A REVIEW ON QUORUM SENSING INHIBITORS
Full text
Figure
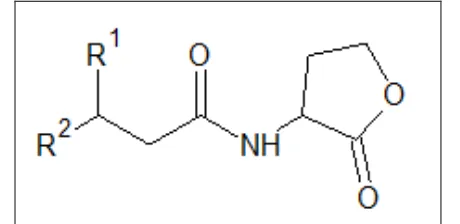

Related documents
The purpose of this paper is to prove a common fixed point theorem for hybrid contractions with generalized contractive conditions by using the concept of weakly compatible
mitigate the important power quality issues such as voltage sags, distortions, unbalanced voltages and single.. and three phase
Accordingly, [37] examined the effect of financial crisis, external debt management on the economic growth of Nigeria using GDP as endogenous variable while exogenous
Grounded on the respective analyses and explorations that have been made of the poetic narrations, poetic stories, the narrative discourses, and the non-narrative discourses
We aimed to determine the location of cellular DNA in non-degraded human teeth to quantify the yield of nuclear DNA from cementum, the most accessible and easily sampled dental
Next I shimmed the cabinet plumb and level, and joined the face frames with screws through predrilled holes.. I finished by screwing the cabinet to
The main contribution of this paper is to show how business process modelling (specifically proc- ess maps) can be used in order to develop such business process models that will
As she goes deeper in the hierarchy (Fig.10), she notices that temporal alignment starts fading out, and that sequential emerges as smaller directories are rendered.
