Composition, abundance and biomass of the benthic fish fauna from
the Guaritico river of a Venezuelan floodplain
C . L a s s o1 J . C a s t r o v i e j o2 K e y w o r d s : Fish c o m p o s i t i o n , a b u n d a n c e , b i o m a s s , b e n t h i c fish, f l o o d p l a i n s y s t e m s , G u a r i t i c o R i v e r , V e n e z u e l a . T h e c o m p o s i t o n , a b u n d a n c e a n d b i o m a s s o f t h e b e n t h i c fish f a u n a w e r e s t u d i e d a l o n g a 6 k m s t r e t c h of t h e G u a r i t i c o R i v e r . T h i s w a t e r c o u r s e is l o c a t e d in t h e w e s t e r n p l a i n s o f V e n e z u e l a a n d b e l o n g s t o t h e A p u r e River f l o o d p l a i n s y s t e m . M o n t h l y c o l l e c t i o n s f r o m J u n e , 1990 t o J a n u a r y , 1991 w e r e m a d e b y m e a n s o f a s m a l l t r a w l i n g a p p a r a t u s r i g g e d t o d u g o u t c a n o e s a c c o r d i n g t o t h e d e s i g n o f L o p e z - R o j a s et a l . ( 1 9 8 4 ) . 42 fish species w e r e i d e n t i f i e d with a d o m i n a n c e of s p e c i e s f r o m t h e o r d e r s S i l u r i f o r m e s a n d G y m n o t i f o r m e s . T h e fish a s s e m b l a g e d i f f e r e d m a r k e d l y from o t h e r V e n e z u e l a n r i v e r s y s t e m s . S e a s o n a l l y , h i g h e r d i v e r s i t y a n d e v e n n e s s w e r e o b s e r v e d at t h e h i g h w a t e r p h a s e while t h e a b u n -d a n c e , r e l a t i v e b i o m a s s a n -d C P U E w e r e h i g h e r a t l o w w a t e r .
Composition, abondance et biomasse des poissons bentniques dans la rivière de plaine Guaritico du Venezuela M o t s clés : A b o n d a n c e , b i o m a s s e , p o i s s o n s b e n t h i q u e s , s y s t è m e d e p l a i n e d ' i n o n d a t i o n , rivière G u a r i t i c o , V e n e z u e l a .
L a c o m p o s i t i o n , l ' a b o n d a n c e et la b i o m a s s e d e la f a u n e p i s c i a i r e b e n t h i q u e o n t é t é é t u d i é e s s u r u n t r o n ç o n d e 6 k m d e la rivière G u a r i t i c o . C e s y s t è m e s e situe d a n s les p l a i n e s o c c i d e n t a l e s d u V e n e z u e l a et a p p a r t i e n t a u réseau h y d r o g r a -p h i q u e d e l a rivière A -p u r e . D e s r é c o l t e s m e n s u e l l e s d e j u i n 1990 à j a n v i e r 1991 o n t é t é f a i t e s à -p a r t i r d e -p i r o g u e s é q u i -p é e s d ' u n c h a l u t ( m o d è l e L o p e z - R o j a s et a l . 1984).
S u r les 4 2 e s p è c e s d e p o i s s o n s i d e n t i f i é e s p r é d o m i n e n t les S i l u r i f o r m e s e t G y m n o t i f o r m e s . L ' a s s o c i a t i o n p i s c i a i r e dif-fère n e t t e m e n t d e celle d e s a u t r e s s y s t è m e s d e rivière d u V e n e z u e l a . Les plus g r a n d e s d i v e r s i t é et é q u i t a b i l i t é ont é t é o b s e r v é e s p e n d a n t la p é r i o d e d e h a u t e s e a u x a l o r s q u e l ' a b o n d a n c e , la b i o m a s s e r e l a t i v e e t la c a p t u r e p a r u n i t é d ' e f f o r t ( C P U E ) é t a i e n t les p l u s f o r t e s e n p é r i o d e d ' é t i a g e .
1 . I n t r o d u c t i o n
T h e study of t h e fish c o m m u n i t i e s associated with the m a i n channel of Venezuelan rivers began in 1978 with the e x p e d i t i o n of t h e r e s e a r c h vessel « E a s t -w a r d » t o t h e L o -w e r O r i n o c o . T h e results revealed the presence of a fish f a u n a practically u n k n o w n u p t o that t i m e . D u r i n g t h e expedition, a trawling a p p a -ratus was designed t o sample in shallower areas. T h e results of t h e efficiency of t h i s gear were published by L ô p e z - R o j a s et a l . (1984). Similar collections
1. Asociacion Amigos de Donana, 2182, 41080 Sevilla, Espana & Museo Historia Natural La Salle, 1930, Caracas 101O-A, Venezuela.
2. Asociacion Amigos de Donana, 2182, 41080 Sevilla, Espana.
were c a r r i e d o u t in t h e N a p o River ( E c u a d o r ) b e t -ween 1981-1983 (Stewart et al. 1987). C o l l e c t i o n s were later m a d e in t h e O r i n o c o River D e l t a using e x p e r i m e n t a l t r a w l i n g nets (Cervigon 1982, 1985, Cervigon & N o v o a 1988, N o v o a 1 9 8 2 , 1 9 8 6 , N o v o a & C e r v i g o n 1982, P o n t e 1990, R a m o s et a l . 1982). S y s t e m a t i c research o n t h e taxonomic, ecological a n d fishery aspects of t h e b e n t h i c fish f a u n a of t h e A p u r e River began in 1983 (Castillo 1988, M a c h a d o -Allison 1987, M a r r e r o 1984, 1990, P r o v e n z a n o & Castillo 1984, P r o v e n z a n o et al. 1984 a, b , c). T h i s p a p e r r e p o r t s t h e results of t h e research c a r r i e d o u t in t h e G u a r i t i c o River ( A p u r e River basin). In 1989, the river w a s d e c l a r e d a part of a specially p r o t e c -ted wildlife a r e a . T h e objectives of our r e s e a r c h were : 1) t o determine the species composition of t h e
Article available at
http://www.limnology-journal.org
72
C. LASSO, J. CASTROVIEJO(2)
fish f a u n a associated with t h e b o t t o m o f t h e river
i n o r d e r t o m a k e c o m p a r i s o n s with o t h e r similar
systems ; 2) t o quantify the a b u n d a n c e , b i o m a s s ,
a n d catch per unit effort ( C P U E ) determined for the
first t i m e by means of the a p p a r a t u s designed by
L ô p e z - R o j a s et al. (1984).
2 . S t u d y a r e a
The study was m a d e on the last 6 km stretch of
the Guaritico River (locally known as c a n o G u a r i
-tico) before its confluence with the Apure River (Fig,
1). The Guaritico River is located on the upper
sec-t i o n of sec-the A p u r e River Basin (Ramia 1972) in a
Fig. 1. Study area (dots indicate the studied river section).
region of seasonally flooded savannas (Scharger &
Gonzalez 1973, W e l c o m m e 1979). T h e climatology
of the area shows t w o contrasting periods ; the dry
season, which extends from N o v e m b e r to April a n d
the rainy season, which extends from May to
Octo-ber. These climatic conditions lead to two
hydro-metric periods : low waters (December-May) a n d
high waters (June-November) (Fig. 2). In this study,
collections of fish were m a d e during b o t h periods.
<
J F M A M J J A S O N D
Fig. 2 . Water level variations of the Guaritico River (1990). Fig. 2 . Variations du niveau de l'eau de la rivière Guaritico ( 1990).
During 1990, m e a n monthly air t e m p e r a t u r e ranged
between 25.1 ° C (July) a n d 28.9 ° C (April). T h e
Guaritico River supports a seasonally inundated
gal-lery forest. T h e m o s t representative tree species are
Nectandria pichurini ( H . B . K . ) Mez. a n d Dugnetia
riberensis Arist. O n the river side next to the
gal-lery forest, there is also a t r e e c o m m u n i t y k n o w n
locally as « mangle », d o m i n a t e d by Coccoloba
obtusifolia Jacq. (Castroviejo & Lopez 1985).
According to Sioli's classification o f A m a z o n i a n
river waters, the Guaritico River shows clear-waters
characterized by a smaller concentration of
suspen-ded solids and a higher water transparency t h a n
white-waters. Ranges for p H , water t e m p e r a t u r e ,
and conductivity were : 6.0-7.8 p H units ; 28-32 ° C ;
a n d 19.4-21.9 ^ S . c m
- 1, respectively. W a t e r
trans-parency (Secchi) showed seasonal oscillations during
the year a n d was generally higher near t h e c o n
-fluence of the Guaritico with the Apure River. Water
t r a n s p a r e n c y was 25 cm at the end of t h e dry
sea-son a n d 20 cm at the beginning of the rainy s e a s o n .
T h e b o t t o m of the channel showed the t w o types o f
clays (grey and red) described by Marrero (1990) for
t h e A p u r e River.
3 . M a t e r i a l s a n d M e t h o d s
Monthly collections were made during the high a n d
low water periods between June 1990 a n d J a n u a r y
1991. Collections were made with a trawling fishing
apparatus adapted to dugout canoes according to the
design made by Lôpez-Rojas et al. (1984). Each
traw-ling lasted for 10 min, covering a mean distance of 147
m . , at depths ranging from 2-10 m (Table 1). Catches
were preserved with formaldehyde in the field. At the
laboratory, fishes were identified, weighed and
mea-sured. Species abundance was estimated with respect
to the total fishes caught monthly. Diversity ( H ' ) was
calculated by the Shannon-Weaver index (1949) a n d
the evenness (V*) was determined as the inverse value
of H* max. Species biomass was calculated as the
per-centage in weight with respect to the total fish caught
a n d also in terms of k g . h a
- 1. The latter value a n d
density (ind. ha~ ') refer to the area covered by a 10
min trawling effort. The C P U E is expressed as
k g . h a . h o u r ~
4. R e s u l t s
4.1. Fish species
42 species from 5 orders, 13 families and 38 genera
were identified (Table 2). W i t h 26 spp (61 °7o) t h e
Siluriformes were the best represented g r o u p ,
fol-lowed by the Gymnotiformes (11 spp., 26.2 % ) , a n d
by the Rajiformes, Perciformes, a n d P l e u r o n e c t i
-formes which together represented 12 % (5 s p p . ) o f
the identified species (Table 3). The dominant
families were t h e Loricariidae, Pimelodidae, a n d A p t e
-ronotidae with 10,9 a n d 7 species, respectively. F o u r
t a x a were identified only to t h e generic level
(Pota-moirygon s p . , Hemiancislrus s p . , Duopalatinus s p . ,
a n d Rhamphichthys s p . ) . A new genus a n d species
from the family Loricariidae was recorded ( P r o v e n
-z a n o F . , pers. c o m m . ) as well as a new species from
t h e genus Porotergus (Apteronotidae).
Aphanotu-rulus frankei (Loricariidae) represents a new record
for the Venezuelan fish fauna. Before o u r r e p o r t ,
this species was k n o w n only from the Ucayali River
Basin in P e r u .
74 C . LASSO, J. CASTROVIEJO
(4)
Table 1. Total number of trawls, effective trawls a n d time of trawling .Period : June. 1990 - January, 1991. Tableau 1. Nombre total de chaluts, chaluts efficaces, et durée de chalutage. Période : juin 1990 - janvier 1991. M O N T H S J U N E JULY AUGUST SEPTEMBER OCTOBER NOVEMBER DECEMBER JANUARY N u m b e r of trawls 13 9 18 12 12 18 11 9 N u m b e r of efectivetrawls 12 9 18 II 11 16 11 8 t total trawls
(minutes) 120 90 180 110 110 160 110 80
Table 2. List of fish species associated with the bottom of the Guaritico River. Tableau 2. Liste d'espèces de poissons benthiques de la rivière GuanrJco. R A J I F O R M E S
P O T A M O T R Y G O N I D A E
01 Potamotrygon orbignyi (Castelnau) 1885 02 Potamotrygon sp.
S I L U R I F O R M E S A G E N E I O S I D A E
03 Ageneiosus brevififis Cuvier & Valenciennes 1840 04 Ageneiosus ucayalensis Castelnau 1855 C E T O P S I D A E
05 Cetopsis coecutiens (Lichtenstein) 1819 D O R A D I D A E
06 Leptodoras Imne/li Eigenmann 1940 07 Megalodoras irwini Eigemann 1925 08 Pterodoras apurensis (Fernandez- Yépe/r 1965 H Y P O P H T H A L M I D A E
09 Hypophlhalmus edematus Spi\ 1829 L O R I C A R I I D A E
10 Aphanoiorulus Jrankei Isbrucker y Nijssen 1985 11 Hemiancistrus sp.
12 Limatutivhthys punctatus (Regan) 1904 13 Loricaria cataphmcta Linnaeus 1758 14 Loricarichthys niaculatus (Bloch) 1794 15 Panaque nigroimeutus (Peters) 1877 16 Pseudohemioduri luticepts (.Regan) 1904 17 Pterygoplichthys ntultirrudiatus (Hancock) 1824 18 Sturisoma rosirarumes (Spi.x) 1829 19 Loricariidae ncn. et sp. nov. P I M E L O D I D A E
20 Callophysus mucrupterus (Lichtenstein) 1819 21 Duopatatmus *p.
22 Hemisorubirn platyrhynchos (Cuvier & Valenciennes) 189(1 23 Pimelodidae gen. et sp. nov.
24 Pimelodelta gracilis (Cuvier & Valenciennes) 1890 25 Pimetodus blochii Valenciennes 1840 26 Pimetodus altissimus Eigenmann & Pearson 1942 27 Pinirampus pinirampu (Spix) 1829
28 Pseudoplatystoma fasciatum (Linnaeus) 1766 GYMNOT1FORMES
APTERONOTIDAE
29 Adontosternarchus devenanzii Mago-Leccia, Litndber y Baskin. 1985
30 Adontosternarchus sachsi (Peters) 1877 31 Apieronotus bonapartii (Castelnau) 1855 32 Porotergus sp. nov.
33 Sternarchoxiton porcinum Eigenmann & Allen 1842 *4 Siernarchorhamphus muelteri (Steindachner) IH81 35 Sternurchorhynchus curvirosrris (Boulenger) 1877 R H A M P H 1 C H T H Y I D A E
36 Rhamphichthys s p . STERNOPYGIDAE
37 Distocyctus conirostris Eigenman & Allen 1942 39 Rhabdolichops eatswardi Lùndberg & Mago-Leccia, 1985 PERCIFORMES
C I C H L I D A E
40 Geophagus altifrons Heckel 1840 SCIAENIDAE
41 Plagtoscion squamostssimus (Heckel) 1840 SOLFIDAE
42 Hypociinemus mentalis (Gùnther) 1862
Table 3. Families, genera, and species for each of the orders trom the Guantico River. Tableau Nombres de familles, de genres et d'espèces de chaque ordre de la rivière Guaritico.
O R D E R FAMILIES GENERA SPECIES/ORDER %
R A J I F O R M E S 1 1 2 4,8 S I L U R I F O R M E S 6 24 26 61,9 G Y M N O T I F O R M F S 3 10 11 26,2 P E R C I F O R M E S 2 2 2 4,8 P L E U R O N E C T I F O R M E S 1 1 1 2,4 T O T A L 13 38 42 100
4 . 2 .
Diversity a n d a b u n d a n c e
Highest diversity values were recorded between
June and September with a m a x i m u m in August
(Fig. 3). Diversity decreased between October a n d
J a n u a r y with lowest values in December. Species
richness values were highest in J u n e (21 ssp.) a n d
lowest in January (13 ssp.). Evenness showed a simi
lar trend to diversity. Highest equity values were
recorded in August while lowest values were recor
ded in December.
Considering the collections for the enure sampling
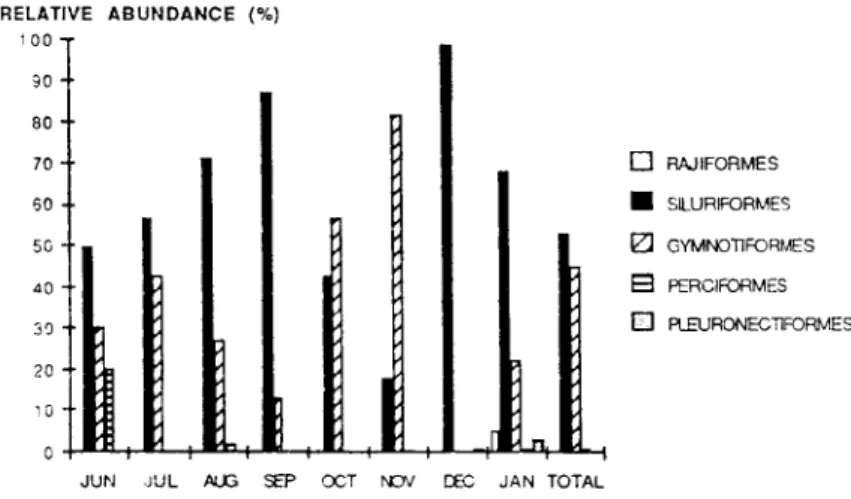
period, the Siluriformes was the most a b u n d a n t
order (53.3 9b) followed by the G y m n o t i f o r m e s
(44.7 % ) . The remaining orders represented only
2 9o. Except in O c t o b e r a n d November, the Siluri
formes was the dominant group throughout the sam
pling period. In December, this g r o u p represented
almost 100 9b of the catches (Fig. 4). Except for the
Gymnotiformes, the c o n t r i b u t i o n of the other
groups was small. In J u n e , however, a b u n d a n c e of
the Perciformes reached 20 9b. Figures 5 a-c show
the monthly relative a b u n d a n c e of each species.
MINT!
(vi
jun Jul Aug Sap Oct Nov Dac Jan
Fig. 3. Variations of fish diversity ( H ' ) and evenness. Fig. 3. Vaiiations de la diversité (H') et de l'équitabilité pisciaires.
RELATIVE ABUNDANCE (%)
1 0 0 t 9 0 8 0 7 0 5 050
40
30
20
1 0
0
i
U T
• RAJIFORMES • S I L U R I F O R M E S 0 GYMNOTIFORMES B P E R C I F O R M E S EU PLEURONECT1FORMESJ U N J U L AUG SEP OCT NOV DEC J A N T O T A L
Fig. 4. Monthly relative abundance of each fish order. Fig. 4. Abondance mensuelle relative des 5 ordres de poissons.
76 C . LASSO, J. CASTROVIEJO (6) JUr€, 1 9 9 8 SP8 s p l 2 Spl4 s p l 8 5P22 sp27 sp32 sp34 sp37 sp39 SP41 SP9 SP13 SP16 SP19 SP21 SP29 SP33 SP35 SP38 SP4B flUQUST, 1998 Sp4 s p l 0 s p l 3 s p l 8 Sp23 sp25 Sp27 S p 3 1 sp34 sp39 sp7 SP12 3pl6 3p21 sp24 sp26 sp29 3p32 sp38 sp41
I A B U N D A N C E {%)
IWEIGHT(%)
Fig. 5 a. Relative abundance and biomass of fish species for June-August, 1990 (codes on Table 2). Fig. 5 a. Abondance relative et biomasse des différentes espèces de poissons en juin-août 1990 (code-chiffres : tableau 2).
J u n e was the only m o n t h in which the m o s t a b u n -d a n t species (Plagioscion squamossisimus, 17.8 % ) did n o t belong to t h e Siluriformes or G y m n o t i f o r -mes. This species was followed by Hypophthalmus
edentatus (12.2 °/o). In July, Pimelodus blochii s h o
-wed a 20.4 °7q a b u n d a n c e follo-wed by
Sternarchorhamphus muelleri with 13.6 % . In August, a b u n
-dance was similar a m o n g species except for
Loricaria cataphracta a n d Duopalatinus sp. which d o m i
-nated with 17.7 % and 15.2 % , respectively. P.
blo-chii was again the most a b u n d a n t species (35.7 % )
SEPTEMBER, 1998
s p 5 S P 6 Sp8 5P13 SP14 SP16 SP18 s p l 9 SP21 SP22 sp25 Sp26 SP28 sp38OCTOBER. 1990
SP13 SP14 SP18 S P 1 9 SP21 S P 2 5 Sp26 S P 2 9 S P 3 8 S P 3 2 S P 3 8 s p 3 9 sp41NOVEMBER. 1998
Sp6 S P 1 3 Spl4 S P 1 8 Sp21 S P 2 5 s p 2 6 5 P 2 7 S P 2 9 Sp38 Sp34 s p 3 7 s p 3 8 S P 3 9 Sp41• A B U N D A N C E ^ ) ^ W E I G H T ( % )
Fig. 5 b . Relative abundance a n d biomass of fish species for September-November, 1990 (codes on Table 2 ) . Fig. 5 b . Abondance relative et biomasse des différentes espèces de poissons en septembre-octobre 1990 (code-chiffres : tableau 2).
(18.4 °7o). In O c t o b e r , a G y m n o t i f o r m
(Eigenman-nia macrops) showed highest abundance values (24.9
% ) for the first t i m e . This species was followed b y
L. cataphracta (23.4 % ) . The Gymnotiformes clearly
dominated in November {Rhabdolichops eastwards
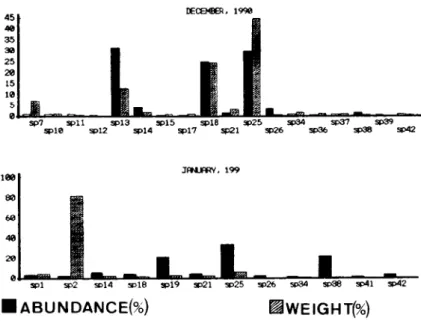
28.6 % ; E. macrops, 24.9 °7o). In December, L .
cataphracta a n d P. blochii showed again highest
a b u n d a n c e (60.8 % for b o t h species). Finally, in
J a n u a r y , P. blochii was again dominant (32.7 % ) .
7 8 C. LASSO, J . CASTROVIEJO (8)
1 0 0
s p l s > 2 s o l 4 SP18 SP19 S P 2 1 S P 2 5 S P 2 6 S P 3 8 seAl 5 P 4 2
• ABUNDANCE(%) ^WEIGHT(%)
Fig. 5 c. Relative abundance a n d biomass of fish species for December (1990)-January (1991) (codes on Table 2). Fig. 5 c. A b o n d a n c e relative et biomasse des espèces de poissons en décembre 1990-janvier 1991 (code-chiffres : tableau 2).
In t e r m s of i n d . h a ~ lowest values were recor-d e recor-d b e t w e e n J u n e a n recor-d S e p t e m b e r with m i n i m u m v a l u e s in A u g u s t (115 i n d . h a-1) - H i g h e s t
(0
1 0 0 0 . C"b
s e eç
—
c
2 0 04)
a
0values were recorded between October a n d J a n u a r y ( m a x i m u m of 870 ind. h a-1 in December (Fig. 6).
Fig. 6. Monthly fish abundance variations (ind. ha ' ) . Fig. 6 . V a r i a t i o n s m e n s u e l l e s d ' a b o n d a n c e pisciaire ( i n d . h a- 1)
-B E N T H I C FISH FAUNA F R O M T H E GUAR1TICO R I V E R
4
. 3 .
Biomass and C P U I
W i t h respect lo the total catch, the relative bio
mass indicated a d o m i n a n c e of the Siluriformes
(64.9 % ) followed by t h e Rajiformes (21.5 °7o), and
Gymnotiformes (12.9 °?o). T h e Perciformes and the
Pleuronectiformes together indicated biomass values
< 1 % . During the 8 m o n t h s sampling p e r i o d , bio
mass of the Siluriformes was the highest during 6
m o n t h s (65 % ) . Only in N o v e m b e r a n d J a n u a r y ,
was this g r o u p displaced by the G y m n o t i f o r m e s
(52.9 % ) and by the Rajiformes (85.1 <7o), respecti
vely (Fig. 7). T h e relative b i o m a s s values for each
species are shown in Figures 5 a-c. In J u n e t h e rela
tive b i o m a s s v a l u e for Pimetodus altissimus was
24.2 % while a new genus a n d species (Loricariidae)
had a value of 17.4 °/o. In J u l y - A u g u s t ,
Megalodo-ras irwini had t h e highest b i o m a s s values (36.6 %
a n d 82.8 % . respectively). Only in S e p t e m b e r , did
we observe highest a b u n d a n c e values corresponding
to highest b i o m a s s values for one species (P. blo
chii, relative b i o m a s s - 42.9 °/o). In O c t o b e r , L.
cataphracta was first (32.3 °7o) followed by E.
macrops (32.5 °?o). In N o v e m b e r , a G y m n o t i f o r m
(Sternarchorhamphus muelleri) showed the highest
biomass values (3.0 % ) . P. blochii (44.7 % ) a n d
Stu-risoma rostrata (24.3 °7o) represented t h e most
important species in December. Highest b i o m a s s of
the Rajiformes (81.3 % ) was observed in J a n u a r y
for t h e first time.
In terms of k g . h a
- 1, t h e lowest biomass value
was recorded in J u n e (1.7 k g . h a ' ) • '
nJuly-August, biomass increased slightly to 9.6 k g . h a ~
1and then decreased in September-November. Hig
hest values were observed in D e c e m b e r - J a n u a r y
(26.3 k g . h a - 1 in January) (Fig. 8). The C P U E sho
wed a similar pattern t o biomass. Lowest values cor
responded to J u n e (10.6 k g . h a
- 1) a n d highest
values to J a n u a r y (164.4 k g . h a
-•). (Fig. 9).
5 . D i s c u s s i o n
Most of the species from the Guaritico River
belong to t h e O r d e r s Siluriformes and G y m n o t i f o r
mes (88.1 % ) . This situation contrasts with records
from other neotropical freshwater ecosystems cha
racterized by the d o m i n a n c e of characoid fishes
RELATIVE BIOMASS (%)
1 0 0
T • R A J I F O R M E S • SILURIFORNES E2 GYMNOTIFORMESB
PERCIFORMES
D PLEURONECTlFORtvtSJ U N J U L A U G S E P OCT NOV CEC J A N TOTAL
Fig. 7. Monthly variations of the relative biomass of Fish order. Fig. 7. Variations mensuelles de la biomasse relative des 5 ordres de poissons.
80
C . L A S S O , J. CASTROVIEJO(10)
ta
2 8 2 4 O>20 JE—
1 6 (0 1 2 CD 1 2m
8E
8o
4m
aN
Fig.Fig. 8. Monthly fish biomass variations ( k g . h a- 1) . . Variations mensuelles de la biomasse de poisson (Kg. h a- 1) .
ta
1 8 8 • C 1 6 0 O) 1 4 0 _ 1 2 0 UJ 1 9 8 1 3 8 0a.
6 0O
40 2 0 0 ,.S
N
Fig. 9. C P U E variations ( k g . h a . h1) .Fig. 9. Variations de la capture par unité d'effort : C P U E ( K g . h a . h- 1) .
( L o w e - M c C o n n e l 1975, 1987 ; Lowe-McConnell &
H o w e s 1981). Similar results have been reported for
t h e b e n t h i c fish c o m m u n i t i e s of the A p u r e River
( P r o v e n z a n o & Castillo 1984 ; P r o v e n z a n o et al.
1984 a, b , c) a n d of t h e Orinoco River Delta
(Lôpez-R o j a s et al. 1984 ; (Lôpez-R a m o s et al. 1982). In the lower
O r i n o c o River, L ô p e z - R o j a s et al. (1984) distinguis
h e d t w o fish assemblages ; o n e assemblage in t h e
deepest sections of t h e m a i n channel d o m i n a t e d by
G y m n o t i f o r m e s , a n d a n o t h e r o n e in adjacent shal
lower a r e a s d o m i n a t e d by high richness of
chara-coid fishes. In the G u a r i t i c o River, however,
cha-racoids specially the « p i r a n h a s » (Pygocentrus a n d
Serrasalmus) represent accessory species. Their pre
sence in o u r catches m a y be considered accidental
as they were caught in the net while preying on other
fishes.
In spite of the absence of complete fish lists, we
c o m p a r e d the preliminary fish inventories of ben
thic species of the Guaritico a n d Apure rivers a n d
o f the O r i n o c o Delta. This comparison revealed a
higher similarity between the Apure and the Guari
tico River. T h e basic difference between these two
rivers was the absence of certain genera in the Gua
ritico River : Aspredinidae (Xyliphius), Cetopsidae
(Pseudocetopsis), Pimelodidae {Megalonema,
Pime-lodina, Platysilurus, Pseudopimelodus), Loricarii
dae (Apistofohcaha, Denteclus, Farioweila,
Lamontichthys, Paraloricaria, Spatutoricaria). A p t e r o n o
-tidae (Sternarchella, plus two new genera) (Proven
z a n o F . , pers. c o m m . ) . T h e presence of o t h e r spe
cies from the Rajiformes, Clupeiformes,
Percifor-mes, a n d Pleuronectiformes recorded by us in
t h e Guaritico River and not yet recorded from the
B E N T H I C FISH FAUNA F R O M T H E G U A R I T I C O RIVER
81
Table 4. Accompanying fish species found during the trawlingof the bottom of the Guaritico River.
Tableau 4. Espèces de poissons accompagnants trouvées pendant le chai ut age du fond d e la rivière Guaritico. C L U P E I F O R M E S
C L U P E I D A E
Pellona Jlavipinnis (Valenciennes) 1839 C H A R A C I F O R M E S
A N O S T O M I D A E
Leporinus fasciatus (Bloch) 1794 C H A R A C I D A E
Acestrocephalus sp. Galeocharax gulo (Cope) 1870 Knodus breviceps (Eigenmann) 1908 Mylossoma duriventris (Cuvier) 1818 Pygocentrus caribe (Valenciennes) 1849 Roeboides a/finis (Gunther) 1868 Serrasalmus alîuvei Ramirez 1965 Serrasalmus irrilans Peters 1877 Serrasalmus medinai Ramirez 1965 C U R I M A T I D A E
Curimata sp.
Steindachnerina argenlea (Gill) 1858 Steindachnerina s p .
H E M IO D O N T I D A E
Hemiodus unimaculatus (Bloch) 1794
A p u r e River, is m o s t p r o b a b l y due t o t h e absence
of m o r e conclusive fish inventories t h a n t o the
absence of these orders in the latter. W i t h regard
to the benthic fish fauna of the Orinoco River
(exclu-ding typically e s t u a r i n e species), some g e n e r a are
found in this region which have not yet been
recor-ded from the Guaritico River : Engraulidae
(Ancho-vict, Lycengraulis), A s p r e d i n i d a e (Hoplomyzon,
Platystacus), D o r a d i d a e (Opsodoras), Pimelodidae
(Nannorhanmdia, Perugia), H y p o p o m i d a e
(Steato-genys), Soleidae ( A p i o n i c h t h y s ) .
T h e difference in the n u m b e r of fish species from
different rivers has been related t o the size of the
d r a i n a g e surface a n d length of the main river c h a n
-nel (Welcomme 1985). This observation m a y explain
the lower a b s o l u t e n u m b e r of fish species (alpha
diversity according to Lowe-McConnell 1987)
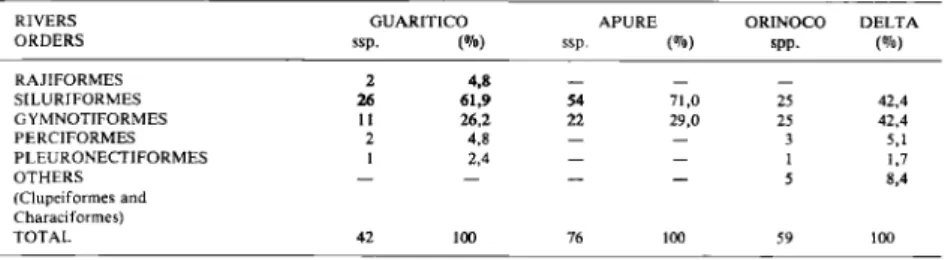
recor-ded from the G u a r i t i c o River in comparison to the
A p u r e River and t h e O r i n o c o Delta (Table 5). T h e
lower number of fish species from t h e Guaritico m a y
also be the result of t h e smaller transects sampled
by us (some 6 km) c o m p a r e d to t h e transects
sampled in the A p u r e River a n d Orinoco Delta. A n o
-ther factor t o consider in explaining the diversity
a n d distribution o f fish species of the G u a r i t i c o
River is the type of w a t e r . T h e clear w a t e r s of
this river differ from t h e white waters of t h e A p u r e
a n d O r i n o c o rivers. The importance of water
types on the distribution a n d diversity of n e o t r o
-pical fishes has previously been discussed (Weitzman
& W e i t z m a n 1982). In the A p u r e River, the type
of s u b s t r a t u m has also been linked to the
distribu-t i o n of some Siluriform and Gymnodistribu-tiform fish
spe-cies (Provenzano & Castillo 1984, P r o v e n z a n o et al.
1984 a, b , c). In o u r study, the available d a t a d o
not allow us to consider yet any possible
relations-hips between the type of s u b s t r a t a and fish species.
In the Guaritico River, diversity a n d evenness
sho-wed a similar temporal p a t t e r n . Highest values were
observed at high waters rather t h a n low w a t e r s . A
g r o u p of fish species was observed d u r i n g b o t h
periods (Table 6) a n d a t least 10 species of catfishes
(Siluriformes) were collected exclusively at high
waters. These species may be migrating f r o m the
A p u r e River into the Guaritico River.
Table 5. Comparison a m o n g the number of fish species from each order from the Guaritico River, Apure and Orinoco River Delta. Tableau 5. Comparaison des nombres d'espèces de chaque o r d r e dans les rivières Guaritico et Apure et dans le delta de l ' O r é n o q u e .
RIVERS G U A R I T I C O A P U R E O R I N O C O D E L T A O R D E R S ssp. ssp. (%) spp.
m
R A J I F O R M E S 2 4.8_
_
S I L U R I F O R M E S 26 61,9 54 71,0 25 42,4 G Y M N O T I F O R M E S 11 26,2 22 29,0 25 42,4 P E R C I F O R M E S 2 4,8 — — 3 5,1 P L E U R O N E C T I F O R M E S 1 2,4 — — 1 1.7 O T H E R S — — — 5 8,4 (Clupeiformes a n d Characiformes) T O T A L 42 100 76 - 59 10082 C. LASSO, J. CASTROVIEJO
(12)
Table 6. Presence and size range (mm) of the species collected in the Guaritico River. Measurement1; for the Gymnotiformes = total length ; for the Rajiformes - discal width ; other orders = standard length.
T a b l e a u 6. Présence et taille moyenne (mm) des espèces récoltées dans la rivière Guaritico. Mensurations pour les Gymnotiformes -longueur totale ; p o u r les Rajiformes = largeur du disque ; autres ordres - -longueur stardard.
SIZE RANGES (mm)
SPECIES JUNE JULY AUGUST SEPTEMBER OCTOBER NOVEMBER DECEMBER JANUARY
Potamotrygon orbignyi 122-125 Potamotrygon sp. 330-430 Ageneiosus brevifilis 37-63 A. ucayatensis 43-59 108 Cetopsis coecutiens 36-39 37 Leplodoras tinnelli 120-146 145 Megaloàoras irwmi 430 400-540 185-271 Pterodoras apurensL 65 66 53 Hypophlhalmus edentatus 36-38 71-100 Aphanotorulus frankei 115 59-155 Hemiancislrus sp. 79-81 Ltmatulichthys punclatus 28-84 114-128 114-145 128 Loricaha cataphracta 62-69 48-81 72-188 88-224 56-124 75-191 80-210 Loricarichthys maculatus 56 118-190 98-114 102 63-162 164-181 Panaque nigrotineatus 260 Pseudohemiodon laliceps 33-34 48-75 60-61 45-92 P. mttltirradialus 119 Sturisoma rostralum 223 88-86 159 75-113 184-227 108-235 176-214
New genus and species 162-166 71-177 123-127 101-182
Catiophysus macropterus 83
Duopalatinus sp. 29 51-150 56-145 165 141-190 142-196 124-159
Hemisorubim platyrhynchos 130 132
New genus and species 164 42-129
Pimelodella gracilis 54-62 52-64 43-74 72-40 52-73 55-57 Pimetodus altissimus 112-127 120-135 53-156 Pimetodus blochii 72-45 77-133 53-156 56-75 62-122 62-135 66-137 Pintrampus pinirampu 62-67 49-248 61-200 Pseudoplalystoma fasciatum 250 A. devenanzi 138 64-121 83-84 115-124 92-111 A. sachsi 93-147 93-139 Apteronotus bonapartit 54-318 279 Porotergus sp. n. 179-218 236-260 69-164 56-93 Slernarchogiton poreinum 88 S. muelleri 104-390 150-253 146-349 S. curvirostn 270 Rhamphichthys sp. 324 Distoctclus coniroslris 223 261 283 285-299 Eigenmannia macrops 205-229 155-244 75-184 96-264 48-259 90-264 75-216 86-120 Rhabdolichops catswardi 180-280 174-243 56-217 57-205 105 Geophagus altifrons 25 Plagioscion squamosstsimus 35-39 62-66 125 105-194 16 Hypoclinemus mentalis 62-80 45-50
BENT HIC FISH F A U N A FROM T H E G U A R I T I C O RIVER
83
T h e overall fish a b u n d a n c e was higher at low
water a n d lower at high watei A similar trend has
been reported for G y m n o t i f o r m a n d L o n c a r i i d a e
fishes from the A p u r e River ( P r o v e n z a n o F . , u n p u
-blished). During the dry season, water level a n d
cur-rent a r e r e d u c e d . At t h e onset ol the rams a n d the
s u b s e q u e n t increase in water level, water depth a n d
water current a r e increased. These conditions m a y
explain the smallei fish a b u n d a n c e . In our case,
however, t h e smallest fish density was recorded at
low water ( J a n u a r y ) . H o w e v e r , b i o m a s s values
during this m o n t h w e t e highest due to t h e presence
of Rajiform fishe.s (low a b u n d a n c e a n d high
b i o m a s s ) .
Similar to r e p o r t s for the O r i n o c o River Delta
( L ô p e z - R o j a s et al. 1984), we did not find a clear
relationship between t h e C P U E and other variables
(time of collections, lengths of trawling distances,
d e p t h of trawlings, type of s u b s t r a t a ) . T h e r e w a s
however, a relationship between the C P U F a n d t h e
period of the year. Highest C P U F valuer were
recor-d e recor-d at low water. Collections ol fishes in the
Gtia-ritico River with the same fishm,. &eu! ranged b>n
ween 1-130 individuals p e r trawi. In r!u. O r i n o c o
Delta, the n u m b e r of fishes caught per trawl was
always higher t h a n 100. This i n f o r m a t i o n is
indica-tive of t h e high a b u n d a n c e and fish b i o m a s s found
in the O r i n o c o Delta with respect t o the G u a r i t i c o
River. T h e selectivity of the fishing a p p a r a t u s ,
favouring the catch of smaller fishes, was observed
in our work where we caught specimens r a n g i n g
from 33 m m to 540 m m s t a n d a r d length
(Megalo-doras irwini) and 430 m m discal width
(Potamotry-gon sp.) (Table 6). T h e b o t t o m of the G u a r i t i c o
River showed an assemblage of fish species in
dif-ferent developmental stages ; species which inhabit
the b o t t o m for a period of their life-cycles (i.e.
ben-thonic y o u n g stages of Siluriforms) or which
inhabit the b o t t o m t h r o u g h most of their d e v e l o p m e n
-tal stages (Table 7).
C o m p a r i n g our d a t a with the information p r o v i
-ded by M a c h a d o - A l l i s o n (1987) a n d by P e n c z a k &
Lasso (1991), the highest b i o m a s s value (26.3 Kg.
ha ' ) recorded for the Guaritico River is low c o m
-pared t o mean values for other tropical freshwater
river systemps.
A c k n o w l e d g e m e n t ! !
We Vkish IO thank the A s o c i a c i ô n A m i g o s de D o nana for the partial funding of this protect and O Castillo ( F O N A I A P ) , C . A . Invega. Fun-d a c i ô n La Salle Fun-de C i e n a .is Naiurales, anFun-d members of the family Mal-dortado for their logistic support ; X. Etguezàbal, H . Pinango and B. Mora (M A . R . N . R . ) for their valuable assistance in t h e field work. W e specially thank E . Vâsquez for hK critical reading and for his English translation. W. Wilbert helped us in the editing o f the English version. In a d d i t i o n we recognize A . Rial, R. Perez and V. P o n t e for their c o n t r i b u t i o n in the production of the graphs ; H . L ô p e z - R o j a s . A . Machado- Allison and F . P r o v e n z a n o (Universidad Central d e Venezuela) who provided
valua-s and finally we thank M. Garcia for her typing o f the papier.
Table 7. Species and m o n t h of collection of juvenile stages associated with the b o t t o m of t h e Guaritico River. Tableau 7. Espèces et mois de capture des stades jeunes récoltés sur le fond de la rivière Guaritico.
MONTHS JUNE JULY AUGUST SEPTEMBER
A. brevifilis X A. ucayalensis X C. coecutiens X X P. apurensis X X X H. edentatus X X A. frankei P. laticeps X X C. macropterus X Duopalatinus sp X X P. altissimus X P blochii X P. fasciatum X P. pinirampu X G. altifrons X P. squamossisimus X X
84 C . L A S S O , J . CASTROVIEJO
(14)
ReferencesCastillo O. 1988. — Aspectos biolôgicos y pesqueros sobre los peces comerciales del Bajo Llano con enfâsis en los bagres (Orden Siluriformes). M . S c . , Universidad Central de Vene-zuela, Caracas, 112 p .
Castroviejo S. & Lôpez G. 1985. — Estudio y descripciôn de las comunidades végétales del « H a t o El Frio » en los Lla-nos de Venezueal. Mem. Soc. Cienc. Nat. La Salle, 45 (124) : 79-151.
Cervigôn F. 1982. — La ictiofauna estuarina del Cano Mànamo y areas adyacentes. In D . Novoa (Comp.). Los Recursos Pes-queros del Rio Orinoco y su Explotacion. Corporaciôn Vene-zolana d e G u a y a n a , Editorial Arte, Caracas, 205-260. Cervigôn F . 1985. — La ictiofauna de las aguas costeras
estua-rinas del Delta del Orinoco en la costa Atlàntica occidental C a r i b e . In A. Yaftez-Arancibia (Ed.). Fish Community Eco-logy in Estuaries and Coastal Lagoons : Towards and Ecosys-tem Integration, 5 : 57-78.
Cervigon F. & Novoa D . 1988. — Las Pesquerias Continenta-les en el Eje Orinoco-Apure. Perspectivas Futuras y Alter-nativas de Ordenamiento. Parte I : Sector Delta del Orinoco. ( P S P A ) Informes Técnicos para planes. D G S P O A / I T P / 6 3 , 109 p .
Lôpez-Rojas H . , Lundberg J. & Marsh E. 1984. — Design and operation of a small trawling apparatus for use with dugout canoes. North Am. Journ. of Fish. Manag. 4 : 331-334. Lowe-McConnel R . H . 1975. — Ecology of Fishes in Tropical
Freshwaters : Their Distribution, Ecology and Evolution. L o n g m a n , London : 337 p .
Lowe-McConnel R . H . 1987. — Ecological Studies in Tropical Fish Communities. Cambridge University Press, London, 382 p .
Lowe-McConnel R . H . & Howes G. 1981. — Pisces. In H. Hul-bert, G. Rodriguez & N. Santos (Eds.). Aquatic Biota of Tro-pical South America, Part. 2, Anarthropoda. San Diego State University Press : 218-229.
Machado-Allison A. 1987. — Los peces de los Llanos de Vene-zuela : Un Ensayo Sobre su Historia Natural. Universidad Central de Venezuela, C D C H : 141 p .
M a r r e r o C . 1987. — Notas preliminares acerca de la historia natural d e los peces del Bajo Llano. I. — Comparaciôn de los hâbitos alimentarios de très especies de peces Gymnoti-formes, en el Rio A p u r e (Edo. Apure, Venezuela). Rev. Hydrobiol. trop. 20 (1) : 57-63.
M a r r e r o C . 1990. — La ecologia trôfica de los Apteronotidae (Pisces, Teleostei, Gymnotiformes), en el rio Apure, Vene-zuela, enfocada a través de su morfologia buco-cefâlica externa y sus relaciones filogenéticas. Ph. D. dissertation, Uni-versidad Central de Venezuela, Caracas : 76 p . N o v o a D . 1982. — Los Recursos Pesqueros del Rio Orinoco y
su Explotacion. Corporaciôn Venezolana de Guayana, Edi-torial A r t e , Caracas, 386 p .
N o v o a D . 1986. — Resultados de los muestreos de fondo en el area estuariana del Delta del Rio Orinoco, Venezuela. I O G / F A O W o r k s h o p on Recruitment in Tropical Coastal Demersal Communities. Suplement 44.
Novoa D. & Cervigôn F. 1982. — Catàlogo de los recursos pes-queros del Delta del Orinoco. In D. Novoa (Comp.). Los Recursos Pesqueros del Rio Orinoco y su Explotacion. Cor-poraciôn Venezolana de Guayana, Editorial Arte, Caracas : 261-360.
Penczak T. & Lasso C. 1991. — Problems of estimating popu-lation parameters and production of fish in a tropical rain forest stream. North Venezuela. Hydrobiologia, 215 : 121-133.
Ponte V. 1990. — Recurso trôfico utilizado por peces juveniles en dos areas del Delta Inferior del rio Orinoco. Graduate thesis, Universidad Central de Venezuala, Caracas : 101 p. Provenzano F. & Castillo O. 1984. — La ictiofauna asociada
al fondo del rio Apure. I. Gymnotiformes. Acta Cient. Venez., 35 (Supl. 1) : 469 (abstract).
Provenzano F., Brull O . , Lasala J. & Castillo O. 1984a. — La ictiofauna asociada al fondo del rio Apure. II. Familia Pime-lodidae. Acta Cient. Venez., 35 (Supl. 1) : 469 (abstract). Provenzano F., Brull O., Lasala J. & Castillo O. 1984b. — La
ictiofauna asociada al fondo del rio Apure. III. Familia Lori-cariidae. Acta Cient. Venez., 35 (Supl. 1) : 469 (abstract). Provenzano F., Brull O . , Lasala J. & Castillo O. 1984c. — La ictiofauna asociada al fondo del rio Apure. IV. Familias Dora-didae, Ageneiosidae. Auchenipteridae, Hypophthalmidae, Cetopsidae y Bunocephalidae. Acta Cient. Venez., 35 (Supl. 1 : 469 (abstract).
Ramia M. 1972. — Cambios en la vegetaciôn de las sabanas del Hato El Frio (Alto Apure) causados por diques. Bol. Soc. Ven. Cienc. Nat., 124-125 : 57-80.
Ramos F., Novoa D. & Itriago I. 1982. — Resultados de los Pro-gramas de Pesca exploratoria efectuados en el Delta del noco. In D. Novoa (Comp.). Los Recursos Pesqueros del Ori-noco y su Explotacion. Corporaciôn Venezolana de Guayana, Editorial Arte, Caracas : 162-191.
Scharger R. & Gonzalez R. 1973. — Estudio Agrobiolôgico pre-liminar. Sectores Bruzual y Mantecal. M . O . P . Direcciân General de Recursos Hidraùlicos. Division Edafologia, Caracas.
Shannon G. & Weaver W. 1949. — The Mathematical Theory of Communication. Univ. Illinois Press, Urbana : 177 p. Sioli H. 1975. — Tropical rivers as expressions of their
terres-trial environments. In F.'Golley & E. Medina (Ed.). Tropi-cal EcologiTropi-cal System. Trends in Terrestrial and Aquatic Research. Springer-Verlag, Berlin : 275-288.
Stewart D., Barriga R. & Ibarra M. 1989. — Ictiofauna de la Cuenca del rio N a p o , Ecuador Oriental : Lista anotada de especies. Politecnica 12 (4) : 9-63.
Weitzman S.H. & Weitzman M. 1982. — Biogeography and evo-lutionary diversification in Neotropical freshwater fishes, with comments on the refugium theory. In G.T. Prance (Ed.). Bio-logical Diversification in the Tropics. Columbia University Press, New York : 403-422.
Welcomme R.L. 1979. — Fisheries Ecology of Floodplain Rivers. London, Longman : 317 p.
Welcomme R.L. 1985. — River fisheries. F A O Fisheries Tech-nical Paper, (262) : 330 p .