Spatial Dynamics in Model Plant Communities:
What Do We Really Know?
Benjamin M. Bolker,1,*Stephen W. Pacala,2 and Claudia Neuhauser3
1. Zoology Department, University of Florida, Gainesville, Florida 32611-8525;
2. Department of Ecology and Evolutionary Biology, Princeton University, Princeton, New Jersey 08544-1003;
3. Department of Ecology, Evolution, and Behavior, University of Minnesota, St. Paul, Minnesota 55108
Submitted March 14, 2002; Accepted December 20, 2002; Electronically published July 21, 2003
abstract:A variety of models have shown that spatial dynamics and small-scale endogenous heterogeneity (e.g., forest gaps or local resource depletion zones) can change the rate and outcome of com-petition in communities of plants or other sessile organisms. How-ever, the theory appears complicated and hard to connect to real systems. We synthesize results from three different kinds of models: interacting particle systems, moment equations for spatial point pro-cesses, and metapopulation or patch models. Studies using all three frameworks agree that spatial dynamics need not enhance coexistence nor slow down dynamics; their effects depend on the underlying competitive interactions in the community. When similar species would coexist in a nonspatial habitat, endogenous spatial structure inhibits coexistence and slows dynamics. When a dominant species disperses poorly and the weaker species has higher fecundity or better dispersal, competition-colonization trade-offs enhance coexistence. Even when species have equal dispersal and per-generation fecundity, spatial successional niches where the weaker and faster-growing spe-cies can rapidly exploit ephemeral local resources can enhance co-existence. When interspecific competition is strong, spatial dynamics reduce founder control at large scales and short dispersal becomes advantageous. We describe a series of empirical tests to detect and distinguish among the suggested scenarios.
Keywords: spatial, competition, competition-colonization, succes-sional niche, phalanx, endogenous.
Spatial heterogeneity at many different scales can change the rate and outcome of competitive interactions. In the past decade, attention has focused on the effects of
short-* Corresponding author; e-mail: [email protected].
Am. Nat. 2003. Vol. 162, pp. 135–148.䉷2003 by The University of Chicago.
0003-0147/2003/16202-020110$15.00. All rights reserved.
distance dispersal and small-scale, endogenous resource heterogeneity such as light gaps in forests (Dalling et al. 1998). Empirical and theoretical studies have demon-strated the effects of small-scale endogenous heterogeneity on invasion, productivity, succession, and diversity in communities ranging from bacterial assemblages to forests (Chao and Levin 1981; Tilman 1993, 1994; Pacala and Deutschman 1995; Rees et al. 1996). Given that dispersal and competitive interactions are fundamentally restricted in space, often to scales only a few times the size of a single organism, we might expect short dispersal and small-scale heterogeneity to have nearly universal effects on community dynamics.
The dynamical richness and technical challenge of spatial processes have encouraged the proliferation of mathematical models for endogenous spatial pattern formation (Levins and Culver 1971; Shigesada et al. 1979; Durrett and Levin 1998; Gandhi et al. 1998). However, most of the models have focused attention on a single class of phenomena (clus-tering and competition-colonization trade-offs) at the ex-pense of other possibilities. In addition, studies that attempt any kind of analysis have made strong simplifying assump-tions about the scales and nature of spatial competition and lead to controversy and confusion when different models have conflicting assumptions (Grace 1990).
This article tackles three basic questions: How does en-dogenous spatial structure affect competition and coex-istence in simple competition models? Do these effects differ depending on the way in which space is modeled? How can these theoretical predictions be tested in the field? We find, reassuringly, that a variety of different models all make qualitatively similar predictions, provided that they incorporate the basic building blocks of discrete individ-uals, local competition, and local dispersal (Durrett and Levin 1994).
When interspecific competition is weak relative to in-traspecific competition and species would coexist nonspa-tially (with global dispersal and competition) and when competing species have similar life histories, endogenous spatial structure may tip the community from coexistence to exclusion in the long term (Neuhauser and Pacala 1999;
fig. 2). However, spatial structure can also change the rate of community dynamics and slow competitive exclusion (Neuhauser and Pacala 1999).
Short dispersal can handicap a competitively dominant species and allow competitive subordinates to coexist with a short-dispersing dominant species; this phenomenon is a form of the classical “competition-colonization trade-off” (CC; Tilman 1994; Holmes and Wilson 1998).
Short dispersal can benefit a competitively subordinate species that reproduces faster than the dominant by al-lowing it to concentrate its reproductive effort in areas that are temporarily free from interspecific competition; this strategy has variously been called “competition” (Grime 1977), “exploitation” (Bolker and Pacala 1999), or a “successional niche” (Pacala and Rees 1998). Here, we call it the “spatial successional niche” (SSN), to avoid con-fusion with temporal successional niches that are main-tained by disturbance.
When interspecific competition is strong relative to in-traspecific competition, short dispersal and small com-petitive neighborhoods benefit individuals by allowing lo-cal control of the environment. In spatial arenas, lolo-cal founder control (dependence of competitive success within a small region on initial density) can still occur, but global founder control disappears; in a large enough habitat, one species will always win as long as both species start with positive densities everywhere in the habitat (Neuhauser and Pacala 1999). The combination of spatial localization and interspecific interactions strongly retards competitive dynamics (Gandhi et al. 1998).
Next, we briefly review the different model frameworks that we and others have used to explore spatial community dynamics. We synthesize recent results from these different frameworks and show that all of them produce similar re-sults that we can classify into the four basic phenomena previously described. We then outline a set of field exper-iments that test for and distinguish among various spatial phenomena that occur in natural communities in the ab-sence of exogenous heterogeneity. (Although exogenous het-erogeneity is obviously important in structuring natural communities, high-diversity communities do exist where no obvious exogenous heterogeneity is acting to maintain di-versity.) Such experiments would fill a major gap because the rich theory of community spatial dynamics remains largely untested by manipulative experiments except in bac-terial communities (Chao and Levin 1981; Korona et al. 1994; Rainey and Travisano 1998).
Review and Synthesis of Spatial Phenomena
Modeling Frameworks
The wide range of simplifying assumptions about the structure of space and the nature of competition used by
different models makes it hard to compare their predic-tions. We have found, however, that several different model frameworks that we and others have explored over the past few years converge independently to the same con-clusions. Interacting particle systems (IPS) are stochastic, continuous-time models with discrete individuals located in the cells of a square lattice; these models often offer qualitative insights into spatial dynamics rather than quan-titative predictions (but see Silvertown et al. 1992 or Law et al. 1997 for counterexamples). Space is discrete rather than continuous in IPS models, but it is contiguous; com-petitive neighborhoods overlap and are connected in an explicitly spatial structure.
Stochastic point processes also track discrete individuals that compete and disperse locally but assume that they occupy a single point in continuous (and, hence, also con-tiguous) space (Gandhi et al. 1998; Bolker and Pacala 1999; Dieckmann and Law 2000; see Diggle 1983 for the theory of static point processes). Stochastic point processes rep-resent a low-density limit of IPS models where very few lattice cells are occupied but competition and dispersal also become long range. We have simulated stochastic point processes and also analyzed them with approximate moment equations that track the mean densities and spa-tial covariances within and between species (Bolker and Pacala 1999).
Finally, in a limit where competition remains local but dispersal is global, we obtain noncontiguous patch models. These models differ from the metapopulation or patch-occupancy models used in many studies (Levins and Cul-ver 1971; Tilman 1993) in that they allow multiple indi-viduals and multiple species per patch. They allow some offspring to remain in the parental sites, which in turn allows spatial pattern to develop in the form of a multi-variate distribution of the fractions of patches with par-ticular combinations of species densities.
Each of these approaches has its own strengths and weaknesses; using all three allows a choice of strengths and lends generality to the results. The IPS framework allows rigorous proof of the long-term coexistence or exclusion of species with different competitive and dispersal abilities in an infinite habitat (Durrett 1992; Durrett and Neuhau-ser 1994; NeuhauNeuhau-ser and Pacala 1999) or approximation by means of pair approximations (Iwasa 2000). Spatial point processes provide the closest link to individual-based field studies and models, with all processes defined in terms of field-measurable life-history and spatial proper-ties such as per capita fecundity and the density of seeds or offspring at a given distance from the parent. In ad-dition, spatial moment equations allow analytical calcu-lations of the shape and scale of equilibrium spatial pat-terns in a community. Finally, patch models provide a model framework that is much simpler to analyze or
sim-ulate than either IPS or moment equations, and they give a point of comparison with popular and even simpler patch-occupancy or metapopulation models (Keeling 2000).
We will discuss results from IPS, point processes, and patch models in order to demonstrate the strengths and weaknesses of each kind of model and to show how all three approaches converge on the same qualitative conclusions.
Model Results: Classes of Phenomena
Nonspatial Foundations.We start with the classical non-spatial Lotka-Volterra (LV) competition model:
n1 n2 ˙n pr 1⫺ ⫺b , 1 1
(
12)
K1 K2 n2 n1 ˙n pr 1⫺ ⫺b , (1) 2 2(
21)
K2 K1whereri(forip1, 2) is the intrinsic exponential growth rate,Kiis the carrying capacity when each species is grown in monoculture, and bij gives the relative strength of within- and between-species competition. (Our definition ofbijgives the competitive effect of a density of speciesj relative to its own carrying capacityKjrather than relative to the target species’ densityKi, which simplifies the no-tation; using the more common notation would replace bij with aijp[Ki/Kj]bij.) The parameters in the model describe whole-population demography rather than in-dividual life history and competitive abilities and as such may be misleading in an evolutionary context; for example, increasing density-independent fecundity or decreasing density-independent mortality would increase bothrand
Krather than justr. However, the model can also be easily derived from an individual-based competition model with parameters fi (fecundity), mi (density-independent mor-tality), and aij (per capita effect of competition; Yodzis 1989; Royama 1992; Bolker and Pacala 1999). The population-level parameterization is simply more conve-nient for describing the criteria for species coexistence.
Although the LV model is a caricature of real compe-tition processes, it provides a useful framework for dis-cussing the possible outcomes of competition (fig. 1). It covers scenarios ranging from simple single-resource com-petition (whenb b p1) to strongly asymmetric
com-12 21
petition, as is commonly incorporated in patch-occupancy models (when b12r⬁, b21r0). The invasion crite-rion—the conditions on life-history, competitive, and spa-tial parameters that allow one species to invade an estab-lished monoculture of another species—for species i to invade speciesjin this model isb !1. With appropriate
ij
definitions, this invasion criterion also applies to a wider range of models, where per capita competition is a possibly nonlinear function of linear combinations of species densities (Chesson 2000b; B. M. Bolker, unpublished manuscript).
The LV model allows any of the possible outcomes of two-species competition: dominance by either species, co-existence of both species, or founder control (exclusion of one species by another depending on initial conditions, usually exclusion of an initially rare species by an initially common one). If we derive the LV equations from an underlying model where individuals compete for a single limiting (but well-mixed) resource, we obtain the same equations, but the plane of possible competitive param-eters shown in figure 1 shrinks to a hyperbola that allows only single-species dominance. Although many processes (e.g., allelopathy or responses to herbivory or pathogens) can move interactions off of the single-resource hyperbola and allow coexistence or founder control in a nonspatial model, we focus on how spatial competition allows co-existence when the underlying nonspatial rules would al-low only one dominant species to persist.
The LV parameter plane also helps organize the different possible effects of spatial pattern formation on competitive dynamics. The possible phenomena, which we discuss in the following sections, are spatial segregation and cluster-ing when the parameters lie in the coexistence region of figure 1 (quadratI), competition-colonization and spatial successional niche dynamics in the single-species domi-nance regions (quadratsII andIV), and phalanx growth in the founder control region (quadratIII).
Before going any further, we should clarify the meaning of “spatial coexistence.” Spatial coexistence occurs when endogenous or exogenous spatial structure allows species to coexist indefinitely (in the absence of demographic sto-chasticity) where they could not if all individuals experi-enced the average conditions in the environment, includ-ing spatially averaged population densities. Real communities can be difficult to homogenize in this way. Conversely, determining the nonspatial outcome of com-petition is simple in a model where we can set the effective density of each species to its spatial mean density (although finding the nonspatial equivalents of patch-occupancy models can be surprisingly difficult). Heuristically, one can think about the nonspatial outcomes as the hypothetical result of competition if all individuals competed and dis-persed globally.
Spatial Segregation and Clustering.The first, simplest com-petitive scenario is where two species could coexist in the nonspatial, homogeneously mixed case (fig. 1, quadratI). Although such coexistence is not possible under compe-tition for a single essential resource, there are many
sit-Figure 1: Invasibility and coexistence regions for the Lotka-Volterra model and the nonlinear (nonspatial) competition model, with regions of interest marked.
uations such as competition for multiple resources or at-tack by host-specific pathogens that lead to coexistence even in a homogeneously mixed environment. In this case, we are interested in whether endogenous spatial structure can prevent (rather than enhance) coexistence and in how it changes the timescale of competitive dynamics.
When interspecific competition is much weaker than intraspecific competition (bothb12andb21are small; fig. 1, regiond), space does not change qualitative outcomes of competition; species that coexist in the absence of space still coexist in the IPS. We can prove coexistence rigorously in the high-density limit, where fecundity is much greater than mortality for both species and there is no empty space left in the habitat (Neuhauser and Pacala 1999).
In contrast, if interspecific competition is nearly as strong as intraspecific competition (b!but≈1) for one species or the other (fig. 1, regionsa1anda2), the com-bination of local competition and discrete individuals ac-tually reduces coexistence in the high-density limit. In the shaded regions, where the stronger competitor would al-most outcompete the weaker in the nonspatial model, the stronger competitor can actually exclude the weaker be-cause of the discreteness of the competitive neighborhood. In this parameter regime, only a low density of the weaker species can survive in the nonspatial model. If the com-petition neighborhood is small enough, fewer than one individual could survive within a competitive neighbor-hood; because individuals are discrete in this model, the weaker species can never establish. Neuhauser and Pacala (1999) prove this phenomenon rigorously for the high-density limit.
Similar phenomena occur in patch models and in the spatial moment equations. In patch models, we restrict movement by reducing the probability of global interpatch movement rather than the size of overlapping neighbor-hoods, but the results are roughly comparable. To explore this effect, we used an approach that uses forward equa-tions to track the entire probability distribution of patch occupancies in an infinite ensemble of patches (Renshaw 1991). Within a patch, population dynamics follow a sto-chastic process that would converge on the LV equations (1) for large patch sizes; deaths occur at a constant density-independent rate, and fecundity decreases linearly accord-ing to the combination of con- and heterospecific popu-lation densities shown in equations (1) down to 0 when . If births are globally distributed among patches
N≥K
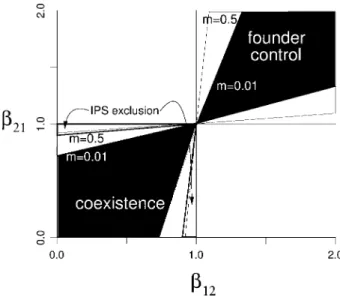
when they occur (mp1), the model converges to the LV model. If we restrict interpatch movement to 50% in a model with patch size (carrying capacity)Kp10, the co-existence region shrinks by approximately 10% (1/K) in the same regions where coexistence fails in an IPS (fig. 2). Reducing interpatch movement further (to 1%) more than doubles this contraction, and it reduces the size of the founder control region (see “Phalanx Strategies”).
In spatial point process models, intraspecific competi-tion is always strengthened by a self-competicompeti-tion term (Bolker and Pacala 1999). An individual-based parame-terization of the LV model starts with the statement that competition with an individual of speciesj at distancer decreases individual fecundity rate or increases individual mortality rate by an amount aijUij(r). (The connection with the population-level parameterization previously given is that K pr/a and b p[a /a][K/K].) In a
i i ii ij ij ii j i
randomly distributed or well-mixed conspecific popula-tion of density Ni, the total strength of competition is , where the second term represents self-aii(Ni⫹Uii[0])
competition that does not occur in the nonspatial model. Self-competition changes competitive outcomes only if it changes bij, which in turn changes only if the ratio of carrying capacitiesK/Kp(r ⫺aU(0))/(r⫺aU(0))is
i j i ii ii j jj jj
different from the original ratio Ki/Kj, which in turn re-quiresU(0)/K (U(0)/K. This criterion depends on the
ii i jj j
relative carrying capacities and the shape of the compet-itive neighborhood. In contrast, the more general neigh-borhood exclusion phenomenon proved for the IPS does not depend on details of spatial neighborhoods (Neuhau-ser and Pacala 1999) but may depend on the high-density limit.
A final spatial phenomenon that does not alter the qual-itative outcome of competition but does change the ap-parent strength of interspecific competition (changing the estimated location of a community in the parameter space shown in fig. 1) is the effect of spatial segregation among coexisting species. In nonspatial models with symmetric
Figure 2:Changes in coexistence regions with limited movement in the metapopulation Lotka-Volterra (LV) model. When all births are dispersed globally (mp1), the coexistence regions are equal to those of the stan-dard LV (fig. 1). When a fraction1⫺mof births are retained locally, the coexistence and founder control regions (calculated by searching numerically for thebvalues where the invasion rate of each species is 0) shrink as shown. The triangles show the equivalent region of con-traction for the interacting particle systems in the high-density limit. Parameters: intrinsic rate of increaser1pr2p3.0, intrinsic reproductive number (competition-free expected offspring per generation) R1p
, carrying capacity .
R2p4.0 K1pK2p10.0
species, increasing interspecific competition (e.g., from in-creasing niche overlap) continuously increases the ob-served strength of competitive interactions between spe-cies. In the equivalent spatial model, however, interspecific spatial segregation also increases with increasing interspe-cific competition and reduces interspeinterspe-cific interaction. Re-moval experiments—observing the increased performance of plants in an established community after the experi-mental removal of conspecific or heterospecific competi-tors—are a standard assay of competitive strength. As we have shown (Pacala and Levin 1998), the endogenous spa-tial segregation generated by competition can lower the estimated strength of competition. For strongly competing, symmetric species (fig. 1, regionb), spatial segregation is so strong that interspecific population dynamics are nearly neutral, as proved in the high-density limit by Neuhauser and Pacala (1999). This phenomenon might explain ob-served differences between quadrat-based and individual-based field estimates of interspecific competition individual-based on removal experiments.
Competition-Colonization Trade-Offs.A more familiar sce-nario is the case where one species is clearly competitively dominant in a nonspatial setting (e.g., in small-plot ex-periments pitting two heterospecific individuals against each other). Strong competitors may be better at pre-empting resources or may have some other advantage such as herbivore or pathogen resistance (Holt et al. 1994). In the LV model and other models that assume both spatial and temporal homogeneity (Tilman 1982), a superior competitor always wins, regardless of handicaps in fe-cundity or dispersal, so any coexistence must be the result of spatial structure. Our conclusions come from a variety of models that all converge to the LV in the appropriate nonspatial limit of large competition neighborhoods or patch sizes, so we can interpret nonspatial parameters such as relative strength of interspecific competition in terms of the LV parameters (fig. 1). In this case, we are looking at quadratsIIandIVof figure 1, where one species dom-inates the other.
One particular form of spatial coexistence is the com-petition-colonization trade-off (CC), where two or more species coexist in a spatial environment because the com-petitively dominant species have poorer colonizing ability: low fecundity, short dispersal, or both. The CC has been studied with a variety of models (IPS, spatial point pro-cesses, patch-occupancy models) and analytical tools (for-mal proofs [Durrett and Swindle 1991], pair approxima-tion [Harada and Iwasa 1994], numerical studies [Holmes and Wilson 1998], and second-order moment equations [Bolker and Pacala 1999]). Competition-colonization trade-offs in patch-occupancy models allow a large num-ber of species to coexist (Tilman 1994; Kinzig et al. 1999),
but most fully spatial analyses have considered only two-species interactions. The basic mechanism underlying all forms of CC is that colonization limitation of the domi-nant species leads to a lower overall (spatial mean) density of the dominant, which in turn allows the inferior species to invade. In contiguous-space models such as IPS and point processes, the dominant’s density decreases because of spatial clustering, which enhances intraspecific com-petition and reduces mean density (Harada and Iwasa 1994; Holmes and Wilson 1998; Bolker and Pacala 1999). The CC is fundamentally spatial but is strongly en-hanced if the dominant species also has low fecundity or high mortality so that its density is low (or it occupies only a small portion of the habitat) even in the absence of competition (Bolker and Pacala 1999). Formally similar offs can also be constructed using nonspatial trade-offs between seed size and number, for example (Adler and Mosquera 2000).
In classical patch-occupancy models (Levins and Culver 1971), fecundity and dispersal ability are confounded in a single measure of nonlocal colonization (the number of offspring available to colonize outside the parent’s site is determined by both the number of offspring produced
and the fraction that disperse beyond the site boundaries). The distinction between low fecundity and short dispersal is important because the two forms of limitation have different implications; fecundity limitation is nonspatial while dispersal limitation is spatial. Fecundity and dis-persal limitation combine to produce colonization, or cruitment limitation, which can be tested by adding re-cruits (seeds or young juveniles) to a site and observing whether population density increases. While the influence of recruitment limitation has been extensively tested in both marine and terrestrial communities (Clark and Ji 1995; Clark et al. 1999; Hubbell et al. 1999; Warner et al. 2000), the effects of dispersal limitation have begun to be studied more recently (Dalling et al. 1998; Ehrlen and Eriksson 2000; Jacquemyn et al. 2001; Verheyen and Hermy 2001; Webb and Peart 2001).
There are biological as well as technical reasons to expect difficulty in separating fecundity from dispersal limitation. Late-successional species that invest heavily in structures for resource competition may, as a result, have fewer re-sources to invest in dispersal structures for their offspring. They are also under less pressure to colonize because their survival does not depend on escaping competition, al-though competition both with kin and with other con-specific neighbors will always give some reasons to dis-perse. They may also choose to produce a small number of large, short-dispersing seeds that have higher germi-nation probabilities and early growth rates (Ezoe 1998; Levin and Muller-Landau 2000). Finally, these species are able to maintain long-term control of their local resource environment and may be playing a “phalanx” strategy against other strong competitors.
Although CCs have been intensively studied by theo-reticians, their influence is actually limited in continuous-space models. In simple models of homogeneous envi-ronments, CCs are limited to a small region of parameter space where strong competitors have extremely low fe-cundity and short dispersal (Bolker and Pacala 1999). When fecundity is realistically high (intrinsic reproductive number or lifetime reproduction in the absence of com-petition greater than two), intraspecific spatial clustering is weak or even negative and leads to evenly spaced pop-ulations and rules out a CC (Bolker and Pacala 1997). Competition-colonization trade-offs are more important in models such as patch-occupancy models that assume some spatial structure or heterogeneity, but even these models require the dominant species to be such a poor colonizer that its patch occupancy never gets high enough to exclude the inferior competitor. We suspect that ex-ogenous heterogeneity and disturbance, which both in-crease the variance in population densities above the level that can be generated by endogenous processes, will en-hance the effects of CC; for example, the apparent CC
identified by Tilman (1988) in Minnesota sand plain com-munities was driven by disturbance at the scale of agri-cultural fields. Whether it is safe to extend the same frame-work we have used for endogenous dynamics to include disturbance and exogenous variability is an open question, although Chesson’s (2000a) framework for spatial coex-istence, which we discuss later, does lump endogenous and exogenous processes.
Spatial Successional Niches. Competition-colonization trade-offs are not the only form of spatial coexistence un-der asymmetric competition; competitively inferior species can also gain from short dispersal. In previous work, we have termed this scenario a “successional niche” (Pacala and Rees 1998) or an “exploitative strategy” (Bolker and Pacala 1999); here, we call it a “spatial successional niche” (SSN). Even when the dominant species does not cluster (which it must do to make a CC work), the inferior species can persist by dispersing its offspring nearby to exploit local, endogenous heterogeneity in the resource environ-ment. The SSN strategists gain the greatest advantage from short dispersal by reproducing quickly to exploit gaps be-fore competitively superior species arrive and take over.
We have found this result in both moment equations for point process models, where it appears as part of the spatial advantage arising from spatial segregation between species (Bolker and Pacala 1999), and IPS. In IPS, the SSN requires that the inferior species reproduce on a fast time-scale and that the empty spaces unfilled by the dominant species percolate—form unbroken corridors across the en-vironment. When these conditions hold, the inferior spe-cies can always find empty space to occupy; the corridors do open and close because of stochastic birth and death in the dominant species, but because of its fast timescale, the inferior species can never be trapped and become ex-tinct. (While it would seem that long-distance dispersal benefits the inferior species in this case, it is true only if dispersal is somehow restricted to corridors and avoids the majority of the habitat that is occupied by the dominant species.)
As with the CC, the SSN encompasses both spatial mechanisms (local dispersal) and nonspatial mechanisms (fast growth; Pacala and Rees 1998). Grime’s (1977) “com-petitive” species exploit a nonspatial successional niche by growing rapidly. Local dispersal and rapid growth reflect different geometries of local growth. New growth of a modular organism can be distributed horizontally, as in clonal swards and grasses, or vertically, as in woody trees. The asymmetric and time-dependent nature of height-structured competition makes vertical growth qualitatively different from horizontal growth and harder to model; there are no analytical models that fully incorporate the trade-offs among vertical growth, horizontal growth, and
dispersal (Pacala et al. 1993; Pacala and Tilman 1994). In contrast to CCs, where nonspatial traits (high fecundity) and spatial traits (good dispersal) co-vary in existing spe-cies, nonspatial aspects of successional niches may go along with spatial CCs. For example, early-successional canopy trees often have fast growth, possibly corresponding to a nonspatial successional niche; long dispersal, correspond-ing to a spatial CC; and high fecundity, which benefits both strategies. There are too many other trade-offs in plant strategies (seed mass and number, germination prob-ability, dispersal prob-ability, etc.) to do a complete accounting or a complete comparative analysis (Rees 1996), but the partitioning of spatial strategies into categories that are robust to differences in model structure is an important first step.
All three of our modeling approaches (IPS, point-process, and patch models) suggest that SSN strategists must have a short generation time relative to their com-petitors. We suspect, therefore, that SSNs will be more important for strongly asymmetric competition between life forms such as grasses and trees rather than for within-guild competition or competition between more similar guilds such as early- and late-successional trees. Although competitive interactions between grasses and trees are of-ten assumed to reflect a trade-off between utilization of water and light, grasses could also persist through a small-scale SSN strategy (and through an interaction with fire frequency in many ecosystems). As in the case of CC, disturbance may also increase the strength of SSN (pre-liminary explorations of patch models suggest that both CC and SSN increase with mild amounts of correlated within-patch disturbance (B. M. Bolker, unpublished man-uscript), but adding either disturbance or other forms of exogenous heterogeneity blurs the distinction between SSN and other forms of spatial coexistence such as tem-poral successional niches.
It may seem hard to distinguish between CC and SSN strategies; among other things, loose semantics allow one to say that any spatial persistence strategy represents a “colonization advantage.” However, there is a simple op-erational test that distinguishes CC from SSN. If a focal species benefits from CC, increasing its dispersal distance will give it minor benefits (because it has good colonization ability already), while increasing a competitor’s dispersal distance will hurt the focal species by eliminating the col-onization advantage. In contrast, increasing the dispersal distance of a focal species that benefits from SSN will hurt it by removing its ability to retain seeds in good areas; in this case, increasing the competitor’s dispersal will have little effect.
Phalanx Strategies.Finally, we come to the scenario where interspecific competition is stronger than intraspecific
competition (either above the single-resource hyperbola in fig. 1 or more strictly in the upper right quadrat, regions
c1 andc2). Here, spatial dynamics affect both the time-scales of community dynamics and the outcome of competition.
Strong interspecific competition leads to strong spatial segregation in this parameter regime, which in turn slows down competitive dynamics. Gandhi et al. (1998) have shown that in a spatial point process starting from random initial conditions, monospecific patches form in the first phase of competitive dynamics as the locally denser species in each neighborhood excludes the sparser species. There-after, the stronger competitor encroaches on patches of the weaker competitor but at a drastically reduced pace that would be equivalent to decades or millennia in most ecological systems.
The qualitative change in competitive dynamics is that unlike in the nonspatial model, founder control never oc-curs; in infinite space and time, the stronger competitor (species 1 in regionc1or species 2 in regionc2 of fig. 1) always wins eventually. Neuhauser and Pacala (1999) rig-orously proved that the founder control region is reduced in the high-density limit model; in a long-range limit, one can show that the founder control region disappears.
From an ecological point of view, however, one may observe either local founder control—dominance of a local region by a weaker competitor that started at higher den-sity—or, at a slightly larger scale, apparent coexis-tence—stability of large monospecific patches over eco-logical timescales. Some authors have concluded on the basis of simulation studies that different species can coexist in this regime (Sole´ and Bascompte 1997; Molofsky et al. 2001), even though competitive exclusion can be proved rigorously for long enough timescales; moment equations and pair approximations also give the wrong result in this regime (strong interspecific competition) because they un-derestimate the effects of large-scale spatial structure (Iwasa et al. 1998). While indefinite coexistence is im-possible in this regime, apparent coexistence over many generations is a real possibility (Frelich et al. 1993).
These timescale effects apply more generally and weaken interspecific competition relative to intraspecific compe-tition whenever strong spatial segregation occurs. Spatial segregation among similar species could amplify the effects of recruitment limitation that have been shown to slow down competitive exclusion in some cases (Hurtt and Pa-cala 1995). In particular, recent theories of the dynamics of neutral communities (Hubbell 2001) could be strength-ened and reconciled with the apparent importance of com-petition on an individual scale, through the effects of small-scale interspecific and interguild spatial segregation. If species within a guild are symmetric and similar in their ability to compete for resources, endogenous spatial
seg-regation could slow dynamics to effective neutrality on ecological timescales (although Chesson and Huntly [1997] argue broadly that mechanisms that simply slow exclusion are not enough to account for diversity). This phenomenon could be tested by analyzing the neighbor-hood densities within a community to see whether inter-specific association (within guilds) is much rarer than ex-pected from a random spatial distribution, although the confounding effects of environmental heterogeneity would have to be taken into account.
We call the scenario of strong spatial segregation in the founder-control region “phalanx growth,” by analogy with the phalanx/guerrilla dichotomy described by observers of growth forms in clonal plants (Lovett-Doust 1981). In nutrient-rich habitats, clonal plants tend to form tightly aggregated patches, whereas in nutrient-poor habitats, they extend their stolons and disperse new ramets farther from existing ones. Spatial moment equations show that this strategy works best when the intrinsic reproductive num-ber (offspring per generation in a noncompetitive envi-ronment) is large (Bolker and Pacala 1999), which cor-responds well with a nutrient-rich environment. This empirical observation suggests that a spatial competition strategy may be an important part of the community dy-namics of competing clonal swards. As we will discuss later in more detail, we could test this prediction by relocating ramets randomly to see whether they gain a competitive advantage by forming patches or whether the patches have more to do with the spatial scale and variability of the resource environment.
Empirical Tests
No matter how clearly it seems to explain the organization of competitive communities, any categorization of spatial strategies stands or falls on its ability to be tested in the field. While recruitment limitation (which could arise from any combination of fecundity, microsite, or dispersal lim-itation) has been both empirically tested and noted in the field (Clark and Ji 1995; Clark et al. 1999; Hubbell et al. 1999), tests of explicitly spatial phenomena are rarer. These tests usually consist of carefully calibrating a spatial model and then using it to contrast the theoretical estimate of the nonspatial behavior of the community with the dy-namics (productivity, species coexistence, etc.) actually ob-served (Pacala and Silander 1987, 1990; Pacala and Deutschman 1995; Rees et al. 1996; Clark et al. 1998). A related approach uses more generic models of competition to extract competition coefficients from observational data on an appropriate scale (Law et al. 1997; Freckleton and Watkinson 2001); these estimates could, in principle, be compared with nonspatial estimates averaging competition over the entire habitat to observe the effects of endogenous
spatial structure on the effective strength of intra- and interspecific competition. We know of only one published example of a manipulative experiment designed to test endogenous spatial mechanisms in plant communities (Stoll and Prati 2001), although there are several such tests of bacterial communities, as previously cited (Chao and Levin 1981; Korona et al. 1994; Rainey and Travisano 1998).
We propose empirical tests to discriminate between dif-ferent mechanisms of spatial coexistence. These range from purely observational tests of field systems to calibration of data-driven models to manipulative experiments. Clearly, there is no magic bullet and no substitute for the natural history and experimental work required to establish the basic processes that operate in a community. We hope, however, that this discussion will clarify what observations are necessary to establish the existence of spatial compe-tition mechanisms and clarify their nature in a particular plant community.
One obvious shortcoming of the tests we propose is that they distinguish only among different endogenous spatial phenomena; they attribute any spatial structure observed to endogenous processes. In experimental systems, one can eliminate exogenous heterogeneity by the usual means of tilling, shade cloth, uniform watering, or fertilization, and so forth, but one can never be absolutely sure that the system has really been homogenized. We feel that exploring endogenous mechanisms is a first step toward understand-ing the full complexity of plant communities, includunderstand-ing both endogenous and exogenous heterogeneity and their interactions.
How can we discriminate among the different spatial scenarios: symmetric species, competition-colonization, SSN, and phalanx growth? We might already know some-thing about the relative strengths of inter- and intraspecific competition and the relative competitive abilities of the two species; then, we could tentatively locate the system on the plot shown in figure 1, which would narrow the possibilities. However, figure 1 is qualitative rather than quantitative, and it would be difficult in practice to reduce the complexity of a real community to a point in the parameter plane. In addition, finding competitive asym-metry between two species would not resolve whether CCs, SSNs, or both were operating. Instead, we suggest a series of qualitative experiments that will discriminate among the different alternatives.
The most straightforward way to test the effects of local dispersal and spatial structure is to relocate individuals so that their local competitors become a random sample of the population. (Note that these experiments are very dif-ferent from classical recruitment limitation experiments, which supply additional seeds or recruits to an area rather than randomizing the spatial patterns of individuals.) All
Table 1:Experiments for discriminating different spatial scenarios Randomize species 1 Randomize species 2 Conclusion 2f 2 0/F Species 2 maintained by CC 1 0/F 1f Species 1 maintained by CC 1f No change Species 1 maintained by SSN No change 2f Species 2 maintained by SSN 1f 2f Phalanx/spatial segregation
Note: Up and down arrows denote increases and decreases, respectively, in fecundity, growth, or survivorship relative to control treatments; 0 denotes no change for one species. CCp
-colonization trade-off; successional niche. competition SNpspatial
of the experiments we suggest are, unfortunately, practi-cally limited to communities of small- or medium-size plants, both by the logistics of transplanting trees and by the timescales required to see an effect, which are on the order of the generation time; forest ecologists will have to continue to calibrate models to determine spatial effects. For logistical reasons, for the ability to distinguish eco-logically distinct individuals (those unconnected by a net-work of underground structure), and in order to avoid transplant shock, it will be easiest to do these experiments with plants that reproduce primarily from seed. Annual plants would be best of all; everything that we describe in terms of transplants could be done by much easier seed manipulation. Randomizing the positions of all individuals is a test of spatial coexistence itself and reduces the system to its nonspatial equivalent by forcing all individuals to compete equally (or at least randomly) with all other in-dividuals. The control treatment is to swap the positions of conspecifics randomly, which will mimic the effects of the randomization treatment but preserve the hetero- and conspecific neighborhood densities of all individuals.
If randomization leads to a significant change in den-sities or population persistence, we would then want to see which spatial mechanism is operating. We modify the previous experiment by randomizing the position of only one species at a time (always controlling for transplant effects by digging up and replanting the “sedentary” spe-cies in the same positions or randomly interchanging the positions of sedentary individuals; table 1).
If the focal species benefits from randomization and increases in relative abundance or drives the other species to extinction, we conclude that it is naturally colonization limited. If the focal species is hurt by randomization and decreases in relative abundance or becomes extinct, we conclude that it exploits endogenous spatial pattern in the environment to survive. If one species clearly dominates in the control treatment and the other exploits spatial pattern, then we conclude that it occupies a SSN. If neither species dominates and both are hurt by randomization, we can infer a phalanx scenario.
If neither species benefits consistently from the
exper-imental treatment but the timescale of competitive dy-namics accelerates significantly, we infer that the system is symmetric and experiences either spatial segregation of coexisting species or phalanx competition. In the spatial segregation case, randomization will not change the co-existence or diversity of the community, but it will increase resilience or the speed at which the community recovers from a change in relative population densities. In the pha-lanx case, randomization will increase the speed of ex-tinction and possibly induce founder control (as in Chao and Levin’s [1981] experiments), although the timescales of plant communities may be too slow to see this effect. In this case, if seeds or seedlings (preferably grown in a common garden) are available, we can assemble com-munities from scratch by planting or transplanting indi-viduals of different species in predetermined random or structured configurations. These assembled communities distinguish between clustering in the coexistence region and clustering in the “founder control” or phalanx region of figure 1. These cases are hard to separate by nonma-nipulative methods because spatial segregation slows the rate of population dynamics severely. In the coexistence region, single individuals or small clusters of an invading species planted within a monoculture of a resident species should increase their local density and/or expand over the course of a few seasons; in the phalanx region, they should disappear as they are overwhelmed by the greater local density of the resident. If interspecific dynamics are too close to neutrality, it could still be impossible to tell co-existence from exclusion, but this protocol at least offers a way to separate individual-level neutral dynamics from the reduced interspecific interactions caused by spatial segregation.
Stoll and Prati (2001) sowed herbaceous plants in ran-dom and “intraspecifically aggregated” mixtures and mea-sured biomass and reproductive output; they found that stronger competitors performed worse and weaker com-petitors performed better in the aggregated treatment. In our terminology, this experiment shows the results of both CC (reduced performance of stronger competitors) and SSN (increased performance of weaker competitors; if the
overall neighborhood density is maintained, intraspecific aggregation implies interspecific segregation).
A completely different way to test and measure the strength of different spatial scenarios is to calibrate a phe-nomenological or mechanistic individual-based model of the plant community by measuring local dispersal and neighborhood competition strengths. Ideally, such a model should also be tested, either against independent obser-vational data or (even better) by using it to predict the outcome of controlled experiments (see van den Bosch et al. 1988 for an example from plant epidemiology). Once the model is calibrated and tested, one can run any ex-periment in the model, including making competition or dispersal global for one or both species, and can measure the effect of endogenous spatial structure on the persis-tence or invasibility of different species. Such a model can test and discriminate among any of the scenarios listed. Completely field-calibrated models have strong advantages but require a great deal of effort to parameterize and test; only a few models of this sort have actually been con-structed and used to test spatial dynamics (Cain et al. 1991; Pacala et al. 1993; Rees et al. 1996).
These experiments will require considerable investment and, as we previously discussed, are practically limited to small plants that reproduce from seed and mature and reproduce quickly. These specifications obviously rule out many important natural communities (forests, old-field communities dominated by long-lived perennials such as
Solidago, etc.), but they are a starting point. We can always fall back on carefully calibrated models for those com-munities that resist experimental manipulation, but we should start with systems where we can do qualitative, manipulative experiments that are independent of specific models, and, if we do use models, we can check them against independent experimental (not just observational) evidence. Given the potential importance of spatial dy-namics and given the near complete absence of experi-mental tests, experiments such as the ones we have sug-gested should be well worth the effort.
Discussion
We believe that our work and the work of many other mathematicians and mathematical ecologists over the past few years have finally made it possible to understand the basic spatial phenomena that underlie both simple stra-tegic models and more complicated models used to un-derstand applied ecological issues such as the generation and conservation of biodiversity. The results presented here suggest that we can understand many of the basic phenomena that underlie spatial competition equally well in patch, lattice, or point-process models. This is good news; in particular, it strengthens the case that the
con-clusions of the voluminous literature on patch dynamics could carry over, qualitatively, to landscapes that are not inherently divided into discrete patches. Models with ex-plicit neighborhood structure such as lattices or point pro-cesses will still be needed to make quantitative or explicitly spatial predictions (such as exploring spatial scales and patterns of dispersal) and measure the relative strengths of different phenomena, but we can hope that patch mod-els will suffice for elucidating basic spatial processes.
While we have intentionally titled this article “Spatial Dynamics in Model Plant Communities,” it is reasonable to ask how important we think the various mechanisms outlined and endogenous spatial structure in general will be in real communities. We argued in the introduction to this article that short scales of dispersal and competition are ubiquitous; conversely, so are exogenous disturbance and heterogeneity, which might be expected to swamp endogenous effects. Some important natural and experi-mental systems such as the Cedar Creek Long Term Eco-logical Research site or the Rothamstead Park Grass ex-periment apparently support high diversity with minimal exogenous heterogeneity. However, these represent only a few (perhaps special) systems, and one can never be sure that exogenous heterogeneity is really absent; color poly-morphisms inLinanthus parryae, long cited as an example of endogenous pattern in population genetics but now believed to be maintained by subtle exogenous heteroge-neity, are but one cautionary example (Schemske and Bier-zychudek 2001).
More evidence for the importance of some kind of spa-tiotemporal mechanism, either endogenous or exogenous, comes from reviews of plant competition experiments and observations (Goldberg and Barton 1992; Gurevitch et al. 1992; Freckleton and Watkinson 2001). These reviews sug-gest that, in general, intraspecific competition is not much weaker than interspecific competition (as would be ex-pected from models of resource partitioning), which rules out regionIdin figure 1 but does not necessarily rule out regionsIaandIb. (Then again, Rees et al. [1996] did find that interspecific competition was relatively weak.) These reviews did not generally try to separate systems on the basis of competitive symmetry (quadrats II and IV vs. quadratsIandIIIin fig. 1); however, asymmetry is at least anecdotally common in natural communities (two ex-amples are the communities studied by Law et al. [1997] and Tilman [1988]). We do not know, in general, the likelihood of competitive near equivalence (b12 or b21≈ ), which would put species pairs near the boundary of 1
quadratI, where strong segregation can occur and where small spatial effects could have important qualitative dif-ferences. (Hubbell [2001] argues for equivalence or near equivalence and bases his argument on macroscopic pat-terns of diversity rather than on competition experiments.)
As for founder control (quadratIIIin fig. 1), we know of no direct evidence for this scenario in plant communities (even in clonal plant communities where one might expect little to no intraspecific competition between ramets), al-though it has been found in bacterial communities (Chao and Levin 1981). Despite all the effort that has been put into field studies of competition, we still cannot make strong a priori predictions about what kinds of endoge-nous effects we expect.
Caveats
The models previously discussed are simple “strategic” or “toy” models (Nisbet and Gurney 1982). As such, they neglect many important complexities such as exogenous spatial and temporal heterogeneity (including disturbance and habitat fragmentation) and nonlinear per capita effects of competition (Chesson 2000a, 2000b), which are certain to shape natural competitive communities. How can we be sure that these phenomena do not somehow reverse our conclusions or that we have not missed some impor-tant strategic spatial axis?
We have explored nonlinear patch models (like the patch models previously discussed but with nonlinear per capita competitive effects), with and without correlated disturbance that drives an entire patch population extinct simultaneously. In the absence of disturbance, SSN strat-egies still dominate CCs over much of parameter space. Alternatively, disturbance, which makes the model more similar to patch-occupancy models by emptying entire patches simultaneously, strengthens the effect of CCs (B. M. Bolker, unpublished manuscript). The only novel phe-nomenon seen in these nonlinear disturbance models is the effect of different curvatures in the effect of compe-tition, which allows one species to do better in rare “gaps” while the other does better in common, medium-density patches (relative nonlinearity; Chesson 2000b).
Peter Chesson (1984, 1985, 1990, 1994, 2000a, 2000b) has extensively explored and categorized the (largely ex-ogenous) effects of spatial and temporal heterogeneity, and their interactions, in patch models. While Chesson’s results may not carry over to contiguous-space models un-changed, the framework is sufficiently general that we can expect that many of the phenomena will be the same. Chesson’s scheme divides spatiotemporal mechanisms into three general categories: relative nonlinearity, which allows some species to capitalize on rare, high-quality sites while others exploit average sites; temporal or spatial storage effects, where carryover from good sites and time periods allows an inferior competitor to persist; and density-growth correlation, where a species manages to inhabit good sites and time periods preferentially, either actively (through directed dispersal) or passively (by retaining
propagules in good habitats). In terms of this framework, the SSN (and interspecific spatial segregation generally) represents a positive density-growth covariance for the in-ferior species; by reproducing on a fast timescale and re-taining its propagules locally, a plant species induces a positive association between population density and good (enemy-free) habitat. In contrast, the CC represents neg-ative density-growth covariance; in intraspecific compe-tition, more individuals are necessarily found in bad (high-density) neighborhoods. We have found that in the presence of exogenous heterogeneity, short dispersal adds an additional component to the density-growth correlation (B. M. Bolker, unpublished manuscript). Chesson’s results suggest that nonlinearity, disturbance, and heterogeneity will add new spatiotemporal mechanisms but not quali-tatively change the ones discussed here.
In contrast to disturbances and heterogeneity, which can be understood at the level of a patch, the effects of frag-mentation—a particular form of exogenous heterogeneity that partially or completely cuts patches off from colo-nization—are explicitly spatial and, thus, harder to un-derstand. They are captured in some ways by our models when movement rates drop very low in patch models—for example—but a full exploration of fragmentation probably requires explicitly spatial models (Bascompte and Sole´ 1997; Fahrig 1998).
Conclusions
We are strongly encouraged by the convergence of three structurally different types of models—IPS, spatial point processes, and patch models—on the same qualitative con-clusions. This convergence strengthens our conclusions, and it suggests, as one would hope, that the fine details of mathematical models do not affect qualitative predic-tions. However, there are two strong caveats to this state-ment. First, quantitative predictions of competitive out-comes in natural communities definitely will depend on choosing an appropriate structure and spatial scale with which to model the system. Second, there are important details as well as unimportant ones (“Everything should be made as simple as possible but not simpler”; Albert Einstein); as we stated at the outset, the properties of in-dividual discreteness and local competition and dispersal are vital. In addition, models that describe only site oc-cupancy or assume that sites are always filled at some stage in the annual cycle may miss important aspects of spatial dynamics, although increasing the number of different types of patches to allow temporary within-patch coex-istence (Pacala and Rees 1998) or multiple resource levels (Wilson et al. 1999) is one way around this problem.
We have shown that despite the potential richness of spatial dynamics, it is possible to generalize about the
ef-fects of endogenous spatial structure on competitive out-comes. Spatial effects are context dependent; whether space enhances or undermines coexistence and whether it speeds up or slows down the rate at which communities approach equilibrium varies according to the degree of asymmetry (in fecundity, mortality, size, etc.) and the po-sition in the competitive plane (fig. 1). More to the point, this variability and context dependence is measurable; we have described a series of experiments that could allow empiricists to distinguish among different spatial scenar-ios. We believe that experiments along the lines we have suggested may help in the ongoing effort to establish which mechanisms are actually most important in maintaining diversity in natural communities.
Acknowledgments
C.N. was partially supported by National Science Foun-dation (NSF) grant DMS-97-03694, and S.W.P. and B.M.B. were partially supported by NSF applied math grant 9807755. We thank P. Chesson, R. Snyder, and four anon-ymous reviewers for helpful comments.
Literature Cited
Adler, F. R., and J. Mosquera. 2000. Is space necessary? interference competition and limits to biodiversity. Ecology 81:3226–3232.
Bascompte, J., and R. V. Sole´. 1997. Models of habitat fragmentation. Pages 127–150in J. Bascompte and R. V. Sole´, eds. Modeling spatiotemporal dynamics in ecol-ogy. Springer, New York.
Bolker, B. M., and S. W. Pacala. 1997. Using moment equations to understand stochastically driven spatial pattern formation in ecological systems. Theoretical Population Biology 52:179–197.
———. 1999. Spatial moment equations for plant com-petition: understanding spatial strategies and the ad-vantages of short dispersal. American Naturalist 153: 575–602.
Cain, M., S. W. Pacala, and J. Silander, Jr. 1991. Stochastic simulation of clonal growth in the tall goldenrod, Sol-idago altissima. Oecologia (Berlin) 88:477–485. Chao, L., and B. R. Levin. 1981. Structured habitats and
the evolution of anticompetitor toxins in bacteria. Pro-ceedings of the National Academy of Sciences of the USA 78:6324–6328.
Chesson, P., and N. Huntly. 1997. The roles of harsh and fluctuating conditions in the dynamics of ecological communities. American Naturalist 150:519–553. Chesson, P. L. 1984. The storage effect in stochastic
pop-ulation models. Lecture Notes in Biomathematics 54: 76–89.
———. 1985. Coexistence of competitors in spatially and
temporally varying environments: a look at the com-bined effects of different sorts of variability. Theoretical Population Biology 28:263–287.
———. 1990. Geometry, heterogeneity and competition in variable environments. Philosophical Transactions of the Royal Society of London B, Biological Sciences 330: 165–173.
———. 1994. Multispecies competition in variable en-vironments. Theoretical Population Biology 45: 227–276.
———. 2000a. General theory of competitive coexistence in spatially varying environments. Theoretical Popula-tion Biology 58:211–237.
———. 2000b. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics 31: 343–366.
Clark, J. S., and Y. Ji. 1995. Fecundity and dispersal in plant populations: implications for structure and di-versity. American Naturalist 146:72–111.
Clark, J. S., E. Macklin, and L. Wood. 1998. Stages and spatial scales of recruitment limitation in southern Ap-palachian forests. Ecological Monographs 68:213–235. Clark, J. S., B. Beckage, P. Camill, B. Cleveland, J.
HilleRisLambers, J. Lighter, J. McLachlan, J. Mohan, and P. Wyckoff. 1999. Interpreting recruitment limitation in forests. American Journal of Botany 86:1–16.
Dalling, J. W., S. P. Hubbell, and K. Silvera. 1998. Seed dispersal, seedling establishment and gap partitioning among tropical pioneer trees. Journal of Ecology 86: 674–689.
Dieckmann, U., and R. Law. 2000. Relaxation projections and the method of moments. Pages 412–455 in U. Dieckmann, R. Law, and J. A. J. Metz, eds. The geometry of ecological interactions: simplifying spatial complexity. Cambridge Studies in Adaptive Dynamics. Cambridge University Press, Cambridge.
Diggle, P. 1983. Statistical analysis of spatial point patterns. Academic Press, New York.
Durrett, R. 1992. Multicolor particle systems with large threshold and range. Journal of Theoretical Probability 5:127–152.
Durrett, R., and S. A. Levin. 1994. The importance of being discrete (and spatial). Theoretical Population Biology 46:363–394.
———. 1998. Spatial aspects of interspecific competition. Theoretical Population Biology 53:30–43.
Durrett, R., and C. Neuhauser. 1994. Particle systems and reaction-diffusion equations. Annals of Probability 22: 289–333.
Durrett, R., and G. Swindle. 1991. Are there bushes in a forest? Stochastic Processes and Their Applications 37: 19–31.
Ehrlen, J., and O. Eriksson. 2000. Dispersal limitation and patch occupancy in forest herbs. Ecology 81:1667–1674. Ezoe, H. 1998. Optimal dispersal range and seed size in a stable environment. Journal of Theoretical Biology 190: 287–293.
Fahrig, L. 1998. When does fragmentation of breeding habitat affect population survival? Ecological Modelling 105:273–292.
Freckleton, R. P., and A. R. Watkinson. 2001. Nonmanip-ulative determination of plant community dynamics. Trends in Ecology & Evolution 16:301–307.
Frelich, L. E., R. R. Calcote, and M. B. Davis. 1993. Patch formation and maintenance in an old-growth hemlock-hardwood forest. Ecology 74:513–527.
Gandhi, A., S. Levin, and S. Orszag. 1998. “Critical slowing down” in time-to-extinction: an example of critical phe-nomena in ecology. Journal of Theoretical Biology 192: 363–376.
Goldberg, D. E., and A. M. Barton. 1992. Patterns and consequences of interspecific competition in natural communities: a review of field experiments with plants. American Naturalist 139:771–801.
Grace, J. B. 1990. On the relationship between plant traits and competitive ability. Pages 51–65inJ. B. Grace and D. Tilman, eds. Perspectives on plant competition. Ac-ademic Press, San Diego, Calif.
Grime, J. P. 1977. Evidence for the existence of three pri-mary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist 111: 1169–1194.
Gurevitch, J., L. L. Morrow, A. Wallace, and J. S. Walsh. 1992. A meta-analysis of competition in field experi-ments. American Naturalist 140:539–572.
Harada, Y., and Y. Iwasa. 1994. Lattice population dynam-ics for plants with dispersing seeds and vegetative prop-agation. Researches on Population Ecology (Kyoto) 36: 237–249.
Holmes, E., and H. Wilson. 1998. Running from trouble: long-distance dispersal and the competitive coexistence of inferior species. American Naturalist 151:578–586. Holt, R., J. Grover, and D. Tilman. 1994. Simple rules for
interspecific dominance in systems with exploitative and apparent competition. American Naturalist 144: 741–771.
Hubbell, S. P. 2001. The unified neutral theory of bio-diversity and biogeography. Monographs in Population Biology. Princeton University Press, Princeton, N.J. Hubbell, S. P., R. B. Foster, S. T. O’Brien, K. E. Harms,
R. Condit, B. Wechsler, S.-J. Wright, and S.-L. De Lao. 1999. Light-gap disturbances, recruitment limitation, and tree diversity in a Neotropical forest. Science (Wash-ington, D.C.) 283:554–557.
Hurtt, G. C., and S. W. Pacala. 1995. The consequences
of recruitment limitation: reconciling chance, history and competitive differences between plants. Journal of Theoretical Biology 176:1–12.
Iwasa, Y. 2000. Lattice models and pair approximation in ecology. Pages 227–251inU. Dieckmann, R. Law, and J. A. J. Metz, eds. The geometry of ecological interac-tions: simplifying spatial complexity. Cambridge Studies in Adaptive Dynamics. Cambridge University Press, Cambridge.
Iwasa, Y., M. Nakamaru, and S. A. Levin. 1998. Allelopathy of bacteria in a lattice population: competition between colicin-sensitive and colicin-producing strains. Evolu-tionary Ecology 12:785–802.
Jacquemyn, H., J. Butaye, and M. Hermy. 2001. Forest plant species richness in small, fragmented mixed de-ciduous forest patches: the role of area, time and dis-persal limitation. Journal of Biogeography 28:801–812. Keeling, M. J. 2000. Metapopulation moments: coupling, stochasticity and persistence. Journal of Animal Ecology 69:725–736.
Kinzig, A. P., S. A. Levin, J. Dushoff, and S. W. Pacala. 1999. Limiting similarity, species packing, and system stability for hierarchical competition-colonization mod-els. American Naturalist 153:371–383.
Korona, R., C. H. Nakatsu, L. J. Forney, and R. E. Lenski. 1994. Evidence for multiple adaptive peaks from pop-ulations of bacteria evolving in a structured habitat. Proceedings of the National Academy of Sciences of the USA 91:9037–9041.
Law, R., T. Herben, and U. Dieckmann. 1997. Non-manipulative estimates of competition coefficients in a montane grassland community. Journal of Ecology 85: 505–517.
Levin, S. A., and H. C. Muller-Landau. 2000. The evolution of dispersal and seed size in plant communities. Evo-lutionary Ecology Research 2:409–435.
Levins, R., and D. Culver. 1971. Regional coexistence of species and competition between rare species. Proceed-ings of the National Academy of Sciences of the USA 6:1246–1248.
Lovett-Doust, L. 1981. Population dynamics and local spe-cialization in a clonal perennial (Ranunculus repens). I. The dynamics of ramets in contrasting habitats. Journal of Ecology 69:743–755.
Molofsky, J., J. D. Bever, and J. Antonovics. 2001. Coex-istence under positive frequency dependence. Proceed-ings of the Royal Society of London B, Biological Sci-ences 268:273–277.
Neuhauser, C., and S. W. Pacala. 1999. An explicitly spatial version of the Lotka-Volterra model with interspe-cific competition. Annals of Applied Probability 9: 1226–1259.
Nisbet, R., and W. Gurney. 1982. Modelling fluctuating populations. Wiley, New York.
Pacala, S. W., and D. H. Deutschman. 1995. Details that matter: the spatial distribution of individual trees main-tains forest ecosystem function. Oikos 74:357–365. Pacala, S. W., and S. A. Levin. 1998. Biologically generated
spatial pattern and the coexistence of competing species. Pages 204–232inD. Tilman and P. Kareiva, eds. Spatial ecology: the role of space in population dynamics and interspecific interactions. Princeton University Press, Princeton, N.J.
Pacala, S. W., and M. Rees. 1998. Field experiments that test alternative hypotheses explaining successional di-versity. American Naturalist 152:729–737.
Pacala, S. W., and J. A. Silander, Jr. 1987. Neighborhood interference among velvet leaf,Abutilon theophrasti, and pigweed,Amaranthus retroflexus. Oikos 48:217–224. ———. 1990. Field tests of neighborhood population
dy-namic models of two annual weed species. Ecological Monographs 60:113–134.
Pacala, S. W., and D. Tilman. 1994. Limiting similarity in mechanistic and spatial models of plant competition in heterogeneous environments. American Naturalist 143: 222–257.
Pacala, S. W., C. D. Canham, and J. A. Silander, Jr. 1993. Forest models defined by field measurements. I. The design of a northeastern forest simulator. Canadian Journal of Forest Research 23:1980–1988.
Rainey, P. B., and M. Travisano. 1998. Adaptive radiation in a heterogeneous environment. Nature 394:69–72. Rees, M. 1996. Evolutionary ecology of seed dormancy
and seed size. Philosophical Transactions of the Royal Society of London B, Biological Sciences 351:1299–1308. Rees, M., P. J. Grubb, and D. Kelly. 1996. Quantifying the impact of competition and spatial heterogeneity on the structure and dynamics of a four-species guild of winter annuals. American Naturalist 147:1–32.
Renshaw, E. 1991. Modelling biological populations in space and time. Cambridge Studies in Mathematical Bi-ology. Cambridge University Press, Cambridge. Royama, T. 1992. Analytical population dynamics.
Chap-man & Hall, New York.
Schemske, D. W., and P. Bierzychudek. 2001. Evolution of flower color in the desert annual Linanthus parryae: Wright revisited. Evolution 55:1269–1282.
Shigesada, N., K. Kawasaki, and E. Teramoto. 1979. Spatial segregation of interacting species. Journal of Theoretical Biology 79:83–99.
Silvertown, J., S. Holtier, J. Johnson, and P. Dale. 1992. Cellular automaton models of interspecific competition for space: the effect of pattern on process. Journal of Ecology 80:527–534.
Sole´, R. V., and J. Bascompte. 1997. Emergent phenomena in spatially extended model ecosystems. Pages 1–26in
J. Bascompte and R. V. Sole´, eds. Modeling spatiotem-poral dynamics in ecology. Springer, New York. Stoll, P., and D. Prati. 2001. Intraspecific aggregation alters
competitive interactions in experimental plant com-munities. Ecology 82:319–327.
Tilman, D. 1982. Resource competition and community structure. Princeton University Press, Princeton, N.J. ———. 1988. Plant strategies and the dynamics and
struc-ture of plant communities. Princeton University Press, Princeton, N.J.
———. 1993. Species richness of experimental produc-tivity gradients: how important is colonization limita-tion? Ecology 74:2179–2191.
———. 1994. Competition and biodiversity in spatially structured habitats. Ecology 75:2–16.
van den Bosch, F., H. D. Frinking, J. A. J. Metz, and J. C. Zadoks. 1988. Focus expansion in plant disease. III. Two experimental examples. Phytopathology 78:919–925. Verheyen, K., and M. Hermy. 2001. An integrated analysis
of the spatio-temporal colonization patterns of forest plant species. Journal of Vegetation Science 12:567–578. Warner, R. R., S. E. Swearer, and J. E. Caselle. 2000. Larval accumulation and retention: implications for the design of marine reserves and essential fish habitat. Bulletin of Marine Science 66:821–830.
Webb, C. O., and D. R. Peart. 2001. High seed dispersal rates in faunally intact tropical rain forest: theoretical and conservation implications. Ecology Letters 4: 491–499.
Wilson, W. G., C. W. Osenberg, R. J. Schmitt, and R. Nisbet. 1999. Complementary foraging behaviors allow coexistence of two consumers. Ecology 80:2358–2372. Yodzis, P. 1989. Introduction to theoretical ecology. Harper
& Row, New York.