CHANGES IN QUANTITATIVE TRAITS OF TRIBOLIUM UNDER IRRADIATION AND SELECTION1
ALAN C. BARTLETT2, A. E. BELL A N D V. L. ANDERSON Population Genetics Institute, Purdue University, Lafayette, Indiana
Received May 9, 1966
ELECTION is recognized as a major force in the evolution of domesticated species. Yet when experimental populations are subjected to continuous selec- tion for long periods, responses to selection become refractory in spite of demon- strable genetic variation. While differing in details, the general pattern has been a linear response in the early generations followed in the long term by plateaus in the response curve (see LERNER, 1954, for a comprehensive review of the problems).
Several methods such as crossing of highly selected strains, environmental stress, change in system of mating, new criteria of selection and enhanced genetic variability by mutagenic agents have been suggested to circumvent plateaus in selection response. This paper is concerned with the effects of X-radiation on populations under artificial selection. In plants OKA et al. (19.58) and GREGORY (1961) have shown that X-radiation increased usable genetic variability in rice and peanuts, respectively. The most extensive investigation of the problem with animals has been that of SCOSSIROLI (1954a, b, 1959). In replicated studies of bristle number in Drosophila, he found increased genetic variability and greater response to selection in irradiated isogenic, hybrid and plateaued populations than in nonirradiated controls. On the other hand, CLAYTON and ROBERTSON
(1955, 1964) in reporting on two series of Drosophila experiments did not con- firm SCOSSIROLI’S findings.
The aim of the present paper is to study the effects of selection and X-radiation on quantitative traits in another laboratory organism, Tribolium castaneum. A factorial experiment will be reported which extended over 11 generations and involved two strains, three levels of irradiation and two kinds of selection. The selected trait was body weight at the pupal stage. Correlated responses in repro- ductive fitness were observed as number of offspring per mating, percent sterility and adult mortality.
MATERIALS A N D METHODS
Strains: Two strains of T . castaneum differing in their previous selection history but related in origin were chosen for this study. Representative pupae from the two strains are shown in Figure 1. The ‘‘Foundation Strain” originated in 1954 with the systematic crossing of eight
Journal Paper No. 2633, Purdue University Agricultural Experiment Station. Based on a dissertation submitted by
the senior author in partial fulfillment of the requirements for the Doctor of Philosophy degree at Purdue University. This investigation was supported by research contract AT(ll-1)-965 from the Atomic Energy Commission.
700 A. c. BARTLETT et al.
FIGURE 1.-Representatiw ~)upiic froin t l i r Imsc strairis il.;cd i n this study-Large Strain helow and Foundation Strain abovc.
laboratory stocks obtained from various laboratories in the United States. Since then this strain has been reproduced in mass with approximately 250 parents per generation. T h e “Large Strain” used in the present study and pictured in Figure 1 originated in 1955 from Foundation, but since that time it had been maintained as a closed population under selection (individual and family merit) for increased pupal weight. Response of this population during the first 16 generations was reported in 1958 (BELL and MOORE). The response curve remained linear until a decline in reproductive fitness caused it to plateau a t about the 22nd generation. Fitness improved during a period of relaxed selection and further response to high selection was realized. A second and higher plateau in response was evident hy the 44th generation. Again, selection was relaxed and the population was mass mated for eight generations before the present study was initiated.
Cufturing conditions: Standard culturing procedures for Tribolium were used in this experi- ment. Four grams of media consisting of 95% whole wheat flour and 5% hrewers yeast were placed into a creamer (20 ml glass bottle with cardboard pull-cap) for each single-pair mating. A 44-hour egg collection in fresh media was made from each mating about ten days after adult emergence to obtain offspring of the next generation. Temperature was maintained a t 33* 1°C
and humidity was held at 70 f 2% relative humidity.
Irradiation. Radiation treatments were provided by an X-ray machine operated a t 30 kv and 20 ma. The distance from target to the insects was one meter. The beam was filtered with 1 mm of copper plus the inherent filtration of the tube (0.25 m m copper).
Experimental design: The design of the experiment was a 2 x 2 x 3 factorial arrangement of treatments with two strains, Foundation and Large; two levels of selection, high random; and three levels of radiation, 0, 100r, and 1000r. Thus, six treatment groups (lines) per strain as summarized in Figure 2 were established and maintained as distinct closed populations with the specified treatments repeated each generation for 1 1 consecutive generations.
Each experimental line or subpopulation originated from 50 single-pair matings randomly taken from its base strain (Foundation or Large). Reproduction of a line in each subsequent
Foundetion Strain (Ir) Large strain (L)
--
n i w pcr Rnndom 11,iKll Rnndom Iligh
gcnrration (r) selection (R) selection (S) selection (R) selection IS)
0 (0) FOR FOS LOR LOS
100 (C) FCR FCS LCR LCS
(M) F M R FMS LMR LMS
IRRADIATION A N D SELECTION 70 1 generation was by 50 single-pair matings with mates being taken at random (barring full-sibs) from ten selected families. In the random selection lines (Rj, the ten selected families were chosen at random. High selection (S) was f o r increased pupal weight and was based on family merit with a planned selection intensity of 20%. If any of the 50 matings were sterile, the selec- tion intensity was lowered to provide the requisite ten selected families.
To allow comparisons of the immediate effects of radiation on reproduction of treated indi- viduals with the accumulated effects transmitted from irradiation of previous generations, the number of progeny per mating was measured each generation in a #-hour egg collection (pre- radiation) taken just before irradiation of the parents and a second 4-8-hour collection (post- radiation) taken immediately after. When the majority of the progeny had pupated (19 days following egg collection for the foundation lines and 21 days for large lines) the offspring for each mating were collected and counted by passing the media through a 20-gauge screen. Measurements on the trait of selection, pupal weight, were taken only on progeny from the postirradiation collection. Three males and three females were taken at random from each mating and weighed as a group to the nearest 1/100 mg on a micro-analytical balance. The remaining progeny per mating were saved to be included for future matings in case their family was selected. When the reproductive fitness declined in any line to a point that the ten selected families did not yield the 50 males and 50 females required f o r new matings, the selection intensity was relaxed to include additional families. The degree to which this occurred was recorded and will be reported.
RESULTS
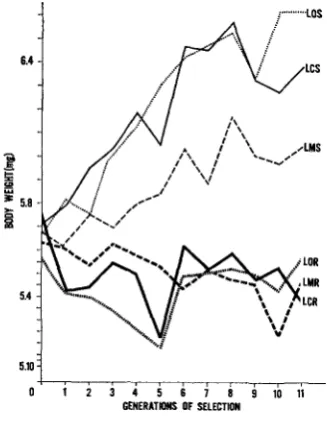
The observed response of the selected trait, pupal weight, in each of the experi- mental lines initiated from the Foundation Strain is shown in Figure 3. Com- parable responses for the Large Lines are presented in Figure 4. In order to examine statistically early versus late responses the generations were divided into
4 , , , , , , , , , , ,
0 1 2 3 4 5 6 7 8 9 l O l l LIKRATHIWS OF SELECTW
FIGURE 3.-Selection response in the Foun- dation Strain to various combinations of arti- ficial selection and X-irradiation. (See Figure 2 f o r line code. j
5.m
_ , I
0 1 2 3 4 5 6 1 8 9 1 0 1 1 CENERATiVNS OF SELECTWJN
702 A. c. BARTLETT et
al.
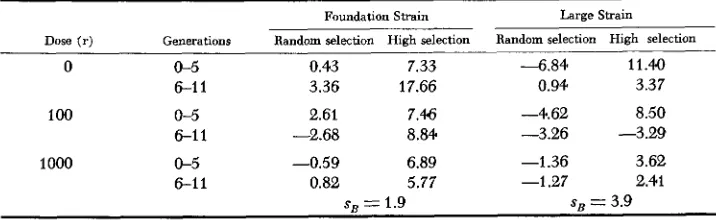
early (0-5) and late (6-11) periods. Response within each line was investigated by the usual least squares procedure and the deviations from linear regression were found to be not significant at the .10 probability level. Hence the linear trend was accepted for all combinations of strains, doses and selections in each of the periods and are summarized in Table 1.
Deviations of response from linear regression were homogeneous among lines within each strain, but heterogeneous across strains, Therefore, analyses of regression coefficients as presented in Table 2 were made separately for the two strains. It is not surprising that “Selection Methods” (Random us. High Selection) identifies a major source of variation among responses for both strains. The response of pupal weight to other factors was not so consistent across both strains.
An examination of the response for the Foundation Lines reveals that the significant overall effect of “&ses’’ resulted primarily from the greater response of the selected line receiving no irradiation. In fact all lines subjected to high selection (FOS, FCS, FMS) responded equally well through the early generations, but in the later generations the population receiving no irradiation (FOS) showed a significantly greater response by DUNCAN’S test (1955) than either of the other selected lines. The response of the high selection line receiving 1OOr (FCS) during this late period was intermediate while that for FMS was little more than a third
of that observed for FOS. The exceptionally large response for FOS in the late period also reflects in the highly significant interaction mean square found for Periods x Doses in Table 2.
No consistent radiation effects on pupal weight were observed between Founda- tion Lines subjected to random selection (FOR, FCR, and FMR). The slight positive trend for FOR was statistically significant when measured over all generations and could reflect a positive trend in the environment.
As .indicated before, the responses of the various experimental lines originating from the Large Strain (Figure
4)
were more variable from generation to genera- tion than those of corresponding Foundation Lines. Yet, again the high selection line receiving no radiation (LOS) showed the greatest overall response even though not statistically significant by DUNCAN’S test. The line receiving mild radiation (LCS) responded equally well as LOS during the early generations,TABLE 1
Average selection responses (10-f mg) per generation for various lines during early (0-5) and late (6-11) generations (estimated as regression of mean pupal weight on generation of selection)
Foundation Strain Large Strain
Dose (r) Generations Random selection High selection Random selection High selection
0 0-5 0.43 7.33 - 6 . 8 4 11.40
6-1 I 3.36 17.66 0.94 3.37
1 0 0 0-5 2.61 7.46 -4.62 8.50
6-1 1 -2.68 8.84 -3.26 -3.29
1000 0-5 -0.59 6.89 -1.36 3.62
6-1 1 0.82 5.77 -1.27 2.41
IRRADIATION A N D SELECTION 703
TABLE 2
Analysis of linear regression coefficients by strain for responses in pupal weight during
early and late generations of selection for various treatment groups
Mean squares
~-
Degrees of
Source of variation f i eedom Foundation Large
Selection Methods 1 215.53** 150.01**
Periods 1 9.20 11.56
Doses 2 15.58* 8.34
Selection x Periods 1 9.53 76.33**
Selection x Doses 2 5.86 9.22
Periods x Doses 2 22.25** 7.95
Selections
x
Periods x Doses 2 6.96 14.92Error 4 3 3.60 15.24
_ _ _ _ _ ~ _ _ _ _ _ _ _ _ _ ~ * Significant a t 5 % level. * * Significant at 1% level.
but response in the late generations fell behind. The Large Line receiving 1000r per generation (LMS) responded poorly to selection from the start of the experi- ment.
No evidence for a positive trend in pupal weight was found in any of the ran- domly selected Large Lines (LOR,
LCR,
LMR).
I n fact, the significant Selectionx
Periods interaction reported for Large Lines in Table 2 was due, in part, to all random lines declining more in pupal weight during the early period than during the late generations. While none of the individual regression coefficients for the Large Random Lines differed significantly from zero, the overall trend is a negative one.Factors aflecting response to selection: In studying the effects of any external
agent on response to selection a deeper insight might result if one examines the basic elements determining the response. Theoretically, this response can be predicted as R =
i
oph2, where R is the response to selection, i is the selection intensity (standardized selection differential), up is the phenotypic standarddeviation and h' is heritability of the selected trait. Data were collected in this experiment relative to these variables.
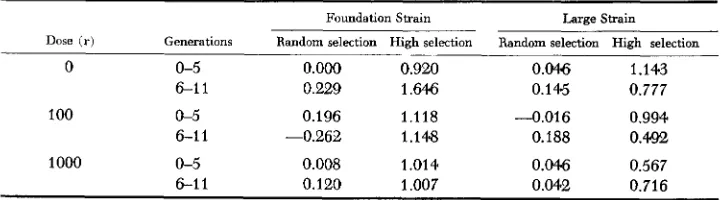
The selection intensities for the various lines are summarized in Table 3. Since there was a n indication from the response curves that some lines responded differ-
TABLE 3
Average selection intensities for pupal weight b y lines for early and late generation
Foundation Strain Large Strain
Dose ( r ) Generations Random selection High selection Random selection High selection
0 0-5 O.OO0 0.920 0.0% 1.143
6-1 1 0.229 1.646 0.145 0.777
100 0-5 0.196 1.118 -0.016 0.994
6 1 1 -0.262 1.148 0.188 0.402
1000 0-5 0.008 1.01 4 0.046 0.567
704 A. c. BARTLETT et al.
ently late in the experiment in contrast to the early generations, the average selection intensities are reported separately for these two periods. An analysis of this variable revealed strains, selection methods and strains by methods inter- action to be significant sources of variation.
The selection intensities for all of the randomly selected lines were small (as intended) and reflected little more than sampling variation. However, the selec- tion intensities for both sets of high selection lines are found to be related directly to differences among selection responses observed in Table 1. For example, the accelerated response in late generations for FOS was associated with a more than
50% increase in the average selection intensity. A close comparison of these intensities with corresponding observed responses within each strain reveals very close to a one to one relationship.
The second element in the above prediction equation is the phenotypic standard deviation ( u p ) a statistical measure of phenotypic variability within each line. Since selection f o r pupal weight in this study was based each generation on differ- ences among family means within each line, phenotypic variance (0:) among
these means were calculated for each generation per line. BARTLETT'S test rejected the hypothesis of homogeneity among these phenotypic variances. The possibility that this heterogeneity was associated with either irradiation or selection merited further study.
Since the distribution of the logarithm of variances of normally distributed variables is approximately normal (BARTLETT and KENDALL 1946)
,
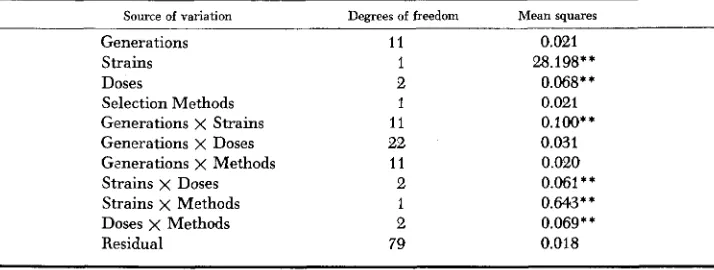
the pheno- typic variances were transformed to logarithms for analysis. Box (1 954) has shown for similar conditions that the robustness of the F-test keeps the probability of the Type 1 error nearly constant. The resulting analysis of variance, among logarithms of phenotypic variance f o r family mean pupal weight within genera- tion and treatment group, is presented in Table 4.TABLE 4
Analysis of phenotypic variation for pupal weight within the various treatment groups (variance among family group means within generation and treatment groups
transformed to logarithms for analysis of variance)
Source of variation Generations Strains Doses
Selection Methods Generations x Strains Generations x Doses Generations x Methods Strains x Doses Strains x Methods Doses x Methods Residual
Degrees of freedom
11 1 2 1 11 22 11 2 1 2 79
Mean squares
0.021 28.198**
0.068** 0.021 0.100** 0.031 0.020 0.061 * *
0.643** 0.069**
0.018
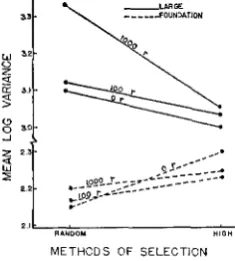
I R R A D I A T I O N A N D S E L E C T I O N 705 While the significant Strain effect on phenotypic variance might be attributed to scaling, the significant mean square due to Doses is of direct interest to the present study. It is interesting that the overall effect of Selection Methods was not significant, yet a highly significant Strain
x
Selection Methods interaction existed. This interesting interaction is graphically shown in Figure 5 where one sees phenotypic variance increase under high selection for Foundation Lines while the opposite situation resulted for all Large Lines. Furthermore, it can be seen in Figure5
that unirradiated Foundation Lines (Or) were the major contributors to the significant Strains x Doses and Dosex
Methods interactions. The phenotypic variance increases with level of irradiation in both strains and selection methods with the exception of Line FOS. While these effects of strain, method of selection and level of irradiation on phenotypic variation are of con- siderable interest in themselves, it is obvious that differences in selection responses (Table 1, Figures 3 and 4) are not directly related to differences in phenotypic variances. In fact, with the exception of FOS, the responses in both strains from high selection were inversely related to phenotypic variances.Heritability (h')
,
the third element determining response to selection, will now be examined in relation to selection and irradiation. Theoretically, it is possible for either irradiation or selection to effect an increase or decrease in heritability. Since matings were made each generation while the subsequent parents were in the pupal stage, it was possible to determine mid-parental pheno- types by weighing each of the 50 pairs. An estimate of heritability for each line and generation was made from the regression of mean offspring weight on mid- parental weight. These estimates were pooled to yield the average heritability per line shown in Table5.
Before relating the h' values to selection responses, it is of interest to check for trends in regard to the external factors of irradiation and selection. The herit- ability values for high selection lines in both strains were significantly lower than corresponding randomly selected lines. This supports the theoretical expectation of reduced genetic variation i n closed populations under continued selection for
3
'2
2 2
RANDOH " l o "
2 ,
M E T H O D S OF S E L E C T I O N
706 A. c . BARTLETT et al. TABLE 5
Heritabilities of pupal weight and t h i r standard errors as estimated by pooled regression of offspring on mid-parent
Foundation Strain Large Strain
Dose (r) Random selection Hight selection Random selection High selection
0 0.439 f 0.054 0.318
*
0.045 0.338 f 0.077 0.316 k 0.055100 0.413
*
0.057 0.408 f 0.057 0.488*
0.067 0.333+
0.0681000 0.365 ? 0.058 0.356 & 0.064 0.456 f 0.059 0.315 f 0.059
a single trait. The effects of irradiation on heritability are of greater interest to this study, but a general trend is not obvious.
A comparison of the hz values in Table 5 with high selection responses (Figures
3 and 4) reveals that differences in response are not due to differences in herit- ability. For example, lines
with
the highest response (FOS and LOS) had two of the lowest average hZ values. Thus differences in selection intensities remain the major factor related to the observed differential responses to selection.Reproductive fitness: The design of this experiment called f o r 50 single-pair matings each generation in each of the 12 experimental lines. In order to main- tain population size and to minimize inbreeding, the 50 males and 50 females for matings were to be taken equally from ten selected families. Thus, if all matings in each experimental line were fertile, the proportion of families selected would be a uniform 10/50 or 20%. However, if the reproductive fitness declined in any particular line, the intensity of selection would decline in order to meet the established selection criterion.
A 48-hour egg collection was taken under uniform conditions from each of the
50 matings within each line immediately before and after exposing the mated pairs to their respective radiation treatment (0, 100r or 1000r). After the first generation, these prospective parents for the irradiated lines could manifest ge- netic effects accumulated in their particular line from irradiation in previous gen- erations. Obviously, the broad category of dominant lethals (genetic or somatic) would not be represented in the first egg collection. The average number of “pre- radiation” progeny (solid line) and number of “postradiation” progeny (dotted line) per mating each generation are graphically presented in Figure 6 for the Foundation Lines. Similar data for the Large Lines are shown in Figure 7.
The first graph in each row represents a comparison between preradiation and postradiation collections for unirradiated lines. No difference would be expected here other than a possible age effect due to the females being 48 hours older for the second collection. While considerable variation was observed between generations, the two collections within any generation were very similar as evidenced by the small “G x C” interaction in Table 6.
IRRADIATION A N D S E L E C T I O N 707
40
30
40
30
20
10
0
E
E o
FMS
-
I I I I I I I I I I I 2 3 4 5 6 7 8 9 IO 40 I
0
-0 1 2 3 4 5 6 7 8 9 1 -0
I
FCSl o t
0
-0 1 2 3 4 5 6 7 8 9 1 0
..-...
..
..
. .
2 0 - ,:.-. ... . . . . j
: . .: :.
.,..
./'
10
-
0 I 2 3 4 5 6 7 8 9 IO II 0 I 2 3 4 5 6 7 8 9 IO GENERATIONS OF SELECTION
FIGURE 6.-Number of progeny per mating in preradiation (-) and postradiation ( - - - )
collections by generations for all Foundation Lines. (See Figure 2 for line code.)
accumulated deleterious effects of irradiation on fitness can be seen from a com- parison of preradiation progeny numbers (solid line) in successive graphs of each row. In general, one sees a decrease in numbers of progeny as he moves from unirradiated lines to those receiving 100r each generation and to those receiving 1000r. Yet within any irradiated line the decline over generations is hardly noticeable and certainly is not proportional to the accumulated dose. Apparently, natural selection within these populations was eliminating deleterious mutations rapidly enough to prevent their build-up over generations.
While Figures 6 and 7 give an overall view of the effects of irradiation and selection on reproductive fitness in the two strains, the results are of sufficient interest to merit a more detailed analysis. The following components of fitness will be examined: ( 1 ) number of off spring per fertile mating, (2) percent sterility among matings and (3) percent mortality among prospective parents.
Fertile matings were defined as those matings which produced one or more offspring in either of the two collections (pre- or postradiation). Even though 50
708 A. c. BARTLETT et al.
40
I
40r
30’
40 I 1l o t
0
0 1 2 3 4 5 6 7 8 9 l O l l
lCS
0
-0 I 2 3 4 5 6 7 8 9 1 -0
40
I
0
0 1 2 3 4 5 6 7 8 9 1 0 1 1
30
t
40
LMR
30
-
. . -.... :
0 I I I I I I I I I I 0 1 2 3 4 5 6 7 8 9 1 0 1 1
GENERATIONS OF SELECTION
FIGURE 7.-Number of progeny per mating in preradiation (-) and postradiation (-
-
-)collections by generations for all Large Lines. (See Figure 2 for line code.)
found not to be heterogeneous and a combined analysis of variance on the average number of offspring per fertile mating per treatment combination was made. The analysis as presented in Table 6 treated “collections” (pre- and postradiation) as subplots to the factorial combinations of the main factors of strains, selection methods, doses and generations.
One finds in Table 6 that all the main sources of variation significantly in- fluenced this component of fitness. Reductions in numbers of offspring were associated with the Large Strain, high pupal weight selections and higher levels of irradiation. The significant mean square for Generations appeared to be ran- dom environmental time fluctuations and of no significance in the present study since all treatment groups were exposed simultaneously. The significant inter- actions of Strains with both Selection Methods and Doses are of special interest in reference to the previous selection histories of these strains. Trends evident in Figures 6 and 7 indicate that the significant S
x
M interaction arises from the Large Lines being more sensitive to the deleterious effects of high selection. On the other hand, greater sensitivity of the Foundation Lines to the deleterious effects of irradiation lead to the S x D interaction.I R R A D I A T I O N A N D S E L E C T I O N 709 TABLE 6
Analysis of number of progeny in preradiation and postradiation collections from fertile m t i n g s in various treatments and over generations
Source of variation
Degrees of freedom Strains (S)
Selection Method (M) Doses (D)
Generations (G)
S X D S X M
S X G M x D M x G D x G S X M X D Residual (error “a”)
Collections (C)
s x c
M x C D x C G x C S X M X C S X D X C Residual (error “b”)1 1 2 11 1 2 11 2 11 22 2 77 1 1 1 2 11 1 2 123 Progeny numbers mean square 36.863** 6.064* *
27.974** 2.460** 2.591 ** 3.793** 0.537 0.492 0.197 0.363 2.278* * 0.191 19.096** 0.820** 0.215 9.101** 0.029 0.793** 0.568** 0.059
* * Significant at 1% level.
mation. The significant mean square for iCollections simply confirms the fact that the postradiation collection had less offspring per mating than was observed in the 24-hour collection made before radiation exposure. Likewise, the Dose x
Collection interaction is a natural consequence of differences between collections being a function of dose (0, 100, or 1000r). Of greater interest to the geneticist are the significant first and second order interactions involving strains. While both strains reflected a reduction for number of offspring in the post-radiation collection, these significant interactions indicate that the strains react differently to the immediate effects of irradiation. It can be seen in Figures 6 and 7, that, in general, the Foundation Lines suffered the greater reduction in numbers of off- spring from irradiation each generation.
710 A. c . BARTLETT et al. TABLE 7
Analysis of proportional radiation resistance (ratio of the difference in progeny numbers per mating between pre- and postradiation egg collections to total progeny per mating), sterility (arc sin in percent) and mortality (arc sin percent) in various treatment groups
Degrees of Radiation resistance Sterility Mortality Source of vairation freedom mean squares mean squares mean squares
Strain
Selection Methods Dose
Generations
S X D
S X M
S X G M x D M x G D x G Residual 1 1 2 11 1 2 11 2 11 22 79 .1196** ,0154 1.0249**
.ow
.0604* .0904** .0912** ,0017 .0141) .0149 ,0088 3.566** 0.535** 2.233** 0.133' *0.843* * 0.21 5* *
0.087** 0.040 0.030 0.041) 0.040 1.391** 0.124** 0.393** 0.115** 0.065 0.001 0.043 0.010 0.026 0.026 0.01 7 ~
* Significant at 5% level. * * Significant at 1 % level
Therefore, the hypothesis of a proportional reduction to account f o r differential resistance to radiation damage can be rejected. The significant mean squares due to Doses is an obvious manifestation of radiation damage, while the significant interactions of Strains with the other main effects further confirm the earlier point that environmental effects are conditioned by the genetic background of the treated material.
IRRADIATION A N D SELECTION 71 1
TABLE 8
Average percent sterile matings and percent mortality (pupa to 14-G?~Y imago) in each strain and treatment combination over generations
Dose (r)
Percent sterile matings 0 100 1000 0 100 1000 Percent mortality
Foundation Strain
Random selection High selection
6.8 4.5
13.7 11.8
22.8 23.7
1.2 1.5
3.8 2.8
4.2 4.9
Large Strain
Random selection High selection
15.3 24.3
15.2 31.8
28.7 36.5
3.7 6.3
5.8 10.6
8.8 9.4
DISCUSSION
The induction of new and favorable genetic variation in predominantly homo- zygous material by ionizing radiation has been amply demonstrated (e.g. GREG-
ORY 1961). In fact, mutation breeding is an accepted adjunct to hybridization in the genetic improvement of self fertilized species.
Genetic variation per se is seldom a problem in the improvement of normally cross fertilized species. The primary concern has been with mating and selection techniques that maximize genetic gain or selection response. However, a decline in additive genetic variation frequently follows long term selection even though significant amounts of nonadditive genetic variation remains in such populations
(BROWN and BELL 1961 )
.
Thus the induction or release of usable genetic variation in such “plateaued” populations by ionizing radiation is a possibility worthy of study. Favorable results obtained with homozygous material may not be appli- cable to the heterogenic situation which normally prevails in animal populations. The limited amount of published work on the latter case needs confirmation and extension to a wider range of genetic situations.The factorial nature of the experiment reported here allowed examination of the effects of X-radiation separately and in conjunction with artificial selection for increased pupal weight in two genetically dissimilar populations of Tribolium custuneum. The predictive factors (selection intensity, heritability and pheno- typic variation) contributing to selection response over the course of l l genera- tions were examined in each treatment group or line. Also, reproductive fitness in terms of number of offspring, sterility and adult mortality was observed.
712 A. e. BARTLETT et al.
observed in this study declined somewhat proportionally with increasing levels of irradiation.
Obviously, any generalization regarding the usefulness of artificial irradiation in a selection program for an outbreeding species must recognize the biology of the experimental organism plus the specific experimental design applied. If the reproductive capacity of Tribolium had been such that the deleterious effects of irradiation were not reflected in a reduced selection intensity, the outcome of the present study might have been quite different. Likewise, a different experimental design, whereby irradiation treatments were followed by one or more generations of natural selection or whereby a uniform selection intensity was required, could have changed the relative selection responses observed.
In contrast to similar responses discussed above, other lines of evidence revealed the two strains reacting differently to irradiation and selection. For example, high selection for pupal weight without irradiated (FOS) in the Foundation Strain was accompanied by a n increase in reproductive fitness, higher selection intensities and a n enhanced selection response in the late generations. Yet the same selection procedure applied to the Large Strain resulted in lowered fitness and reduced selection intensities with a corresponding decline in selection response. Also, irradiation without selection was associated with some increase in pupal weight for Foundation Lines while comparable Large Lines showed an overall decrease. Since the Large Strain had originated from Foundation, the selection history of the two strains previous to the present study is a prime suspect for the differential patterns of response.
One additional point of interest observed in the present study is that the Large Strain was more resistant to both immediate and accumulated deleterious effects of X-irradiation than was the Foundation Strain even though the latter had a higher overall fitness. This confirms our earlier finding regarding these two populations (BARTLETT and BELL 1962).
If .one considers X-radiation and high selection for pupal weight as environ- mental stresses applied to both strains, a strain by treatment interaction was mani- fested for all components of fitness. That is to say that the strain affected least in terms of fitness by selection (Foundation) was the most susceptible to irradi- ation damage.
SUMMARY
The consequences of various combinations of selection (high and random) and levels of X-irradiation (0, 100, and 1 OOOr per generation) on pupal weight were investigated in two strains of
Tribolium
castaneum for 11 generations. Repro- ductive fitness was measured each generation in terms of sterility, adult mortality and number of offspring. The Foundation Strain was an unselected panmictic population with an initial weight of 2.10 mg. The second strain, Large, had originated earlier from Foundation and had been selected for large pupal weight for 4 4 generations. At the beginning of the present study its average pupal weightwas 5.66 mg.-High selection contributed to significant responses in pupal
IRRADIATION A N D SELECTION 713 irradiation. Even though an increase in heritability and phenotypic variance was observed in some irradiated lines, a decline in reproductive fitness contributed to smaller selection differentials and in turn less selection response.-Several strain by treatment interactions possibly were related to the previous selection history of the genetic material. For example, reproductive fitness increased with high selection in Foundation and responded inversely in the Large Strain. Yet the latter was more resistant to the deleterious effects of irradiation than was the Foundation Strain.
LITERATURE CITED BARTLETT, A. C., and A. E. BELL, 1962
BARTLETT, M. S., and D. G. KENDALL, 1946
Bell, A. E., and C. H. Moore, 1958
The effect of irradiation on reproduction in two strains
The statistical analysis of variance-heterogenic and log transformation. J. Royal Stat. Soc. Ser. 8 : 128-138.
Further comparison of reciprocal recurrent selection with conventional methods of selection for the improvement of quantitative characteristics.
(Abstr.) Proc. 10th Intern. Congr. Genet. 2: 20-21.
Effect of inequality of variance in one-way classification. Ann. Math. Statist. 25: 29g302.
Genetic analysis of a plateaued population of Drosophila melanogaster. Genetics 46: 407-425.
Mutation and quantitative variation. Am. Naturalist
89: 151-158.
-
1964 The effects of X-rays on quantitative characters. Genetic Res.5 : 410-422. DUNCAN, D. B., 1955 GREGORY, W. C., 1961
LERNER, I. M., 1954
OKA, H.-I., K. HAYASHI, and I. SHIOJIRI, 1958
SCOSSIROLI, R. E., 1954
of Tribolium castaneum Herbst. Radiation Res. 17: 864-877.
Box, G. E. P., 1954
BROWN, W. P., and A. E. BELL, 1961
CLAYTON, G., and A. ROBERTSON, 1955
Multiple range and multiple F tests. Biometrics 11: 1-42.
The efficacy of mutation breeding. Mutation and Plant Breeding. Natl. Acad. Sci.-Natl. Res. Counc. Pubn. 891 : 4 6 4 5 .
Genetic Homeostasis. Oliver and Boyd, Edinburgh,
Induced mutation of polygenes for quantitative characters in rice. J. Heredity 49: 11-14.
Effectiveness of artificial selection under irradiation of plateaued popu- lations of Drosophila melanogaster. pp. 42-66. Symp. on Genetics of Population Structure. Intern. Union Biol. Sci. Publ., Series B, No. 15. __ 1954b Artificial selection of a quantitative trait in Drosophila melanogaster under increased mutation rate. Proc. 9th Intern. Congr. Genet. (Caryologia 6 Suppl. Vol.) 861-864.