J. exp. Biol. 160, 1-23 (1991)
Printed in Great Britain © The Company of Biologists Limited 1991
THE EVOLUTION OF ACTIVITY CAPACITY
BY ALBERT F. BENNETT
Department of Ecology and Evolutionary Biology, University of California, Irvine, CA 92717, USA
Summary
The capacities of animals for activity (burst speed, maximal exertion, endur-ance) are examined in relation to their selective importance in extant populations and the pattern of their evolution in major animal taxa.
Activity capacities have been demonstrated to be both heritable and highly variable in natural populations and hence susceptible to natural selection. Some field studies have demonstrated significant positive associations between activity capacities, particularly burst speed, and survivorship; other studies have not. The potential for such selection therefore clearly exists, although it may not operate in all populations.
Comparative studies of major taxa have linked endurance capacities to maximal rates of oxygen consumption; speed and exertion are correlated with capacities for anaerobic metabolism, either the catabolism of phosphagens or the production of lactic acid or octopine, depending on taxon. In vertebrates, the primitive metabolic pattern involved the use of aerobic metabolism to support moderate swimming performance, supplemented by bursts of activity fuelled through lactic acid production. Because of much greater locomotor costs, the transition of vertebrates onto land entailed a decrease in endurance, which was greatly expanded again only after the evolution of the higher rates of aerobic metabolism characteristic of the birds and mammals. These greater aerobic capacities may have been selected for thermoregulatory reasons and/or for increased activity capacity itself.
Introduction
The animal kingdom consists of organisms of very diverse activity capacities, maxima] performance levels of which an animal is capable. Some animals are completely sessile, some are slow as snails, some are fleet but tire quickly, and some possess seemingly endless stamina. Physiologists, behaviourists, ecologists and natural historians have been interested in quantifying these activity capacities, because animal athletic abilities are of intrinsic interest and have functional and ecological implications. Knowing activity capacities may, for instance, permit a biologist to identify those structures and functions that impose intrinsic limitations
2 A. F . B E N N E T T
on performance, to define quantitatively the constraints within which any behaviour must occur (e.g. locomotor performance space, Bennett, 1989) or to predict the outcome of an encounter such as that between predator and prey. Most commonly examined are capacities for three different types of activity: burst speed, maximal exertion and endurance. Burst speed is the greatest velocity an animal attains over a short distance; maximal exertion is the work output during activity to rapid exhaustion, usually measured as distance travelled; and endur-ance is measured as the greatest speed an animal can sustain or the length of time an activity can be sustained. Different morphological, physiological and biochemi-cal processes are thought to underlie and limit each of these different types of behaviour (see Bennett, 1989). Research has concentrated on defining these limits; only now have investigations of the intermittent use of these capacities begun (e.g. R. B. Weinstein and R. J. Full, in preparation). Capacities are generally measured as absolute, instead of size-relative, speeds or distances because interactions among organisms in natural conditions depend on absolute and not relative performance.
How did the great diversity of activity types come about in different animals and how is it maintained? There has recently been considerable interest in and speculation on the evolution of activity capacity (e.g. Bennett and Ruben, 1979; Arnold, 1983; Bakker, 1983; Huey and Bennett, 1986; Pough, 1989; Bennett and Huey, 1991), and it is currently one of the most fertile areas of interactive research between the disciplines of evolutionary biology and comparative physiology/ physiological ecology. It is a logical bridge between these fields because scenarios of natural selection in which activity capacity is crucial for survival can be relatively easily envisioned. These include, for example, predator-prey, reproductive or territorial interactions in which minor differences in speed or exertion may mean success or failure, perhaps with fatal consequences. Such direct and decisive selection on other physiological variables, such as rates of maintenance metab-olism or sodium excretion, is less easy to envision. Interest in the evolution of activity capacity has ranged from investigations on its variability and importance in extant natural populations (microevolutionary studies) to its derivation in differ-ent major taxa, such as classes or phyla (macroevolutionary studies). As such, it touches on many fundamental issues in organismal biology and ecology. This essay examines some data and major issues that have been studied in regard to the evolution of activity capacities.
Microevolutionary studies
... 'the race is not to the swift nor the battle to the strong... but time and chance
happeneth to them all.' (Ecclesiastes 9:11)
The evolution of activity capacity 3
fother factors that selection operates only against individuals with greatly impaired capacities, e.g. birds that cannot fly or fish that cannot swim? Do time and chance, as suggested in Ecclesiastes, outweigh athletic ability? Only empirical studies on natural populations can answer these questions, and thereby inform speculation in both physiological ecology and evolutionary biology on the natural significance of activity capacity.
Measurement of natural selection
Two conditions must obtain for a trait to be potentially susceptible to natural selection (Endler, 1986): the trait must be heritable, that is, have a genetic basis so that offspring resemble parents in regard to the trait, and it must be variable within the population. Laboratory and pedigree studies have demonstrated that activity capacities have both these attributes. Performance capacities such as burst speed and endurance have been found to possess significant levels of heritability [broad-sense h2 (see Falconer, 1989)=0.2-0.7] in mammals (Ryan, 1975; Langlois, 1980; Bouchard and Malina, 1983a,b; Tolley etal. 1983; Gaffney and Cunningham, 1988), reptiles (van Berkum and Tsuji, 1987; Garland, 1988; Tsuji etal. 1989; Jayne and Bennett, 1990a,b; Garland et al. 1990a) and insects (Caldwell and Hegmann, 1969; Curtsinger and Laurie-Ahlberg, 1981). Considerable intrapopu-lation variability in activity capacity also exists, even at birth in precocial organisms. For example, coefficients of variation are 16, 54 and 66%, respect-ively, for burst speed, maximal exertion and endurance in newborn snakes
(Thamnophis sirtalis, Jayne and Bennett, 1990b). Even among adult individuals,
variability remains similarly great (e.g. Garland, 1984). Individual differences in these capacities persist through time, even after 1 year under field conditions (Huey and Dunham, 1987; van Berkum etal. 1989; Huey etal. 1990; Jayne and Bennett, 1990a), and may possess significant physiological or morphological correlates (e.g. Garland, 1984; Miles, 1987; Gleeson and Harrison, 1988; Kolok, 1990). Thus, activity capacities are apparently governed by genetic factors, persist through time and are highly variable among individuals, factors necessary for the potential for selection on the characters.
4 A. F . B E N N E T T
according to a variety of techniques (e.g. Lande and Arnold, 1983; Schluter, 1988;i Jayne and Bennett, 1990ft) that ask whether the survivors are a random subsample of the original group of animals released. If so, selection has not been detected; if they are not a random subsample, selection may be inferred to favour an extreme (directional selection) or the mean value (stabilizing selection) of the trait. As such factors as body size may induce false correlations and patterns within the data (see Bennett, 1987a), these are generally removed statistically before selection is analyzed.
An example of such a study is our work on a natural population of garter snakes
(Thamnophis sirtalis) in northern California (Jayne and Bennett, 1990a,b).
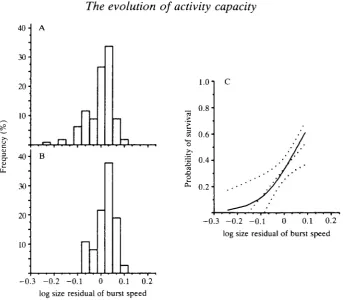
Reptiles are favoured model organisms for such studies because of high local population densities, low mobility and extreme precociality (Huey etal. 1983; Seigel etal. 1987). We measured burst speed, maximal exertion (as total distance crawled under pursuit) and endurance on 275 neonatal snakes born in our laboratory, as well as 382 field-caught snakes (including 86 neonates), and released all of them into their local population. The persistence of these individuals within the population was determined for periods as long as 3 years. When recaptured, an animal had its activity capacities measured again prior to release. As activity capacity in snakes depends greatly on body size (Heckrotte, 1967; Pough, 1978; Jayne and Bennett, 1990a), we analyzed size-corrected residuals. Activity capaci-ties were significantly correlated with year-to-year survival in some age classes of this population (Table 1). During the first year of life, body size (length, not mass) was a significant correlate of survival (P=0.022), but none of the activity capacity traits were significantly involved. However, for yearling animals, burst speed [both size-corrected and absolute (P<0.01)] and maximal exertion were positively and significantly associated with survivorship, and endurance was nearly so. In animals 2 years of age and older, burst speed, both in absolute and in size-corrected terms, was also significantly correlated with survivorship (illustrated in Fig. 1). The intensity of selection on these activity capacities is similar to that previously determined for morphological characters under selection (Schluter, 1988). Since
Table 1. Selection on activity capacities in different age classes in a natural
population of garter snakes (data from randomization tests for 1986-1987 survival from Jayne and Bennett, 1990b)
Size-corrected trait
Burst speed Maximal exertion Endurance
Neonatal
0.262 0.463 0.265
Age class
Yearling
0.007 0.008 0.060
Older
0.001 0.084 0.100
The evolution of activity capacity
40-
30-
20-
10-
40-
30-
20- 10-A
B
-—.
- ,
n
1.0
- 0.8
CO >
'E
3 0.6
0.2
-0.3 -0.2 -0.1 0 0.1 0.2 log size residual of burst speed
[image:5.451.56.400.51.351.2]-0.3 -0.2 -0.1 0 0.1 0.2 log size residual of burst speed
Fig. 1. Survival of garter snakes 2 years of age or older and burst speed capacity. (A) corrected distribution of speeds of snakes released in 1986 (/V=113). (B) Size-corrected speeds (measured in 1986) of those snakes recaptured in 1987 (N=37). (C) Probability of survival as a function of size-corrected speed. The dotted lines are the linear regression and 95 % confidence limits given by Lande and Arnold (1983); the solid line is the cubic spline fitness function (Schluter, 1988). (From Jayne and Bennett, 1990b.)
snakes in this population do not reproduce until they are 3-4 years old, some activity capacities are associated with differential survivorship prior to repro-duction and may therefore have an impact on fitness.
6 A. F. BENNETT
McPhail, 1985) (these individuals were also larger and this result may be complicated by size effects). Faster fence lizards were found to be socially dominant in behavioural interactions with size-matched conspecifics (Garland
et al. 1990b). Comparative studies on taxa from different natural thermal regimes
show shifts in the thermal dependence of activity capacities consistent with the notion of adaptive evolution in response to selection on those capacities (endurance in populations of killifish, Fundulus heteroclitus, DiMichele and Powers, 1982; burst speed in species of scincid lizards, Huey and Bennett, 1987). Several studies, therefore, have found associations between activity capacities and survivorship, important behaviours or temperature adaptation. These results point to the significance of these capacities in evolution. However, the effects are clearly not universal, and selection for these factors may be only episodic, too difficult to detect or simply absent in some natural populations.
Identifying the actual agents of selection on activity capacity through focal animal observations has been suggested as an alternative approach to microevolu-tionary studies (Pough, 1989). This, however, to my mind is a secondary question, to be approached only after the likelihood of selection has been established with the type of experiment outlined above. If selection effects are undetectable in those observations, it is unlikely to be profitable to search for agents. Identifi-cation of such agents is highly problematic, and a series of studies on different species of anuran amphibians has failed to produce correlations between activity performance and characters thought to be related to fitness (Wells and Taigen, 1984; Sullivan and Walsberg, 1985; Walton, 1988a). Studies on small numbers of individuals may, however, fail to detect selection even if it is occurring, because of low statistical power and the stochastic nature of the selection event, or may miss the event if it occurs at a different age from that studied. Ideal studies of this type would require continuous observation of very large numbers of animals in the field throughout their entire lifetime, or at least until they are postreproductive. Such studies are generally impossible.
Macroevolutionary patterns
Considerable interest has been directed towards the evolution of activity capacity and its metabolic and physiological support among and within different animal taxa. As is not uncommon in evolutionary studies, such macroevolutionary considerations do not flow seamlessly from microevolutionary observations and speculation, so these areas of thought regarding activity capacity have largely developed independently of each other. Macroevolutionary studies have been directed primarily towards the analysis of qualitative differences in metabolic pathways among different animal phyla and quantitative differences in metabolic capacities within phyla. These differences determine rates of ATP generation and hence ability to support the skeletal muscle contraction on which activity depends.
Metabolic support of activity
The evolution of activity capacity 7
iu moderate levels of activity. As activity level (e.g. speed) increases, oxygen consumption rises to meet increased demand for ATP production. The quantitat-ive pattern of this increase depends primarily on locomotor mode (e.g. walk-ing-running, swimming) (Schmidt-Nielsen, 1972; Tucker, 1975) and not on taxon. In running animals, for instance, the patterns of speed dependence of oxygen consumption and size-adjusted net cost of transport are very similar in insects (Full
etal. 1990), crustaceans (Herreid, 1981), reptiles (Gleeson, 1979; Walton et al.
1990), mammals and birds (Taylor et al. 1982). Maximal oxygen consumption sets an upper limit on this kind of sustainable behaviour. Endurance and maximal oxygen consumption are well correlated both within and among taxa according to differences in body size, athletic ability and body temperature (Brett, 1972; Taylor, 1982; Bennett and John-Alder, 1984; Garland, 1984; Taylor etal. 1987; Full et al. 1988). The evolution of endurance capacity is therefore intimately linked with, and depends on, evolutionary changes in maximal capacities to consume oxygen.
Activity requiring power output in excess of that supported by oxygen consumption is fuelled by anaerobic metabolism. Part of this may be provided by endogenous stores of ATP or muscle phosphagens (creatine or arginine phos-phate), but these are present in small quantities in comparison to metabolic demand. Additional ATP generation is provided by catabolic anaerobic pathways. A considerable diversity of such pathways is known in the animal kingdom (Hochachka, 1980; Hochachka and Somero, 1984; Gade and Grieshaber, 1986), with a correspondingly diverse array of end products, energetic efficiencies and rates of energy production. In a highly interesting article, Gnaiger (1983) has argued on thermodynamic grounds (challenged by Watt, 1986) that rate of power output and efficiency of energy transformation cannot be optimized simul-taneously within any one of these pathways. Hence, some anaerobic pathways, those terminating in lactic acid, alanine or octopine, emphasize high rates of ATP formation and relatively low ATP yield per unit of substrate catabolized. Others, those terminating in succinate or propionate, have a greater energetic efficiency of substrate use but a low capacity for power output. The former class of pathways is therefore used in fuelling short-term bursts of activity (when ATP demand exceeds aerobic ATP supply) and can be useful in enhancing activity capacity. The latter pathways are used for enduring environmental hypoxia and permit long-term survival in anoxia but do not contribute to activity enhancement. Lactic acid production is used as the principal anaerobic pathway during activity in ver-tebrates (Bennett, 1978), urochordates and echinoderms (Ruben and Parrish, 1991), crustaceans (McMahon, 1981), arachnids (Prestwich. 1988). leeches (Zebe
et al. 1981) and some insects (Harrison et al. 1989; activity metabolism in flying
8 A. F. BENNETT
compounds during activity is still unknown, both because of uncertainties in tlW phylogeny of the invertebrate groups and because of lack of information on end product accumulation in many of them. (The literature on anaerobic product formation in invertebrates is reviewed in Gade and Grieshaber, 1986.) The evolution of the capacity for bouts of intense activity therefore generally depends on the development of capacities for rapid formation and tolerance of high concentrations of lactic acid or octopine. Production of the former is associated with rapid exhaustion, the exact physiological causation of which is still in dispute.
Evolution of activity capacities in the vertebrates
The evolution of activity capacities and their metabolic bases in the vertebrates has engendered considerable interest and speculation. The scenarios proposed will form the basis for the ensuing discussion of the macroevolutionary development of activity capacity within a major taxon. This concentration of interest is not simply a reflection of vertebrate self-absorption. It is due primarily to an excellently documented palaeontological history, a great diversity of types and mechanisms of locomotion exhibited by members of the group, and a very large behavioural and physiological data base.
Ancestral condition
The evolution of activity capacity
Chordates
I
Vertebrates
I
o
£
'OXl
-c
•g
st
r
* in tC
c *
cd
>-»
4)
a. E
en
I s
[image:9.451.135.323.69.296.2]— 13
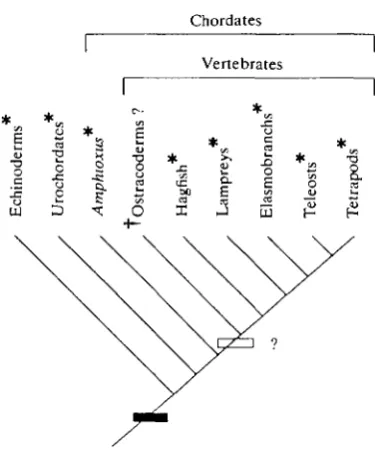
Fig. 2. Phylogenetic distribution in the deuterostome lineage of lactic acid formation during intense activity. At question is whether ancestral vertebrates (marked by the open box) and later ostracoderms (now extinct, marked by a dagger) possessed this character. Extant groups in which lactic acid formation during activity has been demonstrated are marked with asterisks. The most parsimonious interpretation is the presence of the character in ancestral vertebrates and ostracoderms and early evolution of the character within the lineage (marked by black box). References to lactate formation are given in the text; phylogeny from Maisey (1986).
appears to be an inheritance from deuterostome ancestors and not a novel invention of the group.
Aerobic scopes of modern fish are relatively low in comparison with those of endothermic vertebrates or flying insects (Beamish, 1978; Bennett, 1978; Taylor
et al. 1981; Bartholomew, 1981). However, fish have a considerable range of
sustainable speeds supported by these aerobic capacities and, consequently, relatively great endurance. This endurance capacity is due to the low cost of swimming as a locomotor mode (Fig. 3): net costs of transport (mlO2
10
A . F. BENNETTa. E
[image:10.451.107.350.51.250.2]
2-2 4 6 Speed (km h"1)
Fig. 3. Oxygen consumption as a function of speed in a fish (1.43 kg salmon,
Oncorhynchus nerka, 15°C, Brett, 1965), lizard (0.87kg tegu, Tupinambis nigropunc-tatus, 35 °C, Bennett and John-Alder, 1984) and mammal (1.0 kg, slope and intercept
from Taylor et al. 1970; maximal aerobic speed from Garland et al. 1988). Arrows indicate the greatest aerobically sustainable speed for each animal.
Table 2. Endurance (sustainable speed) and burst speed for animals from different
groups of vertebrates
Group
Endurance (rnrnin"1)
Burst speed
(mmin"1) Ratio Fishes
Lizards Mammals
10-15 30-40 37-40
42 13 88
108 240 239
2.6
18.5
2.7
Tb, body temperature.
Predicted values for 100 g animals in each group are derived from general allometric equations for mammals (Garland, 1983; Garland et al. 1988), lizards (T. Garland, personal communication) and fish (calculated from Tables 4 and 6 in Beamish, 1978).
Transition to the land
[image:10.451.42.416.369.436.2]The evolution of activity capacity 11
, decreased in comparison with values measured in fish. Sustainable walking speeds of salamanders are less than Smmin"1 (Full, 1986; Feder, 1986; Else and Bennett, 1987; Full et al. 1988), less than one-tenth those of fish swimming (Table 2). Sustainable speeds of reptiles are greater (Table 2), but still far below those of fish (Fig. 3). This decrement in endurance is the result of greatly increased locomotor costs in the terrestrial environment. Net costs of transport are nearly 10 times greater in terrestrial walkers and runners than in swimmers of equal body size (Schmidt-Nielsen, 1972; Tucker, 1975). The biomechanics underlying this difference are still not fully understood. It may relate to the costs of sequential acceleration and deceleration of limbs and vertical displacement of centre of mass (Alexander, 1977), but the equality of locomotor costs of snakes and limbed reptiles (Walton et al. 1990) suggests that such explanations are less than complete, as snake locomotion has neither of these features. Factors involving the rate of cycling of skeletal muscle contraction may be of greater importance (Full et al. 1990; Kram and Taylor, 1990). In any event, greatly increased locomotor costs coupled with a lack of increment in aerobic capacity meant that limits of endurance were reached at much lower speeds and levels of exertion than in their aquatic ancestors (Fig. 3; Bennett, 1985).
How was capacity for burst speed affected by the transition to land? Data on burst speed capacities of amphibians are scant: tiger salamanders (Ambystoma
tigrinum) attain peak running speeds of only about lOmmin"1 (Else and Bennett, 1987; Shaffer et al. 1991), slow in comparison to fish (Table 2). Burst locomotor performance of anuran amphibians is generally measured as jump distance (e.g. Zug, 1978), which is not directly comparable to other measurements reported here. Lizards are capable of running at 10-30 times sustainable speeds for short bursts (Table 2; Bennett, 1982; Garland, 1982, personal communication), speeds that are substantially greater than those of fish. These bursts of activity are fuelled by ATP or creatine phosphate catabolism and anaerobic glycolysis and lactic acid formation, the capacity for which is very great in lizards (Bennett and Licht, 1972) and some amphibians (Bennett and Licht, 1974). In comparison to fish, a much greater portion of the total behavioural repertoire of these amphibians and reptiles is thus accessed only by anaerobic metabolism (Table 2). Activity capacities of these terrestrial tetrapods are consequently short on endurance, but their maximal exertion and burst speed can be very great. Their behaviour is typically limited to rather slow movements, punctuated with periods of very intense bouts of activity of short duration.
Evolution of higher metabolic rates in birds and mammals
This pattern of activity capacities of the terrestrial ectothermic vertebrates was fundamentally altered during the evolution of the mammals and birds. Endurance increased substantially in these lineages. Maximal sustainable speeds of mammals are 6-7 times greater than those of lizards of equal size and body temperature
12 A . F . B E N N E T T
reptiles (Bennett and Ruben, 1979). Burst speed, however, has apparently been greatly modified: maximal speeds of lizards and mammals of equal size are not significantly different (Table 2; Garland, 1982, personal communication).
What is responsible for these differences in activity capacity? Most approaches to this question have restricted themselves to comparisons of mammals and reptiles, usually lizards. Such comparisons are made because of their similarity of body form and relative ease of measurement (in comparison to birds) of functional capacities during activity. These approaches implicitly assume that metabolic and performance traits that are widely shared among extant reptilian taxa are representative of the ancestral condition from which mammals evolved. Even though endothermy was evolved completely independently in the two lines, mammalian and avian systems appear highly convergent in many of their metabolic aspects (compare allometric equations for many such variables in Calder, 1984). Mammals may, therefore, potentially serve as adequate models in such comparisons for both endothermic taxa. In mammals and birds, both basal and maximal levels of oxygen consumption and aerobic scopes are approximately 6-10 times greater than those of extant reptiles of the same body mass and temperature (Bartholomew and Tucker, 1963; Brett, 1972; Bennett, 1978, 1982; Taylor et al. 1981). Net costs of walking and running are equal in mammals and reptiles (Bakker, 1972; Gleeson, 1979; Bennett and John-Alder, 1984), so the greater aerobic scope of the former affords a much greater range of sustainable speeds that can be maintained without supplementary anaerobic metabolism (Table 2, Fig. 3). Capacities for anaerobic metabolism, both anaerobic scope and anaerobic capacity (Bennett and Licht, 1972), are rather similar in mammals and reptiles (Bennett, 1978, 1982; Ruben and Battalia, 1979), as are activities of glycolytic enzymes (Bennett, 1972a) and myofibrillar ATPase (Gleeson et al. 19806) in saurian and mammalian muscle. Hence, the lack of divergence in burst speed and maximal exertion capacities of the two groups is not particularly surprising.
The evolution of activity capacity 13
mitochondrial volume and surface area. Taylor and Weibel (1981) have argued that all portions of the oxygen transporting and utilizing apparatus should have similar maximal flux capacities, that is that there should be no particular limiting element in the chain and that no part should possess excess transport capacity. If such parity were maintained, the augmentation of all the elements of oxygen transport would have had to have evolved in concert and simultaneously. Given their diverse nature and probable diverse genetic control, this synchroniz-ation seems unlikely. Withers and Hillman (1988) have inferred a cardiovascular limitation on maximal oxygen consumption in anuran amphibians; a similar restriction in this or another element of the system may possibly have occurred in primitive reptiles. Probably the evolution of increased aerobic capacities in birds and mammals involved the sequential improvement in whatever aspect of the system formed its most limiting element at any particular time.
Where in the mammalian and avian lineages did high metabolic rates and endothermy appear? Are they inventions within the classes Mammalia and Aves or did they develop in groups ancestral to these? Since most characters relating to oxygen consumption and thermoregulation do not fossilize, there has been much heat but little light generated from very indirect evidence about the metabolic condition of mammal-like reptiles and dinosaurs. Bennett and Ruben (1986) proposed the following criteria for evaluating a character presumably associated with endothermy in extinct organisms: it must not be present in any extant ectotherm and it must have a clear and direct association with high metabolic rate. Further, the presence of such characters in sister taxa suggests ancestral inheri-tance. Reviewing the evidence, Bennett and Ruben (1986) concluded that the therapsid ancestors of mammals probably possessed high aerobic capacities and may have been well on the path to endothermy prior to the evolution of mammals
per se. This conclusion was based on two lines of evidence. First, cynodont
therapsids possessed elaborate turbinal bones in the nasal cavity, indicative of the conditioning of extensive volumes of ventilated air. These are not present in any ectothermic vertebrate and are directly associated with capacities for high levels of oxygen uptake. Second, all physiological and morphological features related to high levels of oxygen consumption and endothermy are shared by the mammalian sister taxa Prototheria (monotremes) and Theria (marsupials and placentals). These groups diverged fairly early in mammalian evolution and remain distinct in many features of their reproduction, endocrinology and neurology. The pos-session of so many diverse systems related to high levels of oxygen consumption and endothermy (e.g. diaphragm, four-chambered heart, hair) is due either to unprecedented levels of convergence or ancestral inheritance.
The presence or absence of endothermy in dinosaurs has been a topic of considerable discussion and sometimes acrimonious debate over the past two decades (see references in Thomas and Olson, 1980; Czerkas, 1986; Bakker, 1986). Both proponents and opponents have elaborate arguments and
14 A . F. B E N N E T T
aforementioned criteria of absence in all extant ectotherms and a clear and association with high metabolic rates. Further, as the birds are a monophyletic group, no extant endothermic sister taxon exists from which ancestral inheritance of endothermy can be inferred. I therefore personally consider the contention of dinosaur endothermy unproved.
Selective factors in the evolution of higher metabolic rate
What occasioned the evolution of increased aerobic capacity in the mammalian and avian lineages? Given the difficulty in determining selective factors on morphology, physiology and performance in extant populations (see microevol-utionary section above), it is probably foolhardy to attempt it for unknown organisms now extinct that lived in poorly understood environments. Two factors, however, have been suggested as major selective advantages. The first is metabolic thermoregulation. Body temperatures of birds and mammals are both higher and more constant than those of most ectothermic vertebrates. High body tempera-tures are suggested to have been favoured to maximize catalytic capacity of thermally dependent rate processes (Heinrich, 1977; Hochachka and Somero, 1984; Bennett, 1987ft). Constancy of temperature presumably also favours stability of those processes and homeostasis in general, although these purported advantages are more nebulous and the logic behind them often circular. The evolution of endothermic homeothermy in these lineages required the elevation of maintenance rates of aerobic metabolism. Placing an insulatory layer on a vertebrate ectotherm does not confer significant homeothermy, as Cowles (1958) demonstrated directly by outfitting lizards with mink coats. Increased insulation, above that afforded by the reptilian integument, is necessary for homeothermy, at least in small endotherms, but is in itself not sufficient to produce it. Increments in maintenance metabolic rate occurred independently in both avian and mammalian lineages, converging on very similar levels in the two groups, when differences in body temperature are taken into account (equations in Peters, 1983; Calder, 1984). It is important to note that this metabolic increment is fundamentally different from that in other 'endothermic' animals, which elevate heat production by skeletal muscle contraction (e.g. insects, Kammer, 1981; tunas, Stevens and Neill, 1978; brooding pythons, Hutchison et al. 1966) or modified skeletal muscle tissue (e.g. billfish heater muscle, Block, 1987). The high rates of maintenance metabolism of birds and mammals are not myogenically based; they are generated primarily by visceral organs and the central nervous system. In humans, nearly 70% of resting metabolic rate is accounted for by the metabolism of the heart, kidneys, brain, liver and intestines, even though they constitute only 7 % of body mass (Aschoff et al. 1971). Skeletal muscle tissue can produce high levels of heat in the cold or during activity, but is largely uninvolved in the production of the high basal metabolic rate of birds and mammals.
The evolution of activity capacity 15
pBmeothermy and high body temperatures can be achieved. Basal metabolic rates of birds and mammals exceed those of extant reptiles of equal size and body temperature by a factor of 6-10, and this differential is substantially greater at lower ambient temperatures (Dawson and Bartholomew, 1956; Bennett and Dawson, 1976). Minor increments (e.g. a doubling) in maintenance metabolic rates would not have been effective in establishing homeothermy or raising body temperature, but would have added substantially to energetic demands of field-active animals with little apparent benefit. The problem for the thermoregulatory explanation of the evolution of high metabolic rate lies in justifying intermediate stages in selective terms, presumptive adaptive valleys between adaptive peaks. Even if the final goal of endothermic homeothermy is worthwhile in terms of fitness, the condition cannot evolve unless all intermediate stages also confer sequentially greater fitness benefits. In a character as complex as endothermic temperature regulation, involving so many physiological systems, it is highly doubtful that it could have developed in its entirety without many intermediate stages. The problem becomes even more complex if ancestral animals had the capacity for behavioural thermoregulation of high body temperatures, as do many extant reptiles. In this case the benefits of high and relatively constant temperature would already have been available without increased metabolic cost. One argument has been advanced (Crompton et al. 1978; Taylor, 1980) that endo-thermy evolved in the mammalian line in response to nocturnality, when behavioural thermoregulation at high temperature is precluded. This argument loses force, however, if endothermy was already present in the reptilian ancestors of these nocturnal mammals (see above, Bennett and Ruben, 1986).
A second explanation suggested for the evolution of increased metabolic rates in the mammalian and avian lineages is the increased capacity for aerobically supported activity (Regal, 1978; Bennett and Ruben, 1979). Increased levels of maximal oxygen consumption directly expand the range of levels of sustainable exertion and the behaviours that depend on them (e.g. Fig. 3). Animals with greater aerobic limits have greater endurance in a variety of different activities that may influence fitness, such as prey pursuit, predator avoidance, territory maintenance and courtship. It is relatively easy to visualize a direct benefit in selective terms to such increased endurance capacity, particularly in animals, such as reptiles, that have limited aerobic capabilities and must otherwise depend heavily on anaerobic metabolism, with its ensuing exhaustion, to support vigorous activity. Ancestral animals in both the mammalian and avian lineages seem to have been progressively more capable of vigorous activity, as shown by general skeletal rearrangements, such as a change in limb suspension and reduction of skeletal mass (Romer, 1966; Olson, 1971; Kemp, 1982; Carroll, 1988). The expansion of aerobic capacities to support endurance-related behaviours is, thus, plausible in its progressive accrual of benefits during its evolution (i.e. no apparent adaptive valleys) and is also in accord with general palaeobiological patterns of the groups i question.
16 A. F. BENNETT
endothermy is explaining the concomitant rise in maintenance metabolic rate w i ^ that of maximal oxygen consumption (Bennett and Ruben, 1979). If increased aerobic capacity during activity were the only consideration, it seems that a more satisfactory solution would have been to expand maximal aerobic rates without increasing maintenance rates. Thereby, the animals could benefit from both increased endurance and low maintenance costs, the latter being characteristic of ectothermy (Bennett and Dawson, 1976; Pough, 1983). A general linkage between maximal and maintenance aerobic metabolic rates was proposed to explain increments in the latter as the former is increased (Bennett and Ruben, 1979). In all groups of vertebrates, even within individual poikilotherms at different body temperatures, maximal oxygen consumption exceeds maintenance by an average factor of approximately 10. The physiological basis of this linkage is unclear, especially as maximal rates are largely myogenically based and maintenance rates are not. This factorial difference is not fixed or absolute, and comparisons among disparate species of similar body size can show differences in maximal rates without similar differences in maintenance rates (e.g. Bennett, 1972b, 1978; Ruben, 1976; Taylor et al. 1987). In phylogenetically controlled comparisons, which are inherently more reliable (Felsenstein, 1985; Huey, 1987), significant positive correlations between these factors occur among groups of anuran amphibians (Taigen, 1983; Walton, 1988£>). Such correlations have not been found within species (Pough and Andrews, 1984; Garland and Else, 1987; Garland et al. 1987). Thus, this association remains a generality, without a firm mechanistic explanation or intraspecific correlational basis. Two factors, however, bear further investigation. First, all previous studies refer to phenotypic correlations among metabolic rates. Genetic correlations may have very different patterns and link traits evolutionarily without obvious phenotypic associations (Arnold, 1987). Examinations of the genetic relationships between maximal and maintenance levels of oxygen consumption would prove helpful in understanding their association. Second, the relationship between intraspecific and interspecific correlation and its relationship to the analysis of evolutionary pattern is just beginning to be analyzed (Emerson and Arnold, 1989). Further theoretical and experimental studies on these general associations would also be valuable.
In summary, both explanations for the evolution of high metabolic rates associated with endothermy have positive features and difficulties. Perhaps a combination of thermoregulatory and activity capacity factors were involved simultaneously or sequentially. The early stages of its evolution may have been dominated by selection for increased endurance, and its later stages by selection for homeothermy. Unifactorial explanations for the evolution of any complex character are almost certainly incomplete.
Future directions
The evolution of activity capacity 17
ppganismal biology. In the studies cited above, an excellent beginning has been made, both conceptually and experimentally, in addressing the issues. In my opinion, we would benefit from further studies on the operation of selection on these characters in natural populations of diverse taxa, followed by studies attempting to quantify locomotor behaviour under field conditions and to identify selective agents. The bridge between microevolutionary studies on extant popu-lations and diversification of capacities among higher taxa is a difficult one to build, but one well worth the attempt. Comparative studies on different populations within a species or among congeneric species with diverse activity capacities may be useful in uncovering the physiological and morphological bases, and perhaps the ecological determinants, of activity capacity. Experimental manipulations, where feasible, of such factors as food or mate availability or predation may be particularly useful in examining their influence as agents of selection. Choice of appropriate experimental groups will obviously be crucial to the success of these studies. Analysis of ancient historical pattern, for instance the selective causation and mechanism of the evolution of endothermy, will probably always remain speculative. However, the use of phylogenetically based analyses and the future availability of new analytical techniques may help to resolve some of the major questions in this area.
I thank R. J. Full, R. B. Huey and J. A. Ruben for critically reading and commenting on the manuscript and S. M. Reilly for help preparing the figures. Supported by NSF Grants DCB88-20128 and BSR90-18054.
References
ALEXANDER, R. MCN. (1977). Terrestrial locomotion. In Mechanics and Energetics of Animal
Locomotion (ed. R. McN. Alexander and G. Goldspink), pp. 168-203. London: Chapman
and Hall.
ARNOLD, S. J. (1983). Morphology, performance and fitness. Am. Zool. 23, 347-361.
ARNOLD, S. J. (1987). Genetic correlation and the evolution of physiology. In New Directions in
Ecological Physiology (ed. M. E. Feder, A. F. Bennett, W. Burggren and R. B. Huey), pp.
189-215. Cambridge, England: Cambridge University Press.
ASCHOFF, J., GUNTHER, B. AND KRAMER, K. (1971). Energiehaushalt und Temperaturregulation. Munich: Urban and Schwarzenberg.
BAKKER, R. T. (1972). Locomotor energetics of lizards and mammals compared. Physiologist 15, 76.
BAKKER, R. T. (1983). The deer flees, the wolf pursues: incongruencies in predator-prey coevolution. In Coevolution (ed. D. J. Futuyma and M. Slatkin), pp. 350-382. Sunderland, MA: Sinauer.
BARKER, R. T. (1986). The Dinosaur Heresies. New York: Win. Morrow and Co.
BARTHOLOMEW, G. A. (1981). A matter of size: an examination of endothermy in insects and terrestrial vertebrates. In Insect Thermoregulation (ed. B. Heinrich), pp. 45-78. New York: Wiley-Interscience Pub.
BARTHOLOMEW, G. A. AND TUCKER, V. A. (1963). Control of changes in body temperature, metabolism, and circulation by the agamid lizard, Amphibolurus barbatus. Physiol. Zool. 36, 199-218.
18 A. F. BENNETT
BENNETT, A. F. (1972a). A comparison of activities of metabolic enzymes in lizards and r a ^
Comp. Biochem. Physiol. 42B, 637-647.
BENNETT, A. F. (1972i>). The effect of activity on oxygen consumption, oxygen debt, and heart
rate in the lizards Varanus gouldii and Sauromalus hispidus. J. comp. Physiol. 79, 259-280.
BENNETT, A. F. (1973). Blood physiology and oxygen transport during activity in two lizards,
Varanus gouldii and Sauromalus hispidus. Comp. Biochem. Physiol. 46A, 673-690. BENNETT, A. F. (1978). Activity metabolism of the lower vertebrates. A. Rev. Physiol. 40,
447-469.
BENNETT, A. F. (1982). The energetics of reptilian activity. In Biology of the Reptilia, vol. 13 (ed. C. Gans and F. H. Pough), pp. 155-199. New York: Academic Press.
BENNETT, A. F. (1985). Energetics of locomotion. In Functional Vertebrate Morphology (ed. M. Hildebrand, D. M. Bramble, K. F. Liem and D. B. Wake), pp. 173-184. Cambridge, MA: Harvard University Press.
BENNETT, A. F. (1987a). Interindividual variability: an underutilized resource. In New
Directions in Ecological Physiology (ed. M. E. Feder, A. F. Bennett, R. B. Huey and W.
Burggren), pp. 147-169. Cambridge, England: Cambridge University Press.
BENNETT, A. F. (1987ft). Evolution of the control of body temperature: is warmer better? In
Comparative Physiology: Life in Water and on Land (ed. P. Dejours, L. Bolis, C. R. Taylor
and E. R. Weibel), pp. 421-431. Padova, Italy: Liviana Press.
BENNETT, A. F. (1989). Integrated studies of locomotor performance. In Complex Organismal
Functions: Integration and Evolution in Vertebrates (ed. D. B. Wake and G. Roth), pp.
191-202. Dahlem Konferenzen. Chichester, England: John Wiley and Sons.
BENNETT, A. F. AND DAWSON, W. R. (1976). Metabolism. In Biology of the Reptilia, vol. 5 (ed. C. Gans and W. R. Dawson), pp. 127-223. New York: Academic Press.
BENNETT, A. F. AND HUEY, R. B. (1991). Studying the evolution of physiological performance. In Oxford Surveys of Evolutionary Biology, vol. 7 (ed. D. Futuyma and J. Antonovics). New York: Oxford University Press (in press).
BENNETT, A. F. AND JOHN-AIDER, H. B. (1984). The effect of body temperature on the locomotory energetics of lizards. J. comp. Physiol. 155, 21-27.
BENNETT, A. F. AND LICHT, P. (1972). Anaerobic metabolism during activity in lizards. /. comp. Physiol. 81, 277-288.
BENNETT, A. F. AND LICHT, P. (1974). Anaerobic metabolism during activity in amphibians.
Comp. Biochem. Physiol. 48A, 319-327.
BENNETT, A. F. AND RUBEN, J. A. (1979). Endothermy and activity in vertebrates. Science 206, 649-654.
BENNETT, A. F. AND RUBEN, J. A. (1986). The metabolic and thermoregulatory status of therapsids. In The Ecology and Biology of Mammal-like Reptiles (ed. N. Hotton, P. D. MacLean, J. J. Roth and E. C. Roth), pp. 207-218. Washington, DC: Smithsonian Institution.
BLOCK, B. A. (1987). Billfish brain and eye heater: a new look at nonshivering heat production.
News physiol. Sci. 2, 208-213.
BOUCHARD, C. AND MALINA, R. M. (1983a). Genetics for the sport scientist: selected methodological considerations. Exercise Sport Sci. Rev. 11, 274-305.
BOUCHARD, C. AND MALINA, R. M. (1983ft). Genetics of physiological fitness and motor
performance. Exercise Sport Sci. Rev. 11, 306-339.
BRETT, J. R. (1965). The relation of size to rate of oxygen consumption and sustained swimming speed of sockeye salmon (Oncorhynchus nerka). J. Fish. Res. Bd Can. 22, 1491-1501.
BRETT, J. R. (1972). The metabolic demand for oxygen in fish, particularly salmonids, and a
comparison with other vertebrates. Respir. Physiol. 14, 151-170.
CALDER, W. A., LTJ (1984). Size, Function, and Life History. Cambridge, MA: Harvard University Press.
CALDWELL, R. L. AND HEGMANN, J. P. (1969). Heritability of flight duration in the milkweed bug
Lygaeus kalmii. Nature 223, 91-92.
CARRIER, D. (1987). The evolution of locomotor stamina in tetrapods: circumventing a
mechanical constraint. Paleobiology 13, 326-341.
CARROLL, R. L. (1988). Vertebrate Paleontology and Evolution. New York: W. H. Freeman aqfl
The evolution of activity capacity 19
•IWLES, R. B. (1958). Possible origin of dermal temperature regulation. Evolution 12,347-357.
CROMPTON, A. W., TAYLOR, C. R. AND JAGGER, J. A. (1978). Evolution of homeothermy in mammals. Nature 111, 333-336.
CURTSINGER, J. W. AND LAURIE-AHLBERG, C. C. (1981). Genetic variability of flight metabolism in Drosophila melanogaster. I. Characterization of power output during tethered flight.
Genetics 98, 549-564.
CZERKAS, S. (ed.) (1986). Dinosaurs Past and Present. Los Angeles: Los Angeles County Museum Press.
DAWSON, W. R. AND BARTHOLOMEW, G. A. (1956). Relation of oxygen consumption to body weight, temperature, and temperature acclimation in lizards Uta stansburiana and Sceloporus
occidentals. Physiol. Zool. 29, 40-51.
DEJOURS, P. (1981). Principles of Comparative Respiratory Physiology, 2nd edn. Amsterdam: North Holland.
DIMICHELE, L. AND POWERS, D. A. (1982). Physiological basis for swimming endurance differences between LDH-B genotypes of Fundulus heteroclitus. Science 216, 1014-1016.
ELSE, P. L. AND BENNETT, A. F. (1987). The thermal dependence of locomotor performance and muscle contractile function in the salamander Ambystoma tigrinum nebulosum. J. exp. Biol. 128, 219-233.
ELSE, P. L. AND HULBERT, A. J. (1981). Comparison of the 'mammal machine' and the 'reptile
machine': energy production. Am. J. Physiol. 240, R3-R9.
EMERSON, S. B. AND ARNOLD, S. J. (1989). Intra- and interspecific relationships between morphology, performance, and fitness. In Complex Organismal Functions: Integration and
Evolution in Vertebrates (ed. D. B. Wake and G. Roth), pp. 295-314. Dahlem Konferenzen.
Chichester, England: John Wiley and Sons.
ENDLER, J. A. (1986). Natural Selection in the Wild. Princeton, NJ: Princeton University Press. FALCONER, D. S. (1989). Introduction to Quantitative Genetics, 3rd edn. London: Longman.
FEDER, M. E. (1986). Effect of thermal acclimation on locomotor energetics and locomotor performance in a lungless salamander, Desmognathus ochrophaeus. J. exp. Biol. 121, 271-283.
FELSENSTEIN, J. (1985). Phylogenies and the comparative method. Am. Natur. 125, 1-15. FULL, R. J. (1986). Locomotion without lungs: energetics and performance of a lungless
salamander. Am. J. Physiol. 251, R775-R780.
FULL, R. J., ANDERSON, B. D., FINNERTY, C. M. AND FEDER, M. E. (1988). Exercising with and without lungs. I. The effects of metabolic cost, maximal oxygen transport and body size on terrestrial locomotion in salamander species. J. exp. Biol. 138, 471-485.
FULL, R. J., ZUCCARELLO, D. A. AND TULLIS, A. (1990). Effect of variation in form on the cost of terrestrial locomotion. /. exp. Biol. 150, 233-246.
GADE, G. (1981). Energy production during swimming in the adductor muscle of the bivalve
Lima hians: comparison with the data from other bivalve molluscs. Physiol. Zool. 54,
400-406.
GADE, G. AND GRIESHABER, M. K. (1986). Pyruvate reductases catalyze the formation of lactate and opines in anaerobic invertebrates. Comp. Biochem. Physiol. 83B, 255-272.
GAFFNEY, B. AND CUNNINGHAM, E. P. (1988). Estimation of genetic trend in racing performance of thoroughbred horses. Nature 332, 722-724.
GARLAND, T., JR (1982). Scaling maximal running speed and maximal aerobic speed to body mass in mammals and lizards. Physiologist 25, 338.
GARLAND, T., JR (1983). The relation between maximal running speed and body mass in terrestrial mammals. /. Zool., Lond. 199, 157-170.
GARLAND, T., JR (1984). Physiological correlates of locomotory performance in a lizard: an allometric approach. Am. J. Physiol. 247, R8O6-R815.
GARLAND, T., JR (1988). Genetic basis of activity metabolism. I. Inheritance of speed, stamina, and antipredator displays in the garter snake Thamnophis sirtalis. Evolution 42, 335-350.
GARLAND, T., JR, BENNETT, A. F. AND DANIELS, C. B. (1990a). Heritability of locomotor performance and its functional bases in a natural population of vertebrates. Experientia 46, 530-533.
20 A. F. BENNETT
GARLAND, T., JR, ELSE, P. L., HULBERT, A. J. AND TAP, P. (1987). Effects of endurance tr and captivity on activity metabolism in lizards. Am. J. Physiol. 252, R450-R456.
GARLAND, T., JR, GEISER, F. AND BAUDINETTE, R. V. (1988). Comparative locomotor performance of marsupial and placental mammals. J. ZooL, Lond. 215, 505-522.
GARLAND, T., JR, HANKINS, E. AND HUEY, R. B. (19906). Locomotor capacity and social dominance in male lizards (Sceloporus occidentalis). Fund. Ecol. 4, 243-250.
GLEESON, T. T. (1979). Foraging and transport costs in the Galapagos marine iguana,
Amblyrhynchus cristatus. Physiol. Zool. 52, 549-557.
GLEESON, T. T. AND HARRISON, J. M. (1988). Muscle composition and its relation to sprint running in the lizard Dipsosaurus dorsalis. Am. J. Physiol. 255, R470-R477.
GLEESON, T. T., MITCHELL, G. S. AND BENNETT, A. F. (1980a). Cardiovascular responses to graded activity in the lizards Varanus and Iguana. Am. J. Physiol. 239, R174-R179.
GLEESON, T. T., PUTNAM, R. W. AND BENNETT, A. F. (1980£>). Histochemical, enzymatic, and contractile properties of skeletal muscle fibers in the lizard Dipsosaurus dorsalis. J. exp. Zool.
214, 293-302.
GNAIGER, E. (1983). Heat dissipation and energetic efficiency in animal anoxibiosis: economy contra power. J. exp. Zool. Z2&, 471-490.
GRIESHABER, M. K. AND GADE, G. (1976). The biological role of octopine in the squid, Loligo
vulgaris (Lamarck). J. comp. Physiol. 108, 225-232.
GRIESHABER, M. K. AND ZEBE, E. (1978). Die Bildung von Octopin bei Chlamys opercularis und bei Sipunculus nudus. Verh. dt. zool. Ges. 71, 266.
HARRISON, J. F., GLEESON, T. T. AND PHILLIPS, J. E. (1989). Gas exchange in jumping grasshoppers. Am. Zool. 29, 5A.
HECKROTTE, C. (1967). Relations of body temperature, size, and crawling speed of the common garter snake, Thamnophis s. sirtalis. Copeia 1967, 759-763.
HEDMRICH, B. (1977). Why have some animals evolved to regulate a high body temperature?
Am. Nat. Ill, 623-640.
HEMMINGSEN, A. M. (1960). Energy metabolism as related to body size and respiratory surfaces, and its evolution. Report Steno Mem. Hosp. Nord. Insulinlab. 9, 1-110.
HERREID, C. F. (1981). Energetics of pedestrian arthropods. In Locomotion Energetics of
Arthropods (ed. D. F. Herreid and C. R. Fourtner), pp. 491-526. New York: Plenum PTess. HOCHACHKA, P. W. (1980). Living without Oxygen: Closed and Open Systems in Hypoxia
Tolerance. Cambridge, MA: Harvard University Press.
HOCHACHKA, P. W. AND SOMERO, G. N. (1984). Biochemical Adaptation. Princeton, NJ: Princeton University Press.
HUEY, R. B. (1987). Phylogeny, history, and the comparative method. In New Directions in
Ecological Physiology (ed. M. E. Feder, A. F. Bennett, W. W. Burggren and R. B. Huey),
pp. 76-98. Cambridge, England: Cambridge University Press.
HUEY, R. B. AND BENNETT, A. F. (1986). A comparative approach to field and laboratory studies in evolutionary ecology. In Predator-Prey Relationships (ed. M. E. Feder and G. Lauder), pp. 82-98. Chicago: University of Chicago Press.
HUEY, R. B. AND BENNETT, A. F. (1987). Phylogenetic studies of coadaptation: preferred temperatures versus optimal performance temperatures of lizards. Evolution 41, 1098-1115.
HUEY, R. B. AND DUNHAM, A. E. (1987). Repeatability of locomotor performance in natural populations of the lizard Sceloporus merriami. Evolution 41, 1116-1120.
HUEY, R. B., DUNHAM, A. E., OVERALL, K. L. AND NEWMAN, R. A. (1990). Variation in locomotor performance in demographically known populations of the lizard Sceloporus
merriami. Physiol. Zool. 63, 845-872.
HUEY, R. B., PIANKA, E. R. AND SCHOENER, T. W (eds.) (1983). Lizard Ecology: Studies of a
Model Organism. Cambridge, MA: Harvard University Press.
HUTCHISON, V. H., DOWLING, H. G. AND VINEGAR, A. (1966). Thermoregulation in a brooding female Indian python, Python molurus bivittatus. Science 151, 694-696.
JAYNE, B. C. AND BENNETT, A. F. (1990a). Scaling of speed and endurance in garter snakes: a comparison of cross-sectional and longitudinal allometries. /. Zool., Lond. 220, 257-277.
JAYNE, B. C. AND BENNETT, A. F. (19906). Selection on locomotor performance capacity in a natural population of garter snakes. Evolution 44, 1204-1229.
The evolution of activity capacity 21
, A. E. (1981). Physiological mechanisms of thermoregulation. In Insect
Thermoregulation (ed. B. Heinrich), pp. 115-158. New York: Wiley-Interscience Pub.
KEMP, T. S. (1982). Mammal-like Reptiles and the Origin of Mammals. New York: Academic Press.
KOLOK, A. S. (1990). The swimming performance of juvenile largemouth bass: correlates with
physiological and morphological factors. Am. Zool. 30, 134A.
KRAM, R. AND TAYLOR, C. R. (1990). Energetics of running - a new perspective. Nature 346, 265-267.
LANDE, R. AND ARNOLD, S. J. (1983). The measurement of selection on correlated characters.
Evolution 37, 1210-1226.
LANGLOIS, B. (1980). Heritability of racing ability in thoroughbreds - a review. Livestock Prod.
Sci. 7, 591-605.
MAISEY, J. G. (1986). Heads and tails: a chordate phylogeny. Cladistics 2, 201-256.
MCMAHON, B. R. (1981). Oxygen uptake and acid-base balance during activity in decapod crustaceans. In Locomotion and Energetics in Arthropods (ed. C. F. Herreid and C. R. Fourtner), pp. 295-335. New York: Plenum Press.
MILES, D. B. (1987). Habitat related differences in locomotion and morphology in two populations of Urosaurus ornatus. Am. Zool. 27, 44A.
MILES, D. B. (1989). Selective significance of locomotory performance in an iguanid lizard. Am.
Zool. 29, 146A.
NORTHCUTT, R. G. AND GANS, C. (1983). The genesis of neural crest and epidermal placodes: a reinterpretation of vertebrate origins. Q. Rev. Biol. 58, 1-28.
OLSON, E. C. (1971). Vertebrate Paleozoology. New York: Wiley-Interscience.
PERRY, S. F. (1983). Reptilian lungs: functional anatomy and evolution. Adv. Anat. Embryol.
Cell Biol. 79,1-81.
PETERS, R. H. (1983). The Ecological Implications of Body Size. Cambridge, England: Cambridge University Press.
POUGH, F. H. (1978). Ontogenetic changes in endurance in water snakes (Natrix sipedon): physiological correlates and ecological consequences. Copeia 1978, 69-75.
POUGH, F. H. (1979). Summary of oxygen transport characteristics of reptilian blood.
Smithsonian herpet. Info. Serv. 45, 1-18.
POUGH, F. H. (1983). Amphibians and reptiles as low-energy systems. In Behavioral Energetics (ed. W. P. Aspey and S. I. Lustick), pp. 141-188. Columbus, OH: Ohio State University Press.
POUGH, F. H. (1989). Organismal performance and Darwinian fitness: approaches and interpretations. Physiol. Zool. 62, 199-236.
POUGH, F. H. AND ANDREWS, R. M. (1984). Individual and sibling-group variation in metabolism of lizards: the aerobic capacity model for the origin of endothermy. Comp.
Biochem. Physiol. 79A, 415-419.
PRESTWICH, K. N. (1988). The constraints on maximal activity in spiders. II. Limitations imposed by phosphagen depletion and anaerobic metabolism. J. comp. Physiol. B 158, 449-456.
RANDALL, D. J., BURGGRBN, W. W., FARRELL, A. P. AND HASWELL, M. S. (1981). The Evolution
of Air Breathing in Vertebrates. Cambridge, England: Cambridge University Press. REGAL, P. J. (1978). Behavioral differences between reptiles and mammals: an analysis of
activity and mental capabilities. In Behavior and Neurology of Lizards: An Interdisciplinary
Colloquium (ed. N. Greenberg and P. D. MacLean), pp. 183-202. Washington DC: National
Institutes of Health, Government Printing Office.
ROMER, A. S. (1966). Vertebrate Paleontology, 3rd edn. Chicago: University of Chicago Press.
RUBEN, J. A. (1976). Aerobic and anaerobic metabolism during activity in snakes. J. comp.
Physiol. 109, 147-157.
RUBEN, J. A. AND BATTALIA, D. E. (1979). Aerobic and anaerobic metabolism during activity in small rodents. J. exp. Zool. 208, 73-76.
RUBEN, J. A. AND BENNETT, A. F. (1980). Antiquity of the vertebrate pattern of activity metabolism and its possible relation to vertebrate origins. Nature 286, 886-888.
22 A. F. BENNETT
RUBEN, J. A. AND PARRISH, J. K. (1991). Antiquity of the chordate pattern of exercJ^ metabolism. Paleobiology (in press).
RYAN, J. E. (1975). The inheritance of track performance in greyhounds. Unpublished Master's thesis. Trinity College, Dublin, Ireland.
SCHLUTER, D. (1988). Estimating the form of natural selection on a quantitative trait. Evolution
42, 849-861.
SCHMIDT-NIELSEN, K. (1972). Locomotion: energy cost of swimming, flying, and running. Science 177, 222-228.
SEIGEL, R. A., COLLINS, J. T. AND NOVAK, S. S (eds) (1987). Snakes: Ecology and Evolutionary
Biology. New York: MacMillan Publ. Co.
SHAFFER, H. B., AUSTIN, C. C. AND HUEY, R. B. The physiological consequences of metamorphosis on salamander (Ambystoma) locomotor performance. Physiol. Zool. (in press).
SIEGMUND, B., GRIESHABER, M. K., REITZE, M. AND ZEBE, E. (1985). Alanopine and strombine are end products of anaerobic glycolysis in the lugworm, Aremcola marina. Comp. Biochem.
Physiol. 82B, 337-345.
STEVENS, E. D. AND NEILL, W. H. (1978). Body temperature relations of tunas, especially skipjack. In Fish Physiology, vol. 7 (ed. W. S. Hoar and D. J. Randall), pp. 315-359. New York: Academic Press.
SULLIVAN, B. K. AND WALSBERG, G. E. (1985). Call rate and aerobic capacity in Woodhouse's toad (Bufo woodhousei). Herpetologica 41, 404-407.
TAIGEN, T. L. (1983). Activity metabolism of anuran amphibians: implications for the origin of
endothermy. Am. Nat. 121, 94-109.
TAYLOR, C. R. (1980). Evolution of mammalian homeothermy: a two-step process?. In
Comparative Physiology: Primitive Mammals (ed. K. Schmidt-Nielsen, L. Bolis and C. R.
Taylor), pp. 100-111. Cambridge, England: Cambridge University Press.
TAYLOR, C. R. (1982). Scaling limits of metabolism to body size: implications for animal design. In A Companion to Animal Physiology (ed. C. R. Taylor, K. Johansen and L. Bolis), pp. 161-170. Cambridge, England: Cambridge University Press.
TAYLOR, C. R., HEGLUND, N. C. AND MALOIY, G. M. O. (1982). Energetics and mechanics of
terrestrial locomotion. I. Metabolic energy consumption as a function of speed and body size in birds and mammals. J. exp. Biol. 97, 1-21.
TAYLOR, C. R., KARAS, R. H., WEIBEL, E. R. AND HOPPELER, H. (1987). Adaptive variation in the mammalian respiratory system in relation to energetic demand. II. Reaching the limits to oxygen flow. Respir. Physiol. 69, 7-26.
TAYLOR, C. R., MALOIY, G. M. O., WEIBEL, E. R., LANGMAN, V. A., KAMAU, J. M. Z., SEEHERMAN, H. J. AND HEGLUND, N. C. (1981). Design of the mammalian respiratory system.
III. Scaling maximum aerobic capacity to body mass: wild and domestic mammals. Respir.
Physiol. 44, 25-37.
TAYLOR, C. R., SCHMIDT-NIELSEN, K. AND RAAB, J. L. (1970). Scaling of energetic cost of running to body size in mammals. Am. J. Physiol. 219, 1104-1107.
TAYLOR, C. R. AND WEIBEL, E. R. (1981). Design of the mammalian respiratory system. I. Problem and strategy. Respir. Physiol. 44, 1-10.
TAYLOR, E. B. AND MCPHAIL, J. D. (1985). Burst swimming and size-related predation of newly emerged coho salmon Oncorhynchus kisutch. Trans. Am. Fish. Soc. 114, 546-551.
THOMAS, R. D. K. AND OLSON, E. C. (eds) (1980). A Cold Look at the Warm-Blooded
Dinosaurs. Am. Ass. Advanc. Sci. Selected Symp. 28. Boulder, CO: Westview Press. TOLLEY, E. A., NOTTER, D. R. AND MARLOWE, T. J. (1983). Heritability and repeatability of
speed for 2- and 3-year-old standardbred racehorses. /. Anim. Sci. 56, 1294-1305.
TSUJI, J. S., HUEY, R. B., VAN BERKUM, F. H., GARLAND, T., JR AND SHAW, R. G. (1989). Locomotor performance of hatchling fence lizards (Sceloporus occidentalis): quantitative genetics and morphometric correlates. Evol. Ecol. 3, 240-252.
TUCKER, V. A. (1975). The energetic cost of moving about. Am. Sci. 63, 413-419.
The evolution of activity capacity 23
piN BERKUM, F. H. AND TSUJI, J. S. (1987). Inter-familial differences in sprint speed of hatchling lizards (Sceloporus occidentalis). J. Zooi, Lond. 212, 511-519.
WALTON, B. M. (1988a). Relationships among metabolic, locomotor, and field measures of organismal performance in the Fowler's toad (Bufo woodhousei fowleri). Physiol. Zool. 61, 107-118.
WALTON, B. M. (1988ft). The adaptive significance of activity metabolism among anuran amphibians. PhD thesis, University of Chicago.
WALTON, M., JAYNE, B. C. AND BENNETT, A. F. (1990). The energetic cost of limbless locomotion. Science 249, 524-527.
WATT, W. B. (1986). Power and efficiency as indices of fitness in metabolic organization. Am.
Nat. 127, 629-653.
WELLS, K. D. AND TAIGEN, T. L. (1984). Reproductive behavior and aerobic capacities of male American toads {Bufo americanus): is behavior constrained by physiology? Herpetologica 40, 292-298.
WELLS, R. M. G. AND DAVIE, P. S. (1985). Oxygen binding by the blood and hematological effects of capture stress in two big gamefish: mako shark and striped marlin. Comp. Biochem.
Physiol. 81A, 643-646.
WITHERS, P. C. AND HILLMAN, S. S. (1979). Oxygen consumption of Amphiuma means during forced activity and recovery. Comp. Biochem. Physiol. 69A, 141-144.
WITHERS, P. C. AND HILLMAN, S. S. (1988). A steady-state model of maximal oxygen and carbon dioxide transport in anuran amphibians. /. appl. Physiol. 64, 860-866.
ZEBE, E., SALGE, U., WIEMANN, C. AND WILPS, H. (1981). The energy metabolism of the leech
Hirudo medicinalis in anoxia and muscular work. J. exp. Zool. 218, 157-163.