
dek*-8186d
d e k * - 1 3 3 9
dek*-1339 dek*-1339' dek*-2159a dek*-208# dek*-2608 dek*-2410
d e k * - 2 4 1 0
dek*-2689
d e k * - 2 6 8 9
dek*-?193
b m *-4580
b m *-4580
b m * - 4 5 8 0
b r n * - 4 5 8 0
b r n * - 4 5 8 0
d e k * - 5 1 3 2
b z 2
+
+
dek K n l+
+
dek bm2+
-
-
+
dek T 2 - 9 d+
+
dekw?
+
+
dek-
T I - 9
+
+
dekT3-96
+
+
dek c l l+
+
deka e l
+
+
dekT 5 - 9 a
+
+
dek rl+
+
dek 07 +-
-
-
+
dek T9- I Ob+
+
dek T 2 - 9 d+
+
dek T7-9+
-
+
dekJ
?
+
dek s u l+
+ dek
-
J
&
+
dek+
dekc2
+
+
dek-
ijl
+
-
+
dek+
raldek
+
-
-
dek+
+
d l dek+
+
c r l dek+
+ c f l
-
dek
+
+jl
dek
+
107 44 21 86 103 32 36 34 47 17 39 37 16 24 27 60 139 170 47 44 8 4 170 282 9 3 229 148 31 79 63 29 91 102 20 17 30 65 21 42 50 19 28 20 58 97 195 47 36 90 181 302 82 216 139 39 19 18 8 21 21 13 27 10 9 2 44 23 5 2 15 3
1 1 1
51 15 7 95 59 53 24 16 82 21 13 4 9 27 28 14 2 17 7 8 52 18 1 1 11 10 6 80 76 8 1 1 61 69 43 24 16 78 19 218 129 67 225 254 79 82 91 128 48 177 128 51 65 72 127 427 492 117 98 330 479 680 223 477 44 7 110 32 22 17 48 49 27 29 27 6 10 96 4 1 16 13 25 9 191 127 23 18 156 128 96 48 32 160 40
10.1 f 2.0
17.1 f 2.0
25.3 f 5.3
21.3 f 2.7
19.3 k 2.5 34.1 k 5.4 35.4 f 5.3
29.7 f 4.8
12.5 f 2.9
20.8 k 5.9 54.2 f 3.7
32.0 +- 4.1
31.4 f 2.2
20.0 k 5.0 34.7 f 5.6
7.1 f 2.2
44.7 f 2.4
25.8 f 2.0
19.9 f 3.7
18.4 f 3.9
47.2 f 2.7
26.7 f 2.0
14.1 k 1.3 21.5 f 2.8
6.7 f 1.1
35.8 f 2.3
TABLE 4
Continued
Recombinations
Mutation Genotype of F,
Parental types Recombinants
plants Total Total
Percent
su-sh*-5081g
+
T4-9dek
+
28 59 5 1 93 6 6.5 f 2.5dek*-326
2
dek
+
60 92 54 23 229 77 33.6 -C 3.1a waxy-marked translocation 2-9d.
waxy-marked translocation "98918. waxy-marked translocation 3-9c.
waxy-marked translocation 5-9a.
* waxy-marked translocation 9-lob. fwaxy-marked translocation 7.94363.
waxy-marked translocation 4-9g.
TABLE 5
Three-point linkage data
Recombinations Recombinations
Mutation Genotype of FI
Parental types
Region 1 Region 2 Regions 1 and 2 plants Region Total 1 Region 2
dek*-2608
+
sul 14 53 52 6 6 13 15 2 1 148 10.1 f 2.4 20.9 f 3.3dek *-2689 sul g14
+
+
+
dekdek*-1104
-
+ + dek
sul+
bm325 38 10 6 5 12 1 4 101 20.8 -C 4.0 2 1 . 8 f 4.1
y l su2
+
13 26 3 12 12 7 15 8 96 39.6 f 5.0 43.8 f 5.0
dek*-2058 + dek + n 108 125 4 4 3 0 0 0 244 3.3 f 1.1 1.2 f0.7
ae*-33 brnl
+
+
Lg3 155 46 57 8 36 31 9 2 344 22.1 f 2.2 22.6 f 2.1dek
+
dek*-326 + + dekdek*-807
-
p r l+
v 2+
dek+
+ w15
1 y8 g l l+
107 81 20 15 51 50 12 19 355 18.6 f 2.1 37.2 f 4.9
92 84 37 21 41 42 10 13 340 23.8 f 2.3 31.2 f 2.5
dek*-1184 161 177 7 8 8 0 0 1 362 4.4 ? 1.1 2.5 f 0.8
dek*-1184
+
1115 168 173 5 2 14 14 1 0 377 2.1 f 0.7 7.7 f 1.4b m *-4580 + d l L 3
dek
+
+
139 127 39 33 41 40 7 1 429 18.7 f 1.9 20.8 f 2.0tem has gained widespread use as a means of cloning maize loci for which no protein product has been identified. Numerous genes have been cloned using transposon tagging with Mu, including a1 (O'REILLY
et al. 1985), vpl (MCCARTY et al. 1989b), h c f l 0 6
(MARTIENNSEN et al. 1989), bz2 (MCCLAUGHLIN and
WALBOT 1987), y l (BUCKNER, KELSON and ROBERT-
SON 1990) and h m l (JOHAL and BRICGS 1992). T o date, four dek gene loci, d s c l , e m p 2 , r e n l and ren2, have been shown to be tightly linked to specific maize genomic DNA clones isolated based on cosegregation of Mul-containing restriction fragments with the mu- tant phenotype (M. J. SCANLON and M. G. JAMES,
unpublished data).
Applications of genetic characterization of puta- tive Mu-induced kernel mutations: The information
'0
3
w o
CI *&
w %
4 *-
E a : :
e
=
L
L*
g
Maize Kernel Development Mutants 29 1
m z z
.-
& - *
? z z
5 t l i i
2
.g a ? " .
2
3
.E
.H +I"
E
e ' " , : ?E 2
*
2d
2
-
+I +I
.-
5
9
-
2
"
m - L o P- f - C L 5 ; en m
en w
e4
-
z;
.-
L5
2
0
-
m e n
z -
0 0
E "
*
en d m p5-
P-m P-
.- o u
??5 0.I m
z
d - me
2
2
E r n:
-
(D2
8 F D " d - e4 02 %
.-
0.I
a
.- I
E M O
.-
2
s
b o
2
P - ( D
-04
m m
.-
M a
c m o
0 e m
d
.-
??- - m
- L C
-
m w
m m
g g
-
3 5
( D mm m
5
F
' + + +
q;;;
r; $ + - +
M
h
S h P .
'2
F T
c i i
TABLE 7
Summary of linkage aata
Arm Gene locusa location Linkage relationshipsb
emp 1 1 L waxyT1-96
-
34-
empl emf22L empB 2 1
-
waxyT2-9demp2- 19 - w 3
J l
-
46- v4-
8 -em@ - 47 -Chlptdl
-
17 - K n l p t d l-
25 - bm2 ptd 1 1L b22 - 15 - P t d lptd2 7 L ijl - 47
-
ptd2ren 1 5 L p r l
-
24 - renl-
31-
v2a e l - l O - p r 1 - 9 - g 1 8 - 1 9 - r e n l ren2 (ren2-R) 7L ren2
-
34 - ij1y8 - 19 - g l l - 37
-
ren2 dscl 4 s s u l-
3-
dscl - 1-
bm3sul - 3
-
bm3 - 1-
d s c l ren3 1OL r l-
54 - ren307
-
32 - r e d ren3-
32-
waxyT9-10b refl 3 s refl-
30 - c l lwaxyT3-9c
-
35-
reflprgl
-
21 - wzxyT5-9a brnl (brnl-R) 3 s brnl - 7 - crlbrnl
-
22 - d l brnl-
27 - ral brnl - 36-
cll g2 - 14 - brnlbrnl - 19 - d l - 21
-
Lg3 mn3 m n 3 - 4 - w l 5 - 3 - y l 6 Lmn3
-
2-
y l-
8-
115 prol 8 L prol - 29-
j ldek5 (dek5-33) 3 s brnl
-
22 - dek5-
23 - Lg3 dek25 (dek25-2410) 4 s dek25 - 45-
suldek25 - 26 - j l 2 dek7 4 s dek7
-
7 - j l 2d e k 7 - 2 1 - s u l - l O - g l 4 d e k 3 l 4 L dek31
-
20 - g13dek3l - 18
-
c2sul - 21 -g14 - 22
-
dek31or
Prgl 5 L prgl - 13
-
ael or ael-
13 - p r gldek*-2159 2 L dek*-2159
-
20 - waxyT2-9d dek *-20827s
dek*-2082 - 35-
waxyT7-9.363 dek*-1104 6 L y l-
40 - su2-
44-
dek*-llO4 su3 4 s waxyT4-9g - 7-
su3unlinked to sul or g14
a Specific alleles used for mapping are listed in Table 1. In those instances where multiple alleles of a gene were identified in this study, the specific allele used for mapping is denoted in parentheses.
b Data summarized from Tables 4-6.
(or more) independently induced allelic mutations contains a Mu transposon located within the same genetic element, then the likelihood of that element actually causing the phenotype is very high. Several pairs of putative Mu-induced dek alleles were identi-
revertant alleles is not a possible strategy for proving clone identity.
We have reported linkage mapping data for 21 kernel mutations, which reside on 12 different chro- mosome arms. Twenty of these mutations produced inviable, defective kernels. Because of the widespread genetic map locations of these dek genes, they are especially useful for the generation of balanced lethal mutation stocks (MULLER 1918). Balanced lethal stocks are constructed as a means of selecting plants heterozygous for a lethal mutation under study. When plants containing lethal mutations in two different genes closely linked in repulsion are self-pollinated, the viable progeny will be greatly enriched for double heterozygotes due to lethality of any progeny homo- zygous for either of the mutations. T h e detailed ge- netic mapping of several dek loci allows for selection of homozygous-lethal dek mutations best suited for the construction of balanced lethal stocks.
Possible causes of low frequency and altered pat- terns of expression of some mutant phenotypes: Low frequency of expression of the defective kernel phe- notype on self-pollinated ears of heterozygous plants is characteristic of eight mutations described in this study. T h e phenomenon of low frequency expression has been documented in other studies of dek mutations (JONES 1920; CLARK and SHERIDAN 1988) and may be attributed to numerous possible causes. For example, mutant phenotypes resulting from homozygosity at two unlinked, recessive mutations (duplicate factors) are inherited in a 15:l ratio. One mutant phenotype described in this study, attributed to the novel muta- tion su3-R, exhibits an inheritance pattern approach- ing this ratio, indicating this mutant phenotype may indeed result from mutations at duplicate factors (Tables 4 and 5).
Another possible explanation for the observed low frequency of expression is that a dek mutation affects the development or functioning of the male or female gametophyte, such that fewer than the expected 50% of the functional gametophytes (pollen grains or em- bryo sacs) of dek/Dek plants carry the defective allele (CLARK and SHERIDAN 1988). Thus, a reduction in the frequency of mutant kernels on the self-pollinated ears would be observed. In an extreme example, the miniature kernel mutation Mn::Uq disrupts pollen tube growth in the male gametophyte and is male nontransmissible (PAN and PETERSON 1989).
Suppression of Mu-induced inactivation of a locus, as reported by MARTIENSSEN et aZ. (1989; 1990), is another possible explanation for the reduced fre- quency of mutant kernels on ears of self-pollinated plants. Expression of the Mu-induced mutation hcfl06 was correlated with active Mutator stocks. In the sup- pressed state in inactive Mu lines, functional hcfl06 mRNA was transcribed from various initiation sites
within the terminus of the Mu1 transposon in the hcjl06 promoter; this transcription did not occur in Mu-active lines (BARKAN and MARTIENNSEN 1991). Similar genetic behavior was noted for several other mutations, indicating that suppressible alleles may be a common class of Mu-induced mutations (M. FREEL-
ING, unpublished data). Thus, inactivation of the Mu- tator system in individual kernels on an ear segregating for a dek mutation may explain the increased fre- quency of the wild-type phenotype.
Another possible explanation for the observed low frequency of expression is that the mutations may result from Mutator-induced chromosomal deletions that impair gametophyte development or function. MCCLINTOCK (1942; 1944) showed that mutations generated as the result of chromosomal deletions are often transmitted in low frequency through the pol-
len. TAYLOR and WALBOT (1985) and ROBERTSON
and STINARD (1 987) showed that Mutator transposons commonly induce deletions of varying length that can generate stable, null mutant alleles. Thus, some dek mutations that display low frequency of expression may be the result of chromosomal deletions with reduced male transmission.
Pleiotropic effects of dek mutations: Many of the defective kernel phenotypes described in this study and others (NEUFFER and SHERIDAN 1980; SHERIDAN and NEUFFER 1980) indicate a correlation between aleurone disruption in the kernel and altered content of chloroplast pigments in the seedling. These muta- tions include alleles of the dek5, dek7, dek21, dek25, e t l , prgl, b m l , ptdl and et2 genes, and dek*-Mu5191. T h e physiological or biochemical explanation of the link between the aleurone and seedling phenotypes is not known. T h e altered pigment content in the chlo- roplasts of seedlings deficient in these genes may result from nutritional defects caused by altered endosperm storage nutrients and/or improper aleurone function- ing. A more intriguing possibility is that some of the mutations may have a general role in plastid function and/or development. In this view, mutant amyloplast development would cause disruptions in endosperm structure, whereas mutant chloroplast development would lead to discolored, striated or albino seedlings. Abnormalities in chloroplast development have been documented in plants homozygous for the etl-R mu- tation (SANGEETHA et d . 1986). Perhaps the et1 gene and certain other dek genes are required for amylo- plast development as well.
Maize Kernel Development Mutants 293
era1 inviable dek mutations (dek4, dekl2, dekl8, dek29, dek30), when rescued in tissue culture, yielded narrow leafed seedlings. The mutations dek*-PS602 and ptd2-
R both affected leaf and flower formation. Finally, in plants grown from dek*-l364 homozygous kernels, ligule formation, internode length and flower devel- opment all were disrupted. These results indicate that many dek genes play significant roles in the develop- ment of the maize plant, in addition to that of the embryo and endosperm. Detailed analyses of maize leaf development have focused on several dominant, heterochronic leaf mutations that act in the leaf pri- mordium or perhaps earlier, in the shoot apical mer- istem (FREELING 1992). Curiously, there are no iden- tified recessive mutations corresponding to the seven genes identified by their dominant, heterochronic leaf mutations. HAKE (1 992) and FREELINC (1 992) postu- lated the recessive alleles of these genes may be lethal defective kernel or embryo mutations. Possibly several of the inviable mutations described in studies of the defective kernel phenotype are recessive alleles of the heterochronic genes affecting leaf or shoot develop- ment.
We thank S. BRIGGS, N. SHEPHERD and P. S. SCHNABLE for providing some of the defective kernel mutants analyzed in this study, and W. F. SHERIDAN for providing strains carrying numerous known dek mutations. We thank P. S. SCHNABLE for helpful discus- sions and critical reading of the manuscript, and for providing field space for genetic mapping of su3 and et2. This research was s u p ported by U.S. Department of Agriculture research grants 88-
37234-3316 and 91-37301-6344 to A.M.M. and D.S.R. Journal Paper No. 15343 of the Iowa Agriculture and Home Economics Experiment Station, Ames, Iowa, Project No. 3075.
LITERATURE CITED
ANDERSON, E. G., 1956 The application of chromosomal tech- niques to maize improvement. Brookhaven Symp. Biol. 9 23- 36.
BARKAN, A., and R. A. MARTIENNSEN, 1991 Inactivation of maize transposon Mu1 suppresses a mutant phenotype by activating an outward-reading promoter near the end of Mu1. Proc. Nat. Acad. Sci. USA 88: 3502-3506.
BARKER, R. F., D. V. THOMPSON, D. R. TALBOT, J. SWANSON and J. L. BENNETZEN, 1984 Nucleotide sequence of the maize transposable element MuZ. Nucleic Acids Res. 12: 5955-5967. BECKETT, J. B., 1978 B-A translocations in maize. I. Use in locat-
ing genes by chromosome arms. J. Hered. 6 9 27-36.
BUCKNER, B., T. L. KELSON and D. S. ROBERTSON, 1990 Cloning of the y l locus of maize, a gene involved in the biosynthesis of carotenoids. Plant Cell 2: 867-876.
CHANDLER, V. L., C. RIVIN and V. WALBOT, 1986 Stable non-
Mutator stocks of maize have sequences homologous to the Mu1
transposable element. Genetics 114 1007-1021.
CHANDLER, V. L., and K. J. HARDEMAN, 1992 The M u elements of maize. Adv. Genet. 30: 77-122.
CHOMET, P., D. LISCH, K. J. HARDEMAN and V. L. CHANDLER, 1991 Identification of a regulatory transposon that controls the Mutator transposable element system in maize. Genetics
CLARK, J. K., and W. F. SHERIDAN, 1986 Developmental profiles of the maize embryo-lethal mutants dek22 and dek23. J. Hered. 77: 83-92.
129: 261-270.
CLARK, J. K., and W. F. SHERIDAN, 1988 Characterization of the two maize embryo-lethal mutants rgh*-1210 and j7*-1223E:
effects on embryo and gametophyte development. Genetics 120 279-290.
CLARK, J. K., and W. F. SHERIDAN, 1991 Isolation and character- ization of 5 1 embryo-specific mutations of maize. Plant Cell 3:
COE, E. H., M. G. NEUFFER and D. A. HOISINGTON, 1987 The Genetics of Corn, pp. 81-258 in Corn and Corn Improvement,
edited by G. F. Sprague. American Society of Agronomy, Madison.
FLEENOR, D., M. SPELL, D. S. ROBERTSON and S. WESSLER, 1990 Nucleotide sequence of the maize Mutator element,
Mu8. Nucleic Acids Res. 18: 6725.
FREELING, M., 1992 A conceptual framework for maize leaf de-
HAKE, S., 1992 Unraveling the knots in plant development. T I C
HERSHBERGER, R. J., C. A. WARREN and V. WALBOT, 1991
Mutator activity in maize correlates with the presence and expression of M u 9 , a new M u transposable element. Proc. Nat. Acad. Sci. USA 88: 10198-10202.
JAMES, M. G., M. J. SCANLON, M. QIN, D. S. ROBERTSON and A. M.
MYERS, 1993 DNA sequence and transcript analysis of tran- sposon MuA2, a regulator of Mutator transposable element activity in maize. Plant Mol. Biol., IN PRESS.
JOHAL, G. S., and S. P. BRIGGS, 1992 Reductase activity encoded by the H M l disease resistance gene in maize. Science 258: 985- 987.
JONES, D. F., 1920 Heritable characters of maize IV. A lethal factordefective seeds. J. Hered. 11: 161-167.
MANGELSDORF, P. C., 1923 The inheritance of defective seeds in maize. J. Hered. 14: 119-125.
MANGELSDORF, P. C., 1926 The genetics and morphology of some endosperm characters in maize. Conn. Agric. Exp. St. Bull. 279: 509-614.
MARTIENNSEN, R. A., A. BARKAN, M. FREE LING^^^ W. C. TAYLOR, 1989 Molecular cloning of a maize gene involved in photo- synthetic membrane organization that is regulated by Robert- son’s Mutator. EMBO J. 8: 1633-1639.
MARTIENNSEN, R. A., A. BARKAN, W. C. TAYLOR and M. FREELING, 1990 Somatically heritable switches in the DNA modification of M u transposable elements monitored with a suppressible mutant in maize. Genes Dev. 331: 343.
MCCARTY, D. R., C. B. CARSON, M. LAZAR and S. C. SIMONDS, 1989a Transposable element-induced mutations of the viui- parous-1 gene in maize. Dev. Genet. 1 0 473-481.
MCCARTY, D. R., C. B. CARSON, P. S. STINARD and D. S. ROBERT- SON, 1989b Molecular analysis of viviparous-1: an abscisic acid-insensitive maize mutant. Plant Cell 1: 523-532.
MCCARTY, D. R., T. HATTORI, C. B. CARSON, V. VASIL, M. LAZAR and I. K. VASIL, 1991 The viviparous-1 developmental gene of maize encodes a novel transcriptional activator. Cell 6 6
MCCLAUGHLIN, M., and V. WALBOT, 1987 Cloning of a mutable
br2 allele by transposon tagging and differential hybridization. Genetics 117: 771-776.
MCCLINTOCK, B., 1942 Maize Genetics. Carnegie Inst. Wash. 41: 181-186. Year Book
MCCLINTOCK, B., 1944 The relation of homozygous deficiencies to mutations and allelic series in maize. Genetics 2 9 478-502. MULLER, H. J., 1918 Genetic variability, twin hybridsand constant
hybrids in a case of balanced lethal factors. Genetics 3: 422- 499.
NEUFFER, M. G., and W. F. SHERIDAN, 1980 Defective kernel mutants of maize. I . Genetic and lethality studies. Genetics 9 5
NEUFFER, M. G., M. T. CHANG, J. K. CLARK and W. F. SHERIDAN, 935-951.
velopment. Dev. Biol. 153: 44-58.
8 109-1 14.
895-905.