Expression of Id2 in the developing limb is associated with zones of active BMP signaling and marks the regions of growth and differentiation of the developing digits
Full text
Figure
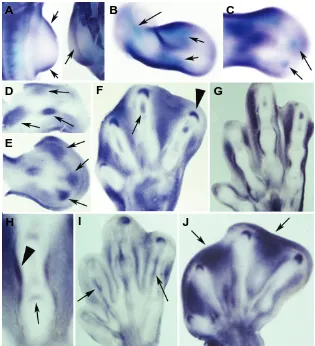
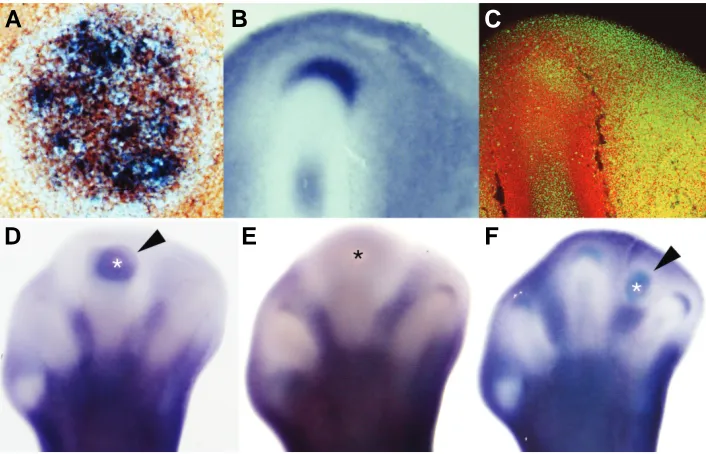
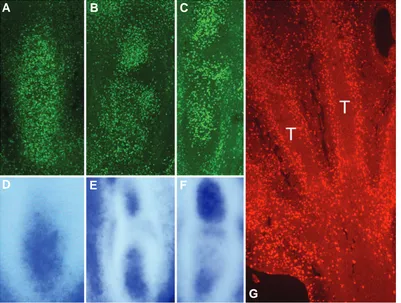
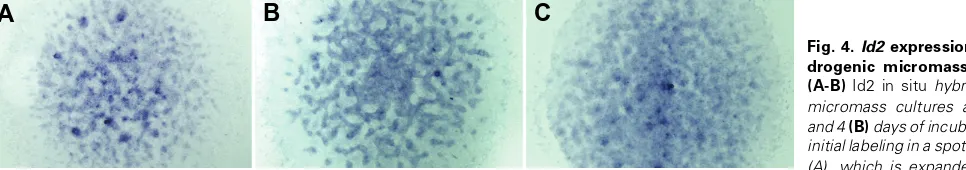
Related documents
The abundance of phage integrase-encoding genes was significantly increased in the viromes of medicated swine over that in the viromes of nonmedicated swine, demonstrating the
Since the bilayer-couple model predicts that spicule formation is the result of asymmetric expansion of the outer leaflet relative to the inner one, this effect should be
At 35 dpf, only three assemblages exhibited significantly different relative abundances in HF versus LF environmental microbiotas from tanks with fish, and all were enriched in the
In this regard, Farnsworth deviated from the customary restitution principle that "fault may be relevant to the measure of unjust enrichment and other features
Noting the features of the assignments, that they were intended to be absolute and that the lender was not required to take any action to en- force them, the
Of all the privacy issues that I have been watching through the privacy kaleidoscope during the past twenty-five years, none are more interesting and challenging,
It was decided that with the presence of such significant red flag signs that she should undergo advanced imaging, in this case an MRI, that revealed an underlying malignancy, which
Focus groups were conducted with partici- pants before each workshop to tailor the workshop con- tent to their needs; session content was adapted based on learning needs to make
