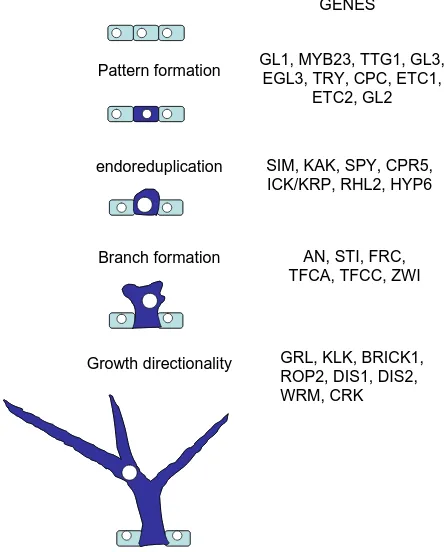
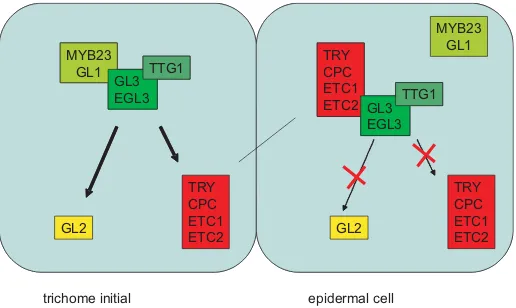
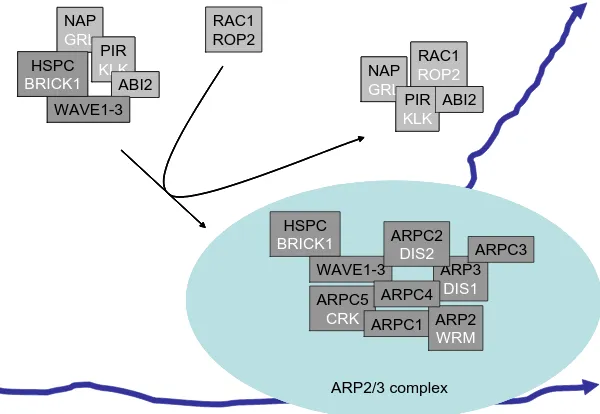
Epidermal differentiation: trichomes in Arabidopsis as a model system
Full text
Figure
Related documents
SSB (the Disaster Ready Schools) program is according to item 1, 3, and 5 of the Hyogo agreement which aims at providing protection to the school community when facing disasters
The remaining quantity from 5-Bromo-1-pentene (8 g, 53.68 mmol) was treated with 10 mL THF and it's added dropwise to the reaction mixture with continued stirring
Keywords: Antibacterial activity, synergy, Staphylococcus aureus , essential oil, Rosmarinus officinalis L., ciprofloxacin.. The essential oil is usually separated
Being a new imaging modality in our hospital, evaluation of the effectiveness of 64–row multislice CTA in detecting intracranial aneurysms in ruptured subarachnoid haemorrhage
Systematic analysis of this cytoplasmic signaling pathway determined that the ANDV N protein uniquely inhibits RIG-I-, MDA5-, MAVS-, and TBK1/IKK-directed ISRE, B, and
developed symptoms immediately after oral milk provocation gave a positive result on the RAST test. Although high serum total IgE was present in majority of the infants, it clearly
56. The court relied upon principles of federal common law to protect the FDIC from ordinary fraud claims of which it lacks knowledge. Consequently, the court held
(B) ELISA used to analyze titers of IgG against O-Ag (left) or porins (right) in serum samples from mice immunized twice with either STmGMMA O-Ag ⫹ (red circles) or O-Ag ⫺
