THE ORIGIN AND FATE OF UREA IN THE
DEVELOPING HEN'S EGG
BY JOSEPH NEEDHAM, JEAN BRACHET
AND ROBERT K. BROWN.
(From the Biochemical Laboratory, Cambridge.)
(Received 20th February, 1935.)
(With Three Text-figures.)
CONTENTS.
PAGE
I. T h e formation of urea by the chick embryo . . 321 (1) Introduction . 3 2 1 (2) Material and methods . . . 323 (3) Experimental results . . . . 323 («) Production of urea from ammonia . . . 323 (A) Production of urea by the ornithine cycle . 324 (c) Production of urea from uric acid . . . 324 (d) P r o d u c t i o n o f u r e a f r o m a r g i n i n e . . . 3 2 4
I I . U r e a a n d t h e f o r m a t i o n o f u r i c a c i d i n t h e e g g . . 3 2 0 ( 1 ) I n t r o d u c t i o n . . . 3 2 9 ( 2 ) M a t e r i a l a n d m e t h o d s . . . 3 3 0 ( 3 ) E x p e r i m e n t a l r e s u l t s . . . 3 3 0
I I I . S u m m a r y . . . 3 3 4 A c k n o w l e d g m e n t s . . . 3 3 5 R e f e r e n c e s . . . 3 3 5
I. THE FORMATION OF UREA BY THE CHICK EMBRYO.
(1) INTRODUCTION.
SINCE the work of Needham (1926), reported in this Journal, the question of the
excretion of nitrogen by the chick embryo during its development has not received much further consideration. In this work it was shown that the embryo at an early stage of its development (fourth to seventh day) may excrete more urea than uric acid, but that later the elimination of uric acid greatly exceeds that of urea. On the tenth day of development, uric acid may form 95 per cent, of the excreted nitrogen.
protein deamination. That the embryo at this age can deaminate amino acids appeared from the work of Glover (1931), who observed small increases of ammonia and urea when embryos were shaken in vitro with amino acids.1
Since up to the present time no definite information about the origin of the urea of the chick embryo has been forthcoming, the following hypotheses presented themselves for examination. (1) The urea might arise, as it would do according to classical theory in a ureotelic animal, from the direct combination of carbon dioxide and ammonia. (2) The urea might be formed, as it is in ureotelic organisms, by means of the ornithine cycle of Krebs and Henseleit (1932). Working with mammalian liver slices, these authors, as is well known, found that ornithine was the only amino acid which would give rise to urea, and that it would do so even when present in minute quantity. Ornithine, in the presence of carbon dioxide and ammonia, forms citrulline, this with another molecule of ammonia forms arginine, and the arginine, acted upon by arginase, gives urea and ornithine. (3) The urea, however, might also be formed from uric acid. Przylecki and Rogalski (1928) showed that the chick embryo, during the first week of its development, is capable of destroying uric acid by a uricase. Unfortunately they did not investigate the products of its action (allantoin, allantoic acid, urea, etc.), so that it might equally well be an enzyme similar to that discovered by Stransky (1933) in Amphibia and fishes, which breaks uric acid down to urea; or the common uricase of mam-mals which only carries the process as far as allantoin. As Przylecki's chick uricase disappears from the embryo at about the time when the excretion of uric acid begins in earnest, the urea formed beforehand might well be derived from this source. (4) Finally, the urea might be derived simply from the action of arginase on arginine. The first of these possibilities was the most unlikely, since there is now little reason for supposing that direct synthesis ever occurs. The recapitulatory aspect of the phenomena would be satisfied by any of the other three possibilities, for the ornithine cycle is present in Amphibia and reptiles, the uricase of Stransky is present in fishes and Amphibia,2 and the presence of arginase, even in high activity, is widespread throughout invertebrates and vertebrates (cf. Baldwin, 1935). If the ornithine cycle were present in the chick embryo, it would be retaining for a transient period a mechanism which had been of value to the Amphibia and which has also been so to the mammals; if on the other hand the embryo derived its urea solely from arginine, it would possess an adult characteristic. In 1932 Clementi clearly showed that neither ammonia nor purines, but only arginine, produces urea in the adult hen. Hens on a normal diet excrete some 70 mg. urea daily, and in hunger this is reduced to about 20 mg. If the animals are then given 350 mg. arginine parenterally, the output goes up to normal. No other substance will bring about this effect.
1
The results of Kaieda (1930) were very confusing. Injecting amino-acids into the hen's egg at o days and estimating the urea at 16 days, he got small increases with some and decreases with others. Ammonium carbonate gave no increase. His control value at 16 days was three time9 as high as that usually accepted.
1
In order to settle the question, we have added ammonia, ammonia and ornithine, uric acid, and arginine to chick embryos in vitro, following subsequently the formation of urea.
(2) MATERIAL AND METHODS.
The White Ixghorn eggs used were obtained from the pure-bred flocks of the Cambridge University Farm, Messrs Chivers, and the Papworth Industries. Mo9t of the experiments were made on embryos of 4 ! or 5 days of development. After separation from amnion, allantois and yolk-sac, the embryos were washed in Ringer solution of the same composition as that used by Benzinger and Krebs (1933) in their work on uric acid formation, previously brought to 370 C. Yolk-sacs, when used, were freed from yolk and white by being dragged for some time over clean dry glass surfaces, after having been washed in Ringer solution. Weighed amounts of tissue were then incubated in suitable vessels slowly agitated for a known period in a thermostatic water-bath at 3 8° C. Buffering at pH 7-4 was accomplished by filling the vessels with a gas mixture of 95 per cent. Oa and 5 per cent. CO2 in
equilibrium with the bicarbonate in the Benzinger-Krebs solution.
After a sufficient time had elapsed (usually 4 hours) an aliquot portion of the suspension liquid was pipetted off, and the urea contained in it estimated according to the delicate manometric method of Krebs and Henseleit (1932) in which the carbon dioxide arising from the decomposition of urea by urease is measured. We express our results in the same manner as these authors:
™s. _ cmm. urea CO2
^ " mg. dry weight x hours *
Dry weights were calculated according to the data in the literature (Byerly, 1932; Needham, 1931), since the material is known to be very constant.
As controls, numerous experiments were made in order to determine the Qn
of the embryos and the membranes without the addition of any substances. The results are given in Table I. Another control was regularly made on the solutions added, in the absence of any tissue; this was always very small, but was deducted from the experiments in which tissue was present.
(3) EXPERIMENTAL RESULTS.
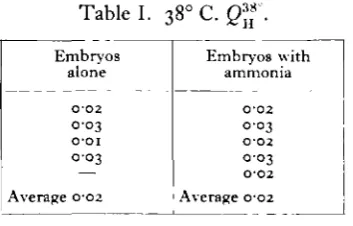
(a) Production of urea from ammonia.
To the Ringer-bicarbonate solution was added an amount of ammonium chloride equivalent to 31-8 mg. NH3 per cent. This concentration of ammonia was
that which in the experiments of Krebs and Henseleit (1932, p. 57) gave the highest syntheses of urea by means of the ornithine cycle in mammalian liver. Table I shows the values found for embryos of 4^—5 days' incubation age. It must be concluded that the urea production at this stage is extremely small (some 100 times less than the starving mammalian liver), and that it is not raised appreciably by the addition of ammonia.
(yolk-sacs). Here also the addition of ammonia caused not the slightest increase in the urea formation, which normally was even lower than that of the embryo (Qu 0-005).
Table I.
Embryos alone
O-O2 0 0 3
O O I
0 0 3 —
Average 002
38° C. Q™ .
Embryos with ammonia
O-O2
0-03
O'O2 OO3 O-O2
Average 002
(b) Production of urea by the ornithine cycle.
To the Ringer-bicarbonate solution was added as before 32 mg. per cent, of ammonia in the form of ammonium chloride, and in addition amounts varying from 4 to 28 mg. per cent, of ornithine. On mammalian liver, the maximum effect is reached at about 30 mg. per cent, ornithine.
The results showed clearly that at this stage the chick embryo does not possess the ornithine cycle. No increase in urea formation was observed in six experiments with different concentrations of ornithine. The Qu was always between o-oi and
0-02 and never rose above that of embryos maintained in physiological solution only. In two further experiments the yolk-sac was found also to lack the ornithine cycle, as no increase above the controls could be observed.
(f) Production of urea from uric acid.
The reasons which make it worth while to test the possibility that chick embryo urea could arise from uric acid have already been mentioned.
The uric acid was added to the Ringer-bicarbonate solution in the form of sodium urate 84-5 mg. per cent. In some experiments a higher concentration (132 mg. per cent.) was obtained by raising the pH to 9-5 by means of a standard glycine buffer. In some cases the embryos were ground to a brei instead of being used intact. But whatever the conditions of the experiment might be, it was never possible to observe an augmentation of the urea formation when uric acid was present (seven experiments). Three experiments were made on the yolk-sac both at/>H 7-5 and 9-5, but here also there was no increase over the controls.
These negative results clearly show that the uricase of the chick embryo studied by Przylecki and Rogalski (1928) does not resemble that of fishes and Amphibia, but probably like that of mammals only decomposes uric acid as far as allantoin.
(d) Production of urea from arginine.
The Ringer-bicarbonate solution was now made up to contain 393 mg. per cent, arginine. In these conditions a Qa some 15-20 times higher than the controls was
[image:5.451.137.317.129.269.2]very regularly found. Table II gives the results for 4J-5 day embryos:
Table II. Arginase in 4^-5 day embryos.
Control, average Arginine
Average
O: w >
O O 2
0-46 0-58
0-50 O-43
0-33
0 2 1 0 2 0
0-19 0-32
This demonstrates the existence of an active arginase, giving amounts of urea of only about four times less than the starving mammalian liver by the ornithine cycle.
The 5th day yolk-sac also appears to possess active arginase, though not in such great amount as the embryo. The results are shown in Table III. All these findings suffice for the conclusion that the urea normally produced by the chick embryo is derived entirely from the breakdown of arginine.
Table III. Arginase in §th day yolk-sac.
Controls
Average Arginine
Average
0-005 0-005 0-005 0-005 0-03 0-04 0-015 O-O2 O-O25
[image:5.451.142.308.375.510.2]connection with the disintegration (or synthesis) of the arginine contained in the protamines and histones. It may here be noted that according to Lieben and Lieber (1934) arginase has no effect upon combined arginine; the substrate must be free. This, however, would not affect the possibility that the significance of arginine in growing tissues was synthetic. The generalisations of Edlbacher rest almost wholly on work with tumour tissue (Edlbacher and Kutscher, 1931, 1932; Edlbacher, Roller and Becker, 1934; Edlbacher and Jung, 1934); only in the paper of Edlbacher and Merz (1927) is there any information about embryonic tissue, and here the results, which concerned guinea-pig material, were few and lacked precision re-garding the age and dimensions of the embryos studied.
It therefore seemed indispensable to study the activity of arginase in relation to the progress of development. We know that the growth rate of the embryo declines progressively, and if Edlbacher's view were correct, one would expect a progressive diminution of the Qn during incubation. Table IV shows that this is actually the
case. The technique described above was used, except that in the case of the larger
Table IV. Change of arginase activity with age in the chick embryo.
Day
2 l
3
3i
8
1 2
Hatched chick
Oft
I O I
i - i 7 i - i 7 I -32
0-87
°-53
o-io 0-08 0-08 0-05 0-04 0-04 0-05
Average 1-16
Average 0 6 3
Average o-oy
Average 0 0 4
embryos (8 and 12 days) a mince made with a razor was substituted for the intact embryos. The last value in this table, that for the hatched chick, was not obtained in the present investigation, but is calculated from the measurements made by Chaudhuri (1927) with the method of Edlbacher and Rothler (1925). The agree-ment is good.
The relative decline of arginase activity may conveniently be represented upon a heterogonic diagram, as in Fig. 1, where the cmm. urea CO2 liberated by one
will be seen that arginase belongs to the former group of substances, and falls off relatively as growth proceeds.
It may however be asked whether the decline of activity during development is not a property common to all the enzymes of the embryo. The older literature (summarised in Needham, 1931) contained so few accurate determinations of enzymic activity referable to dry weight of tissue, that an answer to this question would have been impossible till recently. But the work of Ammon and Schiitte (1935) now provides a comparison, for in measuring the relative activities of tri-butyrinase, esterase, and choline esterase during development, it was found that
10 100 1000 M g m . dry wt. of e m b r y o
F i g . 1. Negative Heterogony of Arginase.
the two first-named enzymes continuously increased (units/gm. dry weight) while the choline esterase increased to a maximum and thereafter decreased. This is in harmony with what is known about the greater importance of fat metabolism in the later periods of embryonic life, and suffices to show that all enzymes do not exhibit the negative heterogony of arginase.
and Ziese, 1934; Purr and Weil, 1934; Karrer and Zehender, 1934), it might be supposed that the decreasing activity of arginase during development was due to the decreasing quantity of activators present. In order to test this possibility, glutathione, cystein, and ascorbic acid were added, each in M/100 concentration, to the 8 day embryos, but in no case could any increased activity over the normals
(Qn 0-09) be observed. The same experiments were made with 5th day membranes,
which show a low Qu, but the same results were obtained, see Table V. From
Table V. Effect of activators on urea production.
8th
8th 8th 8th
5th
5 * 5th 5th
day
day day day
day
day day day
embryo, embryo, embryo, embryo,
yolk-sac, yolk-sac, yolk-sac, yolk-sac,
alone plus GSH plus cystein plus ascorbic
alone plus GSH plus cystein plus ascorbic
Average
Average Average
Average 0-09
0-08 009
o-io
O-OQ
O'O25
O-O25 0-015 0-015 O'O2
these results it seems fairly certain that the diminution of arginase activity during development is not due to a diminution of activators. The results constitute an important argument in favour of the theory of Edlbacher. It would however be desirable to extend them to developing eggs in which the synthesis of nucleoproteins is only partial (cf. Brachet, 1933), such as that of the sea-urchin.
Finally, an attempt was made to estimate the arginase activity of the embryo following more closely the method of Baldwin (1935). Arginase is not normally bound to the cell proteins and its estimation is possible in brei. Unfortunately the results were rather unsatisfactory as the tissue extract after filtration on being made alkaline throws down a slimy mass of semi-denatured protein which makes sampling difficult. It was nevertheless clear that in these tissue extracts the activity of the arginase was of just the same order as in the intact embryos (£)n 0-3-0-4), at \\ days'
incubation age, at all />H's from 7-5 to 9-0. The lack of a marked ^>H optimum corresponds with the findings of Hunter (1934) on arginase from other sources. The action of activators on the arginase of 4.V day embryos in brei was also studied, but found to be absent.
II. UREA AND THE FORMATION OF URIC ACID IN THE EGG.
(1) INTRODUCTION.
No satisfactory explanation exists at the present time of the mechanism of uric acid formation from catabolic nitrogen by the avian organism. All that can be said is that a number of hypotheses have been shown to be untenable. These will be referred to at the end of this section. The suggestion of Wiener (1902), which rather undeservedly became classical, was that a molecule of urea condensed with a three-carbon chain to form dialuric acid, which then condensed with a further molecule of urea to give uric acid. The three-carbon chain was commonly supposed to be tartronic acid, mesoxalic acid, or malonic acid.
COOH COOH COOH
CHOH CO CH,
COOH COOH COOH Tartronic acid Mesoxalic acid Malonic acid
Later Fearon (1932) succeeded in condensing biuret with sarcosine in vitro with the production of small amounts of uric acid.
NH, COOH NH—CO
CO + CH,—NHX -* CO C—NHV
I
XC H , I
J
l > C O
NH—CO—NH, NH—C—NH/ Biuret Sarcosine Uric acid
This condensation, if found to occur in the body, would have the advantage of providing a function for sarcosine, which has long been known to occur widely in the tissues of the vertebrates, but for which no likely function has ever been put forward. Although biuret has never been isolated from tissues, its existence there as an intermediate product in this synthesis might be fleeting.
The following experiments were designed to test these hypotheses with regard to the fate of urea in the hen's egg. Does the urea accumulating during development represent the whole of that which is formed, or only an excess over some further process which removes it? The method of injection of precursor substances into the egg, much used by the Japanese workers of the school of Tomita, was employed. It has the disadvantage in this case that the limits of its applicability are rather narrow. If too small a dose is injected, no marked effect is produced, the normal variation in uric acid formation being considerable, but if the dose is too large, death or severe injury results. Similarly, if the injection is made too soon too many eggs are required for the process to be practicable, and if the injection is made too late the estimation has to be done on allantoic liquids containing a large amount of very insoluble uric acid deposits.
(2) MATERIAL AND METHODS.
Eggs from the same sources as those mentioned in the previous part of the paper were used. Serially numbered and dated, they were incubated at intervals of three days to provide a continuous supply of embryos at the right age.
Injection of precursor substances were made on the 12th day of incubation, which is just before the maximum point of uric acid production and well after the peak of urea production. A sterilised glass hypodermic syringe was used, with a 4-cm. needle, large and stout enough to puncture the shell easily. The solution, usually 0-5 or 1 c.c, was injected into the air space and the hole closed with a drop of paraffin wax. Each egg was carefully sponged with a mercuric chloride solution before injection. In this way the loss by infected eggs was kept down to very low figures.
The eggs were opened for uric acid estimation after three days further in-cubation. This allows sufficient time for the assimilation of the injected material and is early enough (15th day) to avoid the worst accumulation of uric acid deposits as well as any of the complications of hatching. The allantoic liquid was carefully collected, and together with the allantoia themselves ground with sand and washed into a large flask. Frequently small granular masses of yellowish solid uric acid were found in the allantoic cavity. These, which are covered with a protective coat of mucin, require special treatment, according to the work of Fiske and Boyden (1926). They were separated out by hand and let stand for an hour in a few c.c. of concentrated HC1. This was then evaporated off on a steam bath and the uric acid dissolved in the smallest possible amount of Benedict-Hitchcock phosphate solution (Benedict and Hitchcock, 1915). It was then added to the total extract.
Deproteination was effected by boiling at a pH yellow-green to brom-cresol-purple. The precipitate was extracted twice. Uric acid was estimated according to the method of Benedict in a Klett colorimeter.
The compounds injected were:
(1) Ammonium carbonate (B.D.H., A.R.). 10 mg. are equivalent to 8-4 mg. uric acid.
(2) Tartronic acid (B.D.H. special preparation). 10 mg. are equivalent to 14 mg. uric acid.
(3) Urea (B.D.H. recryst.). 10 mg. are equivalent to 8-4 mg. uric acid. (4) Biuret (two specimens, one prepared by W. R. Fearon, the other Kahlbaum). 10 mg. are equivalent to 9-6 mg. uric acid.
(5) Sarcosine (Kahlbaum). 10 mg. are equivalent to 4 mg. uric acid. 10 mg. biuret plus 8-8 mg. sarcosine are equivalent to 13-8 mg. uric acid.
(3) EXPERIMENTAL RESULTS.
The results of the injections, given in Table VI, show a number of points of interest.
Table VI. Injection experiments with uric acid precursors. Injection Controls Ammonium carbonate Urea Tartronic acid
Urea and tartronic acid
Biuret (Fearon)
Biuret (Kahlbaum)
Uiuret and sarcosine
Mg. per egg — — — — — — — — — — 0 9 34 23 1 2 1 2 8 4 1 0 8 1 3 13 1 3 6J 1 3 2 0 1 0 1 2 12 13 9 8 9 8 13 " 1
1 0 1 0 1 0 1 0 1 0 1 0 1 0 71 71 No. Injected — — — -— — — — — — — 6 6 6 6 6 6 5 6 6 1 2 1 2 6 6 6 5 6 7 9 5 5 5 6 9 6 6 6 6 6 5 of eggs Survived 6 6 6 6 6 5 5 5 5 5 5 0 0 0 0 2 5 5 5 5 8 8 4 5 4 0 2 2 0 0 0 0 3 6 6 4 6 4 4 4
Av. wet weight embryos on 15th day gm.
(normal =
11 gm.)
— — — — 9 4 9 4 8 9 — — — — — — — — — — — — — 9 9 8 9 9 9 — — 9 3 — — — — 9'5 8-8 — — IO'I 10-3
1 0 6 9-4 Uric acid mg. per embryo I5-5 18-3 17-8 17-0 14-0 18-7
1 3 9
21-8
ig-8 18-6
1 8 9 — — —
23-8 26-6
1 7 9
26-7 28-6 10-3*
U S *
1 6 6 i 6 -7
17-0
16-8
1 6 5 — — — 23-2 18-1 8-8» 15-8
1 3 6 1 6 3 1 6 6
• Probably some loss owing to insufficient treatment of solid masses.
100-0
100
-o
t
.fi V
i
o
Friderlda (191a! • Silver precipitation
Needh*m (1936) NH4C1 titmtion
FlBke and Boyden (1926) Benedict (col on metric) Tanjonski (1927! Moms and Mcl.eod (rokinmetrn I Kninei (19381 FnUn and Wu (eolorimctric)
Toraita and Tslcahaihi ngjql u olonmetncl CaJvery (1930I • Folln and Marenczy Present worti Benedict |bl.ick dot^l
Tom it and
TakahasI
Days
a colorimetric method instead of the ammonium chloride precipitation method as used in this laboratory in 1926.
From Fig. 3, in which the figures have been represented diagrammatically, it can be seen that the range of normal variation at the 15th day is between 14^ and 20 mg. This range is much exceeded by embryos of eggs into which ammonium carbonate has been injected (24-29 mg.), unless, as in one case, the dose was very small. The first two urea experiments were vitiated by the fact that insufficient treatment was given to the uric acid masses, but later experiments showed a range within the normal. Tartronic acid also gave no increase, and with urea and tartronic
32-
28-" 2 " .
T3
20-.o 16-3
[image:13.451.105.341.217.478.2]i
Fig. 3. Precursors of uric acid.
acid together, it proved impossible to get living embryos. This would not have been expected from the work of Tomita and Takahashi (1929), who after injecting urea and tartronic acid together into the egg obtained increases of more than three times the normal value in their best experiments. As has already been pointed out, however, their normal levels were so low that we cannot have confidence in the view that their increases were outside the limit of normal variation.
sarcosine to give uric acid in the avian organism. More certain is the conclusion that urea itself will not form uric acid.
This is in agreement with the whole trend of work on this subject during the past ten years. The only evidence in favour of the view that urea is a precursor of uric acid in the bird derives from the old work of Meyer and Jaffe (1877) and of Wiener (1902), the technique of which cannot be accepted now as adequate. After the classical liver perfusions of Kovalevski and Salaskin (1901), who showed that uric acid could be formed from ammonium lactate, Izar (1911) reported a similar synthesis by perfusion methods from urea, but this claim was not as convincing as the negative results of Friedmann and Mandel in 1908. In recent times the subject has been exhaustively studied by workers of the Italian school. Urea has been given to birds (hens, geese, pigeons) by injection, at the same time as the adminis-tration of the following substances by the mouth, and in no case was there any significant increase in the quantity of uric acid excreted:
Malonic, tartronic and lactic acids (Clementi, 1929). Pyruvic and propionic acids (Torrisi and Torrisi, 1931). Dialuric acid (Torrisi, 1932).
Glycerol and glyceric acid (Biondi, 1932).
Tartronic, lactic, barbituric and dialuric acids (Russo, 1934).
Glycerol, malonic, tartronic and lactic acids (Schuler and Reindel, 1933). In some of these experiments the urea was quantitatively recovered. Entirely similar results were obtained by Benzinger and Krebs (1933) using tissue-slices of avian liver, and by Russo and Cuscuna (1931) and Cuscuna (1934) using the method of liver perfusion.
On the other hand, the formation of uric acid from ammonium carbonate, first demonstrated by Minkovski in 1886, has been repeatedly confirmed, e.g. by Clementi (1930) and Russo (1933), and is seen again in the present experiments.
III. SUMMARY.
1. The urea formed by the chick embryo during its development is not derived from the ammonia of general protein catabolism, either directly or by way of the ornithine cycle.
2. Nor is it derived from uric acid by a complex uricase similar to that of fishes and Amphibia (Stransky's complex: uricase, allantoinase and allantoicase). The uricase discovered in the chick embryo by Przylecki probably breaks down uric acid only as far as allantoin.
3. The urea of the chick embryo is entirely derived from an active arginine-arginase system present at all stages examined from the second day of incubation onward.
diminution in the relative amount of the enzyme. It forms an important support for the view of Edlbacher, hitherto mainly based on observations of tumour tissues, that arginase is an enzyme associated with high growth rate and probably nuclein synthesis, as well as with the production of urea as a vehicle of waste nitrogen.
5. Arginase is also contained in the yolk-sac, but in much smaller quantity than in the embryo. Here also the low activity is not due to the absence of activators. 6. Experiments made to test the possibility that the urea, once formed, might, as would be the case on Wiener's theory, participate in the formation of uric acid, took the form of injection of possible precursors into the egg on the 12th day of development and estimation of the uric acid in the egg on the 15th. In this way it is shown that urea cannot be converted into uric acid by the chick embryo. This conclusion is in agreement with much recent work on the adult bird.
7. A preliminary test was also made of the possibility that biuret and sarcosine could be condensed to uric acid by the chick embryo. The results were negative.
ACKNOWLEDGMENTS.
The authors wish to express their thanks to the Government Grant Committee of the Royal Society for a grant which partially defrayed the cost of these researches. One of us (J. B.) is indebted to the Administrative Council of the University of Brussels for a Stipend during the tenure of which the present work was carried out. Another (R. K. B.) is indebted to Harvard University for a travelling Fellowship.
REFERENCES.
AMMON, R. and SCHOTTE, E. (1935). Biochem. Z. 275, 216.
BALDWIN, E. (1935). Biochem. J. 29, 252.
BENEDICT, S. R. (1935). In Cole: Practical Phyiiological Chemistry. Cambridge 1935. BENEDICT, S. R. and HITCHCOCK, E. H. (1915). J. biol. Chem. 20, 673.
BENZINGER, T. and KREBS, H. A. (1933). Klin. Wschr. 12, 1206. BIERICH, R. and ROSENBOHM, A. (1935). Z. phyt. Chem. 231, 47. BIONDI, A. (1932). Arch. Fisiol. 30, 174.
BRACHET, J. (1933). Arch. Biol. 44, 519. BYERLY, T. C. (1932). J. exp. Biol. 9, 15. CAI.VERY, H. (). (1930). J. biol. Chem. 87, 691. CHAUDHURI, A. C. (1927). J. exp. Biol. 5, 97.
CI.EMENTI, A. (1929). Boll. Soc. ital. Biol. sper. 4, 9, 508, 1062. (1930). Arch. Sci. Biol. 14, 1.
(1932). Proc. XlVth Int. Physiol. Congr. Rome, p. 54. CUSCUNA, T. (1934). Arch. Fisiol. 33, 402.
DICKENS, F. and GREVILLE, G. D. (1933). Biochem. J. 27, 1123.
EDLBACHER, S. (1934). Rektoratsrede, Basel Univ.
EDLBACHER, S. and JUNG, A. (1934). Z. phys. Chem. 227, 114.
EDLBACHER, S., ROLLER, F. and BECKER, M. (1934). Z. phys. Chem. 227, 99. EDLBACHER, S., KRAUS, J. and LEUTHARDT, F. (1934). Z. phys. Chem. 217, 89.
EDLBACHER, S. and KUTSCHER, W. (1931). Z. phys. Chem. 199, 200. (1932). Z. phys. Chem. 207, 1.
EDLBACHER, S. and MERZ, K. W. (1927). Z. phyt. Chem. 171, 253. EDLBACHER, S. and ROTHLER, H. (1925). Z. phys. Chem. 1*8, 264. FEARON, W. R. (1932). Private communication.
F18KE, C. H. and BOYDEN, E. A. (1926). J. biol. Chem. 70, 535. FRIDERICIA, L. S. (1912). Skand. Arch. Physiol. 26, 1.
FRIEDMANN, M. and MANDEL (1908). Arch. exp. Path. Pharmak. (Schmiedeberg-Festschr.), p. 199. GLOVER, E. C. (1931). C. R. Soc. Biol., Paris, 107, 1603.
HUNTER, A. (1934). Quart. J. exp. Physiol. 24, 177. IZAR, G. (1911). Z. phys. Chem. 73, 3:7.
KAIEDA, J. (1930). Z. phyt. Chem. 187, 207. KAMEI, T. (1928). Z.phys. Chem. 171, 101.
KARRER, P. and ZEHENDER, F. (1934). Helv. Chem. Ada, 17, 737. KLEIN, G. and ZIESE, W. (1934). Z. phys. Chem. 229, 209. KOVALEVSKI, K. and SALASKIN, S. (1901). Z. phys. Chem. 33, 210. KREBS, H. A. and HENSELEIT, K. (1932). Z. phys. Chem. 210, 33. LEUTHARDT, F. and KOLLER, F. (1934). Helv. Chem. Ada, 17, 1030.
LIEBEN, F. and LIEBER, H. (1934). Biochem. Z. 275, 38.
MEYER, H. and JAFFE, M. (1877). Ber. dtsch. chem. Ges. 10, 1930, 1963. MINKOVBKI, M. (1886). Arch. exp. Path. Pharmak. 21, 41.
MURRAY, H. A. (1926). J. gen. Physiol. 9, 621. NEEDHAM, J. (1926). J. exp. Biol. 3, 189, and 4, 114.
(1931). Chemical Embryology. Cambridge 1931. (i934)- Biol. Rev. 9, 79.
PRZYLECKI, S. J. and ROGALSKI, L. (1928). Arch. Intern. Physiol. 29, 423.
PURR, A. and WEIL, L. (1934). Biochem. J. 28, 740.
Russo, G. (1933). Boll. Soc. ital. Biol. sper. 8, 123. (1934). Arch. Fisiol. 33, 124.
Russo, G. and CUSCUNA, T. (1931). Boll. Soc. ital. Biol. sper. 6, 250. SALASKIN, S. and SOLOWJEW, L. (1931). Z. phys. Chem. 200, 259. SCHULER, W. and REINDEL, W. (1933). Z. phys. Chem. 221, 209. STRANBKY, E. (1933). Biochem. Z. 266, 287.
TAKAHASHI, M. (1928). Z. phys. Chem. 178, 294. TARGONSKI, H. (1927). Bull. Ac. Pol. Sci. Lett. 1277. TEI8SIER, G. (1931). Trav. Sta. biol. Roscoff, 9, 27.
TOMITA, M. and TAKAHASHI, M. (1929). Z. phys. Chem. 184, 272. TORRISI, D. (1932). Boll. Soc. ital. Biol. sper. 7, 42.
TORRISI, D. and TORRISI, F. (1931). Boll. Soc. ital. Biol. sper. 6, 262.
TRUSZKOWSKI and CZOTERSKI (1933). Biochem. J. 27, 66. WALDSCHMIDT-LEITZ, E. (193I). Naturwissenschaften, p. 964.