THE MAXIMUM SHORTENING VELOCITY OF
HOLOTHURIAN MUSCLE AND EFFECTS OF
TONICITY CHANGE ON IT
BY T. TSUCHIYA*
Bermuda Biological Station for Research, Ferry Reach 1-15, Bermuda
Accepted 8 May 1985
SUMMARY
1. The maximum shortening velocity of the longitudinal retractor muscle of the holothurian, Isostichopus badionotus, was measured using a 'slack test' in solutions of normal, increased and reduced osmotic strength.
2. At normal osmotic strength, the average maximum shortening velocity was 0-44 ± 0-04 LoS"1 (N=14) which is fast compared to other smooth muscles. This is unexpected considering the slow locomotion of these animals.
3. Raising the osmotic strength by 55 % by addition of sucrose reduced the shortening velocity by more than 50 %. Raising the osmotic strength by 52 % with NaCl caused a 35 % reduction in shortening velocity.
4. Lowering the osmotic strength by 19% or 39%, by reducing NaCl concentration, did not significantly affect shortening velocity.
5. The shortening velocity was found to be proportional to the volume of the muscle preparation.
INTRODUCTION
Holothurians have five longitudinal retractor muscles which run along the inner surface of the body and these muscles are supposed to be for locomotion. A. V. Hill (1926) and Levin & Wyman (1927) suggested that there were fundamental similarities between mechanical properties of these smooth muscles and vertebrate skeletal mus-cle, although the rate of contraction was much slower in the holothurian muscle (see review by Takahashi, 1966). In the present study, the maximum shortening velocity of holothurian smooth muscle was determined by the 'slack test' (Edman, 1979) and the effect of osmotic strength was examined.
MATERIALS AND METHODS
Preparations
The holothurians Isostichopus badionotus were collected from the seashore near Bermuda Biological Station immediately before dissection. They were cut open and
•Present address: Department of Physiology, School of Medicine, Teikyo University, Itabashi-ku, Tokyo 173, Japan.
32
T . TSUCHIYA1
SM
H, —— P , H2
o
[image:2.451.75.377.45.310.2]In
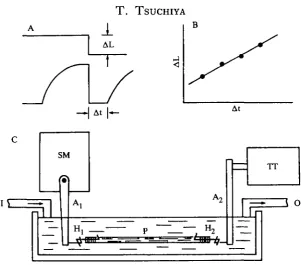
Fig. 1. Schematic diagram showing the analysis of records (A, B) and experimental arrangement (C). (A) The upper and lower traces represent the length and tension changes respectively. The tension falls to zero when the muscle is quickly released by AL during plateau of isometric tension and redevelops after a short delay, At, which is the duration of unloaded shortening. (B) AL is plotted against At and the maximum shortening velocity is calculated from the slope of the line. (C) Aj, arm of servo-motor; A2, arm of tension transducer; Hi, H2, hooks attached to both ends of a preparation by silk thread; I, inlet for bath solution; O, outlet for bath solution; P, preparation; SM, servo-motor; T T , tension transducer.
Table 1. Composition of solutions
Solution IT l-28TNa l-S2TNa l-28TSu ISSTSu 0-81TNa 0-61TNa NaCl (mmol I"1)
462 603 745 462 462 349 235 KC1 (mmol r1)
10 10 10 10 10 10 10 CaCl2 (mmol P1)
11 11 11 11 11 11 11 MgCl2 (mmol 1 48 48 48 48 48 48 48 NaHCO3 ') (mmol P1)
6 6 6 6 6 6 6 Sucrose (mmol P1)
283
567 —
Osmolality (mosmol kg"1)
1030 1320 1570 1320 1600 830 630
[image:2.451.42.412.431.526.2]o
L10- 7 10- 6 10-5 lO"4
[image:3.451.76.363.58.300.2]Concentration of ACh (mol 1
Fig. 2. Relationship between the peak height of contracture tension and acetylcholine (ACh) con-centration.
Solutions
Solutions were as shown in Table 1. The solutions were named according to the osmolality relative to the standard solution (IT solution) and to the constituent responsible for the tonicity change, for example, l*28TNa represents an osmolality 1-28 times that of the standard solution, using NaCl. The osmolality (mosmolkg"1 water) of salt solutions was measured directly with a freezing point osmometer (Ad-vanced Instruments Inc. Model 3L). The osmolality of the sucrose solution was calculated according to the formulae provided by Dydyriska & Wilkie (1963) and that of the solutions containing both salt and sucrose was taken to be the sum of measured and calculated values.
Measurement of shortening velocity
To determine the shortening velocity of an unloaded fibre preparation, a contrac-ture was first induced using acetylcholine (ACh). During the plateau phase, the arm of the servo-motor was moved to allow the fibre to slacken (20 ms for completion of a length step) and the time taken for redevelopment of tension (At) was plotted against the size of the length step (AL) (Fig. 1 A,B). Data points showed a close fit to a straight line, using the least-squares methods. Length steps were within the range of 0-32 of a fibre length (Lo).
Measurement of fibre volume
34
T . TSUCHIYAD
[image:4.451.98.360.47.319.2]0-5 s
Fig. 3. Redevelopment of tension after quick release of muscle preparation during plateau of isometric tension in three different length steps (A, D, 019L«; B, E, 0-23 Lo; C, F, 0-3 L«) and at two different tonicities of the extracellular medium (A, B, C, in I T solution; D, E, F, in l-28TSu solution).
along the length of the strip preparation. The muscle preparations were equilibrated for 20 min in the standard, hypertonic or hypotonic solutions. They were then lightly blotted with filter paper, wrapped with aluminium foil to avoid evaporation and weighed.
RESULTS
Tension development by acetylcholine
For all the measurements of maximum shortening velocity, tension was developed by application of acetylcholine (ACh), which is generally believed to be the trans-mitter in the neuromuscular system of echinoderms (R. B. Hill, 1970; Pentreath & Cobb, 1972; Suzuki, 1982). The isometric tension showed very gradual relaxation during prolonged application of ACh. Tension was reduced by only 5 % after 45 s. A dose-response study of the effects at 10 s showed that ACh concentrations above 3 X 10~s mol I"1 produced maximum tension (Fig. 2). This dose is a little lower than that for the muscle of another holothurian (Sugi et al. 1982).
An ACh concentration of 10~5moll~1 was adopted as the standard for further experiments, rather than 3 x 10~smoll~1, because the muscle could break with repeated full activation.
Effects of acetylcholine concentration on Vm and the dependence on time after the activation
0-25
a 0-2
u
(A
•s
GO
jj 015
0-2 0-4
0-25
0-2
jj 0-
0-150 0-4 0-8 Duration of unloaded shortening (s)
Fig. 4. (A) Plots of length change versus duration of unloaded shortening in three different con-centrations of ACh ( • , 10"*moirl; O, 5 X K T ' m o i r1; • , 10~! moll"1). The maximum shortening velocity was calculated from the slope of the fitted straight line. (B) Plots of length change versus duration of unloaded shortening at two different times ( • , 10 s; O, 45 s) after the application of ACh.
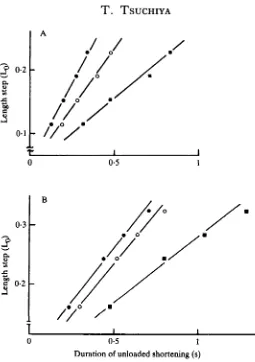
quickly released during the plateau tension induced by 10 5 mol 1 1 ACh. The tension fell to zero at the onset of release and redeveloped after a delay (Fig. 3). It is clearly shown that the duration of unloaded shortening increases with an increase of step length (A, 0 • 19 Lo; B, 0 • 23 Lo; C, 0 • 3 Lo). The step lengths were plotted against the durations of unloaded shortening (Fig. 4). The data showed a good fit to a straight line, and the slope gave the shortening velocity. The velocity was constant at ACh concentrations greater than 5 X 10~6moll~1 (Fig. 4A).
Whereas isometric tension showed only a very gradual reduction as the ACh exposure was prolonged, the maximum shortening velocity was more strongly affected. After45 s of ACh, the velocity was only 50 % of the value at 10 s (Fig. 4B). Shortening velocities were therefore measured 10 s after the activation by ACh for further experiments. The averaged maximum shortening velocity of 14preparationswasO-44 ± 0-04 Los"1.
Effects of hypertonicity on the maximum shortening velocity
[image:5.451.110.327.92.371.2]36
t
a
2
o-i0-5
0-3
a. u
[image:6.451.91.346.46.408.2]0 0-5 1 Duration of unloaded shortening (s)
Fig. 5. Plots of length change versus duration of unloaded shortening in increased osmotic strength made by addition of sucrose (A, • , in IT; O, in l-28TSu; • , in l-55TSu) or NaCl (B, • , in IT; O, in l-28TNa; • , in l-52TNa).
In solutions made hypertonic with sucrose, the duration of the unloaded shortening was longer for a given length step, and the isometric tension was lower, than in the standard solution (compare Fig. 3D-F with Fig. 3A-C). The relationship between the amplitude of the length steps and the duration of unloaded shortening is illustrated in Fig. 5A, in which the maximum shortening velocities in IT (the standard solution), l-28TSu and l-55TSu solutions are 0-48Los"1, 0-38Los"1 and 0-22 Los"1 respec-tively; i.e. with relative values of 1:0-79:0-46. Average relative values were 0-78 (S.D. 0-16; N = 4) for 1 -28TSu, and 0-42 (S.D. 0-05, TV = 4) for 1 -55TSu. The strong effect of the hypertonic sucrose solution upon the maximum shortening velocity, and the observation that the rate of the tension redevelopment is slower in hypertonic solution (Fig. 3), suggests that the turnover rate of crossbridges may be slower in hypertonic solution.
0-3
s
£ 0-2
CO c
0-2
Duration of unloaded shortening (s)
[image:7.451.86.343.46.305.2]0-4
Fig. 6. Plots of length change versus duration of unloaded shortening in reduced oemotic strength. • , in IT; O, in 0-81TNa; • , in 0-61TNa.
shortening velocities in I T , 1-28TNa and 1-52TNa solutions are 0-47 Lo s"1,0-47 Lo s"1 and 0-30 Lo s"1 respectively; with relative values of 1:1:0-64. Average relative short-ening velocities were 0-91 (S.D. 0-l,N = 4) for l-28TNa and 0 5 6 (S.D. 0-05, AT = 4 ) for l-52TNa.
Effects of hypotonicity on the maximum shortening velocity
After measurement of shortening velocity in the standard solution, the preparation was equilibrated for 20min in hypotonic solution. Shortening velocity was little affected. In Fig. 6, the shortening velocities in IT, 0-81TNa and 0-61TNa solutions are 0-58LOS"1, 0-54LOS"1 and 0-67Los-1 respectively; with relative values of 1:0-93:1-16. The average relative shortening velocities were 0-98 (S.D. O04, ./V = 4) inO-81TNaand 1-05 (S.D. 0-15, N = 4) in 0-61TNa.
Tension changes in hypertonic and hypotonic solution
The tension induced by 10~5 mol I"1 ACh was not significantly altered in solutions made hypertonic by addition of NaCl, or in hypotonic solution. In hypertonic sucrose solutions, however, tension decreased significantly both in l-28TSu and in l-55TSu (Fig. 7A). It was observed that in a few preparations appreciable tension developed in hypertonic or hypotonic solution without ACh and these fibres were not used for the measurements of tension or shortening velocity.
Volume changes in hypertonic and hypotonic solutions
38
T . TSUCHIYAwas attained within 20min after a solution change. There was a large reduction in volume in solutions made hypertonic with sucrose, while solutions made hypertonic with NaCl had less effect and hypotonic solutions produced no appreciable change (Fig. 7B).
1 0
05
-0-5 1 0 1-5
1 0
a o-9
o
0-8
0-7
0-5 10
Relative osmotic strength
1-5
[image:8.451.108.324.136.565.2]DISCUSSION
The average maximum shortening velocity of the longitudinal retractor muscle of Isostichopus badionotus was 0-44Los"1. In cat muscle (Close & Hoh, 1967), it is 22-8 Lo s"1. In smooth muscles of pig carotid artery (Herlihy & Murphy, 1974), dog trachea (Stephens, Kroeger & Mehta, 1969), rabbit urinary bladder (Uvelius, 1976) and guinea pig taenia coli (Mashima, Okada & Okumura, 1979), the velocities are 0-12, 0-17, 0-23 and 0-53 LOs"1 respectively. For invertebrate smooth muscle, the shortening velocities have been found to be 0-12—0-25 Los"1 for anterior byssus retractor muscle of Mytilus (Lowy & Millman, 1963; Tameyasu & Sugi, 1976) and 0-42L0S"1 for the foot muscle of Mytilus (Ishii & Takahashi, 1981). Thus, the shortening velocity of holothurian muscle is fast compared with that of other smooth muscles, and not slow as might be expected from the slow locomotion of these animals. Therefore, it is conceivable that the fast contraction of this muscle is not utilized in the locomotion of this animal, but in some other behaviour.
In striated muscle, the maximum shortening velocity, which is much faster than that in smooth muscle, has been shown to decrease remarkably in hypertonic solution and to increase appreciably in hypotonic solution (Edman & Hwang, 1977). If a fundamental mechanism of contraction is common to striated and smooth muscles, it may be that similar phenomena are observed in smooth muscles. Little information is at present available about the effects of hypertonic solution on the contractile tension and the shortening velocity in smooth muscles except for Mytilus (Sugi, Yamaguchi & Tanaka, 1977; Tameyasu, 1978). An essential step in the analysis of the maximum shortening velocity is to find out whether the tension level before the release affects the velocity. In holothurian muscle, shortening velocity is apparently not related to the tension at ACh concentrations above 5 X 10~6 mol I"1 (Fig. 4). Shorten-ing velocity is, however, related to volume (Figs 5, 6, 7). In striated fibres it is suggested that the effect of volume upon velocity could be by change in ionic strength (Edman & Hwang, 1977). But it has recently been demonstrated that the maximum shortening velocity decreases under constant ionic strength in osmotically compressed skinned fibres of frog, suggesting that the decrease in the shortening velocity is a direct effect of the decrease in inter-filament spacing (Tsuchiya, 1984).
The reduction of velocity during ACh exposure (Fig. 4B) is reminiscent of the catch phenomenon in the anterior byssus retractor of Mytilus (Tameyasu & Sugi, 1976; Sugi & Tsuchiya, 1979). A similar phenomenon has also been reported in striated muscle (Julian & Sollins, 1972). The mechanism responsible for these phenomena is still incompletely understood, though ADP has been found to slow down the cycling rate of ATP hydrolysis of crossbridge movement (Giith, Kuhn, Tsuchiya & Ruegg, 1981).
Further analysis of holothurian muscle or smooth muscle may be possible using glycerinated or chemically-skinned preparations, thus enabling independent control of inter-filament spacing and ionic environment as in skeletal muscle (Tsuchiya, 1984).
40 T . TSUCHIYA
Maxwell Fellowship, through the Bermuda Biological Station for Research. He also wishes to express his sincere thanks to Professor R. B. Hill for kind guidance about the Bermuda Biological Station and to Professor H. Sugi for affording an opportunity of working in Bermuda.
R E F E R E N C E S
CLOSE, R. & HOH, J. F. Y. (1967). Force-velocity properties of kitten muscles. J. Phytiol., Land. 192, 815-822. DYDYNSKA, M. & WILKIE, D . R. (1963). The osmotic properties of striated muscle fibres in hypertonic
solutions. J. Physiol, bond. 169, 312-329.
EDMAN, K. A. P. (1979). The velocity of unloaded shortening and its relation to sarcomere length and isometric force in vertebrate muscle fibres..?. Physiol., Land. 291, 143-159.
EDMAN, K. A. P. & HWANG, J. C. (1977). The force-velocity relationship in vertebrate muscle fibres at varied tonicity of the extracellular medium. J. Physiol., Land. 269, 255—272.
GOTH, K., KUHN, H. J., TSUCHIYA, T . & ROEGG, J. C. (1981). Length dependent state of activation - length change dependent kinetics of crossbridges in skinned insect flight muscle. Btophys. struct. Mech. 7, 139-169. HILL, A. V. (1926). The viscous elastic properties of smooth muscle. Proc. R. Soc. B 100, 108-115. HILL, R. B. (1970). Effects of some postulated neurohumors on rhythmicity of the isolated cloaca of a
holothurian. Physiol. Zool. 43, 109-123.
HEJUJHY, J. & MURPHY, R. A. (1974). Force-velocity and series elastic characteristics of smooth muscle from the hog carotid artery. Circulation Res. 34, 461—466.
ISHII, N. & TAKAHASHI, K. (1981). Mechanical properties and fine structure of the pedal retractor muscle of
Mytilus eduHs. Comp. Biochem. Physiol. 70A, 275-283.
JULIAN, F . J. & SOLLJNS, M. R. (1972). Regulation of force and speed of shortening in muscle contraction.
Symp. quant. Biol. 37, 635-646.
LEVIN, A. & WYMAN, J. (1927). The viscous elastic properties of muscle. Pmc. R. Soc. B 101, 218-243. LOWY, J. & MILLMAN, B. M. (1962). The contractile mechanism of the anterior byssus retractor muscle of
Mytilus edulis. Phil. Trans. R. Soc. Ser. B 246, 105-148.
MASHIMA, H., OKADA, T . & OKUMUBA, H. (1979). The dynamics of contraction in the guinea pig taenia coli.
Jap. J. Physiol. 29, 85-98.
PENTREATH, V. W. & COBB, J. L. S. (1972). Neurobiology of echinodermata. Biol. Rev. 47, 363-392. STEPHENS, N. L., KKOEGER, E. & MEHTA, J. A. (1969). Force-velocity characteristics of respiratory airway
smooth muscle. J. appl. Physiol. 26, 685-692.
SUGI, H., SUZUKI, S., TSUCHIYA, T . , GOMI, S. & FUJIEDA, N. (1982). Physiological and ultrastructural
studies on the longitudinal retractor muscle of a sea cucumber Stichopus japonicus. I. Factors influencing the mechanical response. J. exp. Biol. 97, 101-111.
SUGI, H. & TSUCHIYA, T . (1979). The change in the load-sustaining ability and in the series elasticity inMytilus smooth muscle during isotonic shortening. J. Physiol., Land. 288, 635—648.
SUGI, H., YAMAGUCHI, T . & TANAKA, H. (1977). The effect of hypertonic solutions on contracture tension and volume in Mytilus smooth muscle. Comp. Biochem. Physiol. 58A, 405—407.
SUZUKI, S. (1982). Physiological and cytochemical studies on activator calcium in contraction by smooth muscle of a sea cucumber, Isostichopus badionotus. Cell Tissue Res. 222, 11—24.
TAKAHASHI, K. (1966). Muscle physiology. lnPhysiologyofEchinoderm, (ed. R. A. Boolootian), pp. 513-527. New York: Interscience Publishers.
TAMEYASU, T . (1978). The effect of hypertonic solutions on the rate of relaxation of contracture tension in
Mytilus smooth muscle. J . exp. Biol. 74, 197-210.
TAMEYASU, T . & SUGI, H. (1976). The series elastic component and the force-velocity relation in the anterior byssal retractor muscle of Mytilus edulis during active and catch contractions. J. exp. Biol. 64, 497—510. TSUCHIYA, T . (1984). Changes in mechanical properties in osmotically compressed skinned muscle fibers of
frog. In Contractile Mechanisms in Muscle, (edsG. H. Pollack & H. Sugi), pp. 731—733. New York: Plenum Publishing Corporation.