Copyright 0 1988 by the Genetics Society of America
Sequence, Genomic Distribution and DNA Modification of a
Mu1
Element
From Non-Mutator Maize Stocks
Vicki L. Chandler, Luther
E.Talbert’ and Fenella Raymond
Institute of Molecular Biology, University of Oregon, Eugene, Oregon 9 7 4 0 3
Manuscript received October 2, 1987 Revised copy accepted April 1 1, 1988
ABSTRACT
T h e increased mutation rate of Mutator stocks of maize has been shown to be the result of transposition of M u elements. One element, M u l , is present in 10-60 copies in Mutator stocks and approximately 0-3 copies in non-Mutator stocks. T h e sequence, structure and genomic distribution of an intact M u 1 element cloned from the non-Mutator inbred line B37 has been determined. T h e sequence of this element, termed Mul.4-B?7, is identical to M u 1 and it is flanked by 9-bp direct repeats indicative of a target site duplication. Mu1.4-B?7 is not in the same genomic location in all
stocks, which further suggests that it transposed into its genomic location in B37. We previously reported that in genomic DNA this element is modified such that certain methylation-sensitive restriction enzymes will not cut sites within the element. This is similar to that observed for M u
elements in Mutator stocks that have lost activity. We report herein that the Mul.4-B?7 element loses its modification and becomes accessible to digestion when placed in an active Mutator stock by genetic crosses. This suggests that factors conditioning unmodified elements are dominant in the initial cross between Mutator and non-Mutator stocks. In F 2 individuals that have subsequently lost Mutator activity the Mul.4-B?7 element again becomes modified as d o most of the M u elements in the stock. Thus, the modification state of the Mu1.4-B?7 element and the other Mul-like elements correlates with Mutator activity. We hypothesize that factor(s) within an active Mutator stock may inhibit the modification of M u elements, and that this activity is missing in non-Mutator stocks and may become limiting in certain Mutator stocks resulting in DNA modification.
M
CCLINTOCK (1 95 1) first proposed that transpos- able elements interact with structural genes to cause genetic instabilities in maize. Subsequent to McClintock’s report, transposable elements have been studied in many prokaryotic and eukaryotic orga- nisms. Transposable elements from a variety of species share many physical properties, such as inverted re- peat sequences at their termini and a duplication of a short sequence of target site DNA upon insertion. In addition, many transposable elements carry genes en- coding the proteins required for their transposition. Mutations are produced by an element’s insertion into or nearby a genetic locus, or by the induction of chromosome rearrangements (reviewed in SHAPIRO1983). For most transposable elements, the transpo- sition rate is low enough to be undetectable as an increased mutation rate. However, a few elements such as the
P
elements of Drosophila (ENGLES 1983), the Tcl elements of C. elegans (EMMONS et al. 1983), and the M u elements of the ROBERTSON’S Mutator stock of maize move at high enough frequencies in certain strains to increase the mutation rate.T h e ROBERTSON’S Mutator stock of maize was de- scribed approximately 10 yr ago (ROBERTSON 1978).
University, Bozeman, Montana 59717. Genetics 1 1 9 951-958 (August, 1988)
I Current address: Department of Plant and Soil Science, Montana State
When this stock was crossed to a standard stock with a normal mutation rate, approximately 90% of the F1 progeny retained the high mutation rate and new mutant alleles were recovered at frequencies ranging
from to (ROBERTSON 1978). This multifac-
tor segregation, in combination with the observation that approximately one-third of the new mutations isolated from Mutator stocks were unstable, suggested that Mutator activity was associated with an active transposable element family. Consistent with this hy- pothesis, several related transposable elements have been found as insertions in Mutator-induced muta- tions. A 1.4-kb insertion, designated M u l , was cloned from Mutator-induced mutations at A d h l (BENNETZEN et al. 1984), a1 (O’REILLY et al. 1985), and Bz2 (MCLAUGHLIN AND WALBOT 1987). A second cloned M u element is Mu1.7, which differs from M u 1 pri- marily d u e to an extra 380 bp of internal sequence (BARKER et al. 1984; TAYLOR, CHANDLER and WAL- BOT 1986). T h e terminal inverted repeats of M u l . 7
are 98% identical to the M u 1 termini and both ele- ments cause a 9-bp target site duplication upon inser- tion (TAYLOR and WALBOT 1987). Two additional elements were cloned as a result of their insertion into A d h l and waxy (CHEN et al., 1987; WESSLER, FLEENOR
952 Chandler. Talbert and F. Raymond
verted repeats of M u elements but their internal se- quences are unrelated to M u 1 and Mu1.7 or each other. The element at waxy has not been named; the element at A d h l is referred to as Mu?. It is not known which M u elements encode factors required for their transposition, or if additional factors are required.
One level of regulation is suggested by two reports of a correlation between the loss of Mutator activity and DNA modification of M u elements. An unstable allele of the bronze2 locus (bz2-mul) was isolated from a Mutator stock (WALBOT, CHANDLER and TAYLOR
1985; CHANDLER and WALBOT 1986). In this stock, Mutator activity is assayed by examining the ability of a M u 1 insertion at bronze2 to undergo somatic rever- sion, producing purple revertant spots on a bronze2 background. Lineages characterized by lowered activ- ity or no activity were derived from the original spotted kernel by identifying plants segregating few or no spotted kernels (WALBOT 1986). Southern blot analysis revealed that the inhibition of digestion of M u 1 elements by certain methylation-sensitive restric- tion enzymes correlated with the allele’s inability to undergo somatic reversion, producing a stable null phenotype at bz2-mul. T h e inhibition of digestion was shown to be the result of DNA modification, and while the exact nature of the modification is unknown,
it is most likely 5-methyl deoxycytosine. Similarly, BENNETZEN (1985, 1987) reported a correlation be- tween the methylation of internal restriction sites of M u 1 elements and the loss of Mutator activity as measured by forward mutation rate in inbred Mutator stocks (ROBERTSON 1986). However, other mecha- nisms for losing Mutator activity must exist since stocks have been identified that have no Mutator activity, yet contain unmodified M u elements (BEN- NETZEN 1987).
Standard stocks of maize whose pedigrees show no known mutator ancestors contain sequences with ho- mology to M u elements (CHANDLER, RIVIN and WAL- BOT 1986). We refer to these stocks as non-Mutator. Three types of homologous sequences have been iden- tified. First, intact M u l - or M u l . 7-like elements have been found in some inbred lines, but not in others. Second, all maize stocks examined contain a sequence 97% identical to internal portions of M u 1 and M u l . 7
elements that is not associated with M u termini (TAL- BERT and CHANDLER 1988). Third, all stocks exam- ined have approximately 40-50 copies of sequences homologous to the M u terminal repeats that are not associated with M u 1 or M u I . 7 internal sequences (CHANDLER et al. 1988).
There is no genetic evidence of a high mutation rate in these non-Mutator stocks, suggesting the ele- ments are not transposing at detectable frequencies. This lack of activity in non-Mutator stocks could be because the elements have accumulated nucleotide
changes or deletions. Alternatively, the elements may be intact but the small number of elements, the pres- ence of DNA modification or the absence of positive regulatory factors in the stock may prevent transpo- sition at a detectable frequency. To address these hypotheses we have determined the DNA sequence of a Mul-like element, designated Mul.4-B?7, which was cloned from the maize inbred line B37. We have compared the sequence of Mu1.4-B?7 with the se- quence of the M u 1 element that inserted into the A d h l gene (BENNETZEN et al. 1984). We have also examined the distribution of Mu1.4-B?7 in non-Mu- tator maize stocks, and analyzed the DNA modifica- tion of restriction sites within Mu1.4-B?7 in the pres- ence and absence of Mutator activity.
MATERIALS AND METHODS
Plant materials: Maize stocks were obtained from several sources as previously reported (CHANDLER, RIVIN and WAL-
BOT 1986). All stocks were propagated by hand self-polli-
nation either in the field or the greenhouse with supple- mental lighting in Eugene, Oregon.
DNA samples: The M u 1 plasmid, pMJ9, was obtained from J. BENNETZEN (BENNETZEN et al. 1984). T h e M u inter-
nal probe A/B5 was prepared from pMJ9 as previously described (CHANDLER, RIVIN and WALBOT 1986).
T h e endogenous Mu element, Mu1.4-B37, was isolated from a X phage library of total maize DNA from the inbred
line B37 as previously described (CHANDLER, RIVIN and WALBOT 1986). The 3.1-kbp EcoRI fragment containing
Mu1.4-B37 and flanking DNA (Figure 3) was subcloned into the EcoRl site of pUC8 (VIERRA and MESSING 1982).
Maize DNA was isolated from the cobs of immature ear shoots and purified by centrifugation in CsCl/ethidium bro- mide (RIVIN, ZIMMER and WALBOT 1982). Southern blot hybridizations were performed as previously described (CHANDLER, RIVIN and WALBOT 1986).
DNA sequencing: Restriction fragments of Mu1.4-837
were subcloned into M13 vectors (MESSING 1983) and se- quenced by the dideoxy chain termination method (SANGER, NICKLEN and COULSEN 1977) using the 17-mer universal sequencing primer (Pharmacia). To eliminate band compression which occurred when sequencing GC-rich re- gions, 7-deaza-dGTP (Boehringer Mannheim Biochemicals) was used in place of dGTP (MIZUSAWA, NISHIMURA and SEELA 1986). Sequence comparisons were made using the University of Wisconsin Genetics Computer Group software and documentation package (DEVEREUX, HAEBERLI and SMITHIES 1984).
RESULTS
Endogenous Mu1 Element 953
M H T M N A B E E E D l l l M T H M I oobp
I
- -
I I l l I I "II I V 9 I I H"
" "
-
.
"c c
-
c_
-
c *
FIGURE 1 .-Restriction map and subcloning strategy for Mul.4- B37. Mul-homologous DNA is indicated by bar. The arrows directly under the map indicate the inverted and direct repeats. The restric- tion sites indicated are M: Mlul, H: H i d , T: Tth 1 1 1-1, N: N o t l , A:
A v a I , B: BglI, E: BstEII, D: DdeI, I: BstNI. The arrows below the map indicate the direction and extent of the subclones sequenced.
(Figure 2). We sequenced the regions of M u 1 that showed differences relative to Mul.4-B37 using the same sequencing strategy employed for M u l . 4-B37. Our sequencing shows that both M u 1 and Mu1.4-B37 have the GC dinucleotide at positions 453 and 454 and A's at positions 1086 and 1092 as shown in Figure 2. Thus, there are no differences between the two elements when they are sequenced and compared on the same sequencing gel. T h e GC dinucleotide not previously detected slightly alters two of the reading frames as drawn in BARKER et al. (1 984). T h e revised reading frames are shown in Figure 2. T h e sequence identity of Mu1.4-B37 and M u 1 suggests that Mu1.4- B37 should be as capable of transposition as M u l . Consistent with this, Mul.4-B37 is flanked by a 9-bp direct repeat (Figure 2) as has been determined for all sequenced M u element insertions.
We previously reported that restriction mapping of the cloned elements detected one restriction site dif- ference between M u 1 and Mu1.4-837, in that Mu1.4- B37 appeared to be missing a CZaII site relative to M u 1 (CHANDLER, RIVIN and WALBOT 1986). How- ever, the DNA sequence of both elements shows three recognition sites for this enzyme in this region. T h e different restriction patterns may be the result of a differential DNA modification that occurred during growth of the cloned elements in Escherichia coli. In the previous experiment the M u 1 and Mu1.4-B37 elements were in different plasmid vectors and were propagated in different E. coli hosts. When both plas- mids are grown in the same host cells, no restriction site polymorphism is detected (data not shown).
Mu1.4-B37 has the distribution of a transposable element: T h e identity of Mul.4-B37 and M u l , and the fact that Mul.4-B37 appears to be flanked by a target site duplication, suggests that this element in- serted into its current genomic location. We showed previously that the inbred lines B37, W23 and A1 88, as well as an individual from the variety Wilbur's Knobless Flint, all contain Mul-like elements; inbred lines W22 and 1S2P contain an element resembling M u l . 7 , while B73 and Ky21 contain no intact M u l - or M u l . 7-like elements (CHANDLER, RIVIN and WAL- BOT 1986). If the MuI-like element in B37, W23, A188 and Wilbur's Knobless Flint is inserted in the
cctCaCOatGAGATMTTGCCATT~GGACGMGAGGGAAGGGGATTCGACGAAATGGAG
O C ~ ~ O T T O G C I T C T C ~ G G A G A C ~ C G C G A C A G C C M A C T C C A A M C G G A T
A C G A G A C A G C T ~ G G O G C G T A A A C A ~ A ~ A ~ C T G T C C C C ~ A C C G T T C
CCGTGCGCAOACGCCGTCCGCGTACT~CTTGTCTCCGTCGCCGCGCTCTACGGCAGC 1.
ACGGCWTGCTGGCGTACGTGCTGTACTCCGCGCCGGCAGAGGCCGCGCGCGCGTCGCCC
GCGGACGACGCCACGCCGCCACCTCGCCGCGGCCGCGCACATCCTCCTCGCCGCGAGC
GCGTCCGCGGACGCGCGCGCCTTCTCGGGCCTCCACGCCGGGGACCTCCTCCTCCCGCGC
GCCMCGAGGCCGCCGCCGCCGCGGACCGGGCGCTCCGCGTGCTCCTCM~CCCCGCG AA
GTGTCACTGTCGTCCTCGCCCAAGAAGTCCGCCTCGCCGCCGCCGGCCCCGG4GGCGAGG
MGGAGTACCCGCCCGAC~GACGCTGCCGGACCTCAAGAGCGGGCT~CAGCACCGAC
G A G C A A C G G A C G C A C C T T C C A C C T G T C C T A G C A ~ A A T T C A C 4
C M A ~ C A A ~ ~ ~ ~ A G G T I T T G G T A ~ A ~ - A ~ A A G A C T A A A A G ~ A T ~ ~ A 4
CTCGTkmGCC~:GGMIITATTGCCAAM~G~ACCGAAGG~~CC~AT~AAAAATGGC
GGTATCCGTCCACCTGTAGCTTGTCCMGGCTCTTCCAGCAGAGTAGTGCCGCCATGACG
ATTGACAGAGACACGAGACGAAACAAGCTGAAGGTCCCCGCGGCGTCACTGTCGTC~CG
CCGMGMGTCCGCCTCGCCGCCGCCGGCCCCGGAGGCGAGGAAGGAGTACCCGCCCGAC
CTGACGCTGCCCACCGTGTCGCCGCCGCCGCCCAACGGCCTCGGCGACATGCTCAGCCCA
GCGGCCTGGCCCTCCTCCCCCGCGAGCAGGCTCAACAAGGCCGCGCTCGGCGGCGGCCGG
G A C C T G G A C C T ~ C C A G T A C C A G C A ~ ~ ~ ~ C G A C A A G G T ~ ~ G T ~ G T ~ G A G G G C ~
A G T A G G A G A C M G M G A G T A C G C ~ ~ ~ C T G A G A T G ~ G A ~ G G A G M A A A G G G T A ~ G ~ ~ G A
A A
CGGCGTCTGPACACGGGMCGGTAAATGAGGACAGGMAC"MCTGTTTACGCAGCC
CCMGTGCTGTCTTGTGTCCGTITTAGAGTTTGGCTGTCGCGTACGTCTCTAAAACAGAG
A A G C C A A C O C C A A C G C C T C A ~ G T C G A A T C C O C f i C I A
TTATCTCcctcacwt
120
240
360
480
600
720
840
960
1080
1200
1320
FIGURE 2.-DNA sequence of Mu-homologous element Mu1.4- B37. The 9-bp direct repeat flanking the element is shown in lower case. The four major open reading frames are shown above the sequence and are numbered as published in BARKER et al. 1984.
The regions of difference with respect to the published sequence of Mul are indicated by arrows.
same genomic location, we would expect the DNA sequences flanking the Mul.4-B37 element to cohy- bridize to a restriction fragment with the M u 1 probe in each of these stocks. To test this idea, the DNA sequences flanking Mul.4-B37 were used as hybridi- zation probes on Southern blots of DNA samples from several maize inbred lines and varieties (Figure 3, A-
L. E.
3.1 3.1
t7
3.1
1.7
D F B P Y PY T C. I T Y T R H
-
”LEFT FLANKING
uul
FLANKING RIGHT
-
m sFIGURE 3.-Genomic distribution of Mu1.4-B37 in non-Mutator maize stocks. A, EcoRI digests of DNA (3-6 ag) from non-Mutator maize stocks. Southern blot probed with Mu1 internal probe. Lanes a-h: non-Mutator inbred lines and varieties of maize; a, 1S2P b, W23; c, W22; d, Ky21; e, A188; f, B73; g, B37; h, Wilbur’s Knobless Flint. B, Same blot hybridized to M u 1 . 4 3 3 7 right flanking probe, which recognizes the cloned EcoRI fragment and one additional fragment in each stock, although the intensity of the additional fragments varies. C, Same blot except hybridized to Mu1.4-B37 left flanking probe. D, Partial restriction map of Mu1.4-B37 and flank- ing DNA showing regions of homology to the probes. The restric- tion sites indicated are R: EcoRI, H: Hinfl, B BamHI, T : Tthl 1 1-
I , A: AvaI, 1: BstNI. Only the AvaI and BslNI sites within the Mu
element are indicated; there are additional AvaI and BstNI sites in the flanking DNA.
this fragment hybridizes to both the MuZ internal probe (Figure 3A, lanes b and g) and to DNA se- quences flanking both sides of the Mu1.4-B37 element (Figure 3, B and C, lanes b and g). None of the other inbred lines and varieties contains a M u element in this fragment. Instead, most stocks contain a 1.7-kbp fragment that hybridizes to the probes flanking MuZ.4-B37 (Figure 3, Band C), but does not hybridize to the MuZ internal probe (Figure 3A). This 1.7-kbp fragment likely represents the 3.1-kbp fragment of B37 without the 1.4-kbp M u element. The inbred line A 188 and variety Wilbur’s Knobless Flint also contain intact MuZ-like elements. However, in these stocks the elements appear to be in different genomic locations since they are not flanked by the same DNA sequences as MuZ.4-B37 (Figure 3, A-C, lanes e and h).
MuZ.4-B37 appears to have transposed into its cur- rent genomic location, yet there is no genetic evidence that the element is currently active. B37 and W23 have the same element in the same genomic location, yet pedigree analysis reveals that W23 and B37 have no recent common ancestor (W. F. TRACY, personal communication). The Golden Glow stock from which W23 was derived was examined and found to contain the 3.1-kbp EcoRI fragment characteristic of Mu1.4- B37 (data not shown). Golden Glow and W23 are
separated by approximately 40 years, yet the element is in the same location and has not increased in copy number in either stock. This further suggests that MuZ.4-B37 is not as active as MuZ elements in Mutator stocks.
DNA
modification of the MuZ.4-B37 element is lost in active Mutator stocks: In B37 genomic DNA, the termini of MuZ.4-B37 are modified such that the Hinfl sites are not accessible to digestion (CHANDLER,RIVIN and WALBOT 1986). This is similar to M u ele- ments in certain Mutator stocks that have lost activity (CHANDLER and WALBOT 1986). Thus, loss of modi- fication may be a prerequisite for activity of endoge- nous M u elements. T o determine if the modification
of the Hinfl sites in MuZ.4-B37 is stable in the presence of Mutator activity, a W23 stock homozygous for the MuZ.4-B37 element was crossed to an active Mutator stock. The non-Mutator W23 stock was used as a parent for two reasons. First, the same MuZ element is inserted into the same EcoRI fragment in both W23 and B37 (Figure 3, A-C, lanes b and g). Second, the W23 stock was recessive for bz2 which allowed us to score for activity of a Mu1 element at the bronze2 gene (MCLAUGHLIN and WALBOT 1987) that was intro- duced from the Mutator parent. When the MuZ ele- ment at bz2 is active, somatic reversion occurs pro- ducing purple revertant sectors, and when the ele- ment is inactive, stable bronze kernels are observed (WALBOT 1986). The element’s loss of activity has been correlated with DNA modification of the MuZ elements in the stock (CHANDLER and WALBOT 1986). In this experiment, DNA was prepared from the non- Mutator W23 stock and several F1 and FP progeny, and the accessibility of the MuZ.4-B37 HinfI sites examined utilizing Southern blots. As shown in Figure
4, A and B, the sequences flanking MuZ.4-B37 are fused to the element in DNA from W23 plants (lane a), resulting in 2.5- and 2.2-kbp fragments that hybri- dize to both M u and flanking DNA probes. This demonstrates the fusion of the element to its flanking DNA and represents the masking of the MuZ.4-B37 Hinfl sites in W23. The same result was previously described in the non-Mutator line B37 (CHANDLER,
RIVIN and WALBOT 1986). In contrast, in the FI progeny of an active Mutator stock crossed to W23, the DNA sequences flanking MuZ.4-B37 are no longer contiguous to the M u element since digestion and hybridization to the flanking DNA results in the dis- appearance of the 2.5- and 2.2-kbp fragments and the appearance of the 0.95-kb fragment (Figure 4B, lanes b-d) predicted from the restriction map (Figure 3D). This suggests that the HinfI sites are no longer masked and are now accessible to digestion. The F1 progeny were then selfed and F2 individuals screened for loss
Endogenous Mu1 Element
* a b c d e f g 'a b c d e f g
-"
0
-
.C-r
2 5- 2.5-
2.2- 22-
e w . 7
.
"d.95-FIGURE 4.-Modification state of M u l . 4 - 8 3 7 in non-Mutator and Mutator backgrounds. A, Hinfl digestion ofmaize DNA. South- ern blot hybridized to Mu1 internal probe. Lane a: W23 non- Mutator stock. Lanes b, c, and d: F1 progeny from the cross of W23
by an active Mutator stock. Lanes e and f: FP progeny of W23/
Mutator cross that have lost Mutator activity. Lane g: Hinfl-re- stricted Mu1 and Mu1.7 plasmid DNA at three and one copies per
haploid genome, respectively. B, Same blot hybridized to Mu1.4- B37 right flanking probe. The location of Hinfl sites and regions of homology to the probes are shown in Figure 3. The fragments corresponding to the cloned element are indicated with arrows. Both the M u and right flanking probes hybridize to additional fragments when either the Hinfl and EcoRI enzymes are used (Figure 3).
reversion (CHANDLER and WALBOT 1986). DNA sam- ples from two such individuals are shown in Figure 4. Most of the Mu elements are modified as shown by the appearance of restriction fragments hybridizing to the Mu probe larger than the 1.3-kb fragment characteristic of Mul-like elements (Figure 4A, lanes e and f ) . T h e Mu1.4-B37 element is also modified as indicated by the disappearance of the 0.95-kbp flank- ing fragment and the appearance of the 2.5- and 2.2- kbp fusion fragments (Figure 4B, lanes e and f ) .
Fusion fragments larger than 2.5-kbp are not ob- served, demonstrating that the HinfI sites in the flank- ing DNA are cleaved. In addition, the ratios of the various fusion fragments varies between individual plants, showing that the modification of both sites within the element does not always occur. In FP prog- eny that retained Mutator activity the HinfI sites in Mu1.4-B37 and all other Mu elements remain acces- sible to digestion (data not shown). Thus, the accessi- bility of HinfI sites in all Mu elements correlates with Mutator activity. T h e large number of Mu elements in the selfed progeny is typical after self-pollination of Mutator plants (ALLEMAN and FREELINC 1986).
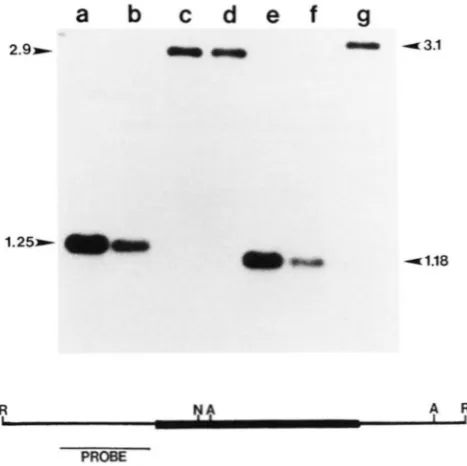
T h e accessibility of other methylation-sensitive re- striction sites within Mu1.4-B37 was examined in the different genetic backgrounds. An experiment show- ing AvaI and Not1 digestion of genomic DNA samples is in Figure 5. These sites are accessible to digestion in DNA from the non-Mutator line (lanes a and e) and in the active Mutator stocks (lanes b and f). However, the sites are masked in Mutator plants that have lost activity as demonstrated by the larger fragments in
a
2.9-
1.25-
R
955
b
c
d
e
f
g
- 0
-
4 3 . 141.18
NA A. R..
PROBE
FIGURE 5.-Modification state of internal Mu1.44337 restriction sites. Southern blots of genomic DNA restricted with either EcoRI and AvaI (lanes a-d) or EcoRl and Not1 (e-g) and hybridized with
Mu1.4-B37 left flanking probe as shown on map. The Mutator plant was in a W23 background so all the plants were homozygous for the Mu1.4-B37 element. Lanes a and e: W23 DNA. Lanes b and f:
FI individual from Mutator X W23 (non-Mutator) cross. Lanes c, d and g: Fn individuals that lost Mutator activity. A partial restriction map of Mu1.4-B37 (thickened region) and flanking region showing the location of EcoR1, AuaI and Not1 sites is shown below the blot.
The sizes of the homologous fragments are indicated in kbp.
lanes c, d, and g. Similar results were obtained with four other enzymes, EcoRII, NcoI, BglI and MluI; the sites are accessible to cleavage in the non-Mutator line and active Mutator plants, but are not accessible in inactive Mutator plants. Thus, of the enzymes tested only the HinfI sites of Mu1.4-B37 are modified in non- Mutator DNA, no sites are modified in active Mutator stocks, and all methylation-sensitive sites tested are modified in Mutator stocks that have lost activity.
These results show that the DNA modification of the Mul.4-B37 element changes, depending on the genetic background of the stock. In addition, the modification of Mu1.4-B37 parallels the modification of the majority of Mu elements in the Mutator stock, suggesting that the element responds to regulatory factors in a manner similar to other Mu elements. We have examined a total of 45 F1 individuals from crosses of W23 with four different active Mutator stocks and the results were identical to those shown in Figure 4. T h e Mu1.4-B37 element lost its modifi- cation independent of whether the Mutator plant was utilized as male or female.
DISCUSSION
956 V. L. Chandler, L. E. Talbert and F. Raymond
bp element. Mu1.4-B37 is flanked by 9-bp direct re- peats as are M u l , M u l . 7 and M u 3 elements (BARKER et al. 1984; TAYLOR and WALBOT 1987; CHEN et al.
1987) indicating that it inserted into its genomic lo- cation in B37. However, there is no evidence that Mu1.4-B37 is currently active. T h e termini of many transposable elements contain the cis-acting sites re- quired for transposition. For example, it appears that sequences surrounded by the Ac/Ds terminal repeats can be induced to transpose if Ac is present to provide the necessary enzymes for excision and transposition (reviewed in DORING and STARLINGER 1986). If this is the case for the Mutator system, M u l . 4 - B 3 7 might be activated to transpose if supplied with the necessary transacting factors.
T h e M u l . 4 B 3 7 element normally exists in a modi- fied state in the inbred lines W23 and B37, and we previously hypothesized that this may contribute to its inactivity (CHANDLER, RIVIN and WALBOT 1986). Given the correlation between DNA modification of M u elements and their lack of activity in Mutator stocks, a necessary prerequisite for the activation of Mu1.4-B37 may be that it loses its DNA modification. W e have demonstrated that when Mu1.4-B37 is intro- duced into an active Mutator stock by genetic crosses its DNA is no longer modified. T h e Mul.4-B37 ele- ment remains unmodified in subsequent generations when the Mutator stocks remain active, and in Muta- tor stocks that subsequently lose activity Mul.4-B37 again becomes modified as do all the M u elements in the stock. However, there is a major difference be- tween the modification state o f M u l . 4 - B 3 7 in the non- Mutator W23 stock and its modification state in Mu- tator stocks that have recently lost activity. In the inactive Mutator stocks all
Mu
elements including Mu1.4-B37 are modified at numerous internal sites as well as terminal sites, whereas in non-Mutator lines the only sites modified in Mu1.4-B37 are the Hinf I sites in the termini. T h e factors influencing this dif- ferential modification and its effects on the cis- and trans-acting element functions remain to be deter- mined. It is possible that the differential modification reflects different levels of inactivation, similar to that observed with the Spm elements (FEDOROFF et al.Factors conditioning unmodified elements appear to be dominant in the initial crosses between Mutator and non-Mutator stocks. This suggests that some ac- tivity within a Mutator stock may be responsible for preventing the modification of M u elements. This could result from the presence of a demethylation enzyme in active Mutator stocks, or simply by the prevention of methylation of M u elements after DNA
replication by blocking the action of a methylase. This hypothesis predicts that this activity would be missing in non-Mutator lines, and can become limiting in
1988).
certain Mutator stocks resulting in DNA modification of M u elements. T h e fact that M u elements may become modified after self pollination (CHANDLER and
WALBOT 1986) or inbreeding of Mutator stocks (BEN- NETZEN 1985, 1987) suggests that Mutator stocks are not simply missing a methylation activity present in non-Mutator stocks. Since the presence of modified M u elements is only correlated with lack of activity we do not know whether DNA modification is the pri- mary mechanism of inactivation, is directly involved in maintenance of the inactive state, or if inactive elements become more easily modified by normal cellular activities.
M u elements are not the only transposable elements whose activities are regulated by DNA modification: the transposition of the prokaryotic transposable ele- ment IS10 is regulated by adenosine methylation
(ROBERTS et al. 1985) and the activities of the maize transposable elements Ac/Ds and Spm ( E n ) are corre-
lated with hypomethylation (DELLAPORTA and
CHOMET 1985; SCHWARTZ and DENNIS 1986;
CHOMET, WESSLER and DELLAPORTA 1987; CONE, BURR and BURR 1986; FEDOROFF et al. 1988; RABOY, SCHIEFELBEIN and NELSON 1988). Only active Ac, Spm, and dSpm elements are hypomethylated, with the bulk of the other cross-hybridizing sequences in the ge- nome remaining extensively methylated. This fact has been successfully exploited to separate the active ele- ments from inactive sequences facilitating the cloning of several genes containing Ac or Sprn elements (CONE, BURR and BURR 1986; CHEN, GREENBLATT and DEL- LAPORTA 1987; SCHMIDT, BURR and BURR 1987). In contrast, the majority of M u elements in the genome are unmethylated in active stocks and methylated in inactive stocks. This may mean that most of the M u elements are active in Mutator stocks, while only a small number of Ac or Spm elements are active in Ac and Spm stocks, respectively. It is also possible that the differences in the methylation-demethylation patterns between M u elements and Ac and Spm may represent different methylation pathways. It remains to be de- termined for all the maize elements whether the fac- tors mediating methylation are general cellular en- zymes or element encoded.
Endogenous Mu1 Element 957
rounds of DNA replication hypomethylated elements would occur which might be sufficient for activation. Transposable elements were first detected in stocks
undergoing chromosome breakage (MCCLINTOCK
1984), and recently a cryptic Ac element was activated during tissue culture (PESCHKE, PHILLIPS and GEN- GENBACH 1987). Other environmental and physiolog- ical conditions have been correlated with increased mutability in plants and have been hypothesized to activate transposable elements (MCCLINTOCK 1978; BURR and BURR 198 1). Thus, the liberation of cryptic transposable elements may play an important role in genome stability, and DNA methylation may be a mechanism for heritably, but reversibly controlling element transposition.
We wish to thank DEVON TURKS for her excellent technical assistance, CAROL RIVIN a n d o u r colleagues for their comments on the manuscript, ELIZABETH COOKSEY for her editorial comments and help in manuscript preparation, and the maize geneticists and breeders for providing the stocks analyzed in this investigation. This research was supported by a National Science Foundation grant (DCE-8451656) and matching funds from Pioneer Hi-Bred International, CIBA-GEIGY, and Northwest Area Foundation to V.L.C.
LITERATURE CITED
ALLEMAN, M., AND M. FREELING, 1986 T h e M u transposable elements of maize: evidence for transposition and copy number regulation during development. Genetics 112: 107-1 19.
BARKER, R. F., D. V. THOMPSON, D. R. TALBOT, J. SWANSON and J. L. BENNETZEN, 1984 Nucleotide sequence of the maize
transposable element M u l . Nucleic Acids Res. 12: 5955-5967.
BENNETZEN, J., 1985 T h e regulation of Mutator function and Mu1
transposition. UCLA Symp. Mol. Cell. Biol. 35: 343-353.
BENNETZEN, J. L., 1987 Covalent DNA modification and the regulation of Mutator element transposition in maize. Mol. Gen. Genet. 208: 45-51.
BENNETZEN, J. L., J. SWANSON, W. C. TAYLOR and M. FREELING,
1984 DNA insertion in the first intron of maize A d h l affects message levels: cloning of progenitor and mutant A d h l alleles. Proc. Natl. Acad. Sci. USA 81: 4125-4128.
BURR, B., and F. BURR, 1981 Transposable elements and genetic instabilities in crop plants. Stadler Genet. Symp. 13: 115-128.
CHANDLER, V., and V. WALBOT, 1986 DNA modification of a maize transposable element correlates with loss of activity. Proc. Natl. Acad. Sci. USA 83: 1767-177 1.
CHANDLER, V., C. RIVIN and V. WALBOT, 1986 Stable non-Mu- tator stocks of maize have sequences homologous to the Mu1
transposable element. Genetics 114: 1007-1021.
CHANDLER, V. L., L. E. TALBERT, L. MANN and C. FABER,
1988 Structure and DNA modification of endogenous M u
elements. pp. 339-350. In: Plant Transposable Elements, Edited by 0. E. NELSON, JR. Plenum Press, New York.
CHEN, J., I . M. GREENBLATT and S. L. DELLAPORTA,
1987 Transposition of Ac from the P locus of maize into unreplicated chromosomal sites. Genetics 117: 109-1 16.
CHEN, C-H., K. K. OISHI, B. KLOECKENER-GRUISSEM and M. FREEL-
ING, 1987 Organ-specific expression of maize A d h l is altered after a M u transposon insertion. Genetics 116: 469-477.
CHOMET, P. S., S. WESSLER and S. L. DELLAPORTA,
1987 Inactivation of the maize transposable element Activator ( A c ) is associated with its DNA modification. EMBO J. 6 295-
302.
CONE. K. C., F. BURR a n d B. BURR, 1986 Molecular analysis of
the maize anthocyanin regulatory locus C I . Proc. Natl. Acad. Sci. USA 83: 9631-9635.
DELLAPORTA, S. L., and P. S. CHOMET, 1985 T h e activation of maize controlling elements. pp. 170-217. In: Plant Gene Re- search: Genetic Flux in Plants, Edited by B. HOHN and E. S. DENNIS. Springer-Verlag, New York.
DEVEREUX, J. R., P. HAEBERLIS and 0. SMITHIES, 1984 A corn- prehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12: 387-395.
DORING, H.-P., and P. STARLINGER, 1986 Molecular genetics of transposable elements in plants. Annu. Rev. Genet. 20: 175- 200.
EMMONS, S. W., L. YESNER, K. RUAN and D. KATZENBERG,
1983 Evidence for a transposon in Caenorhabditis elegans. Cell
32: 55-65.
ENGELS, W. R., 1983 T h e P family of transposable elements in Drosophila. Annu. Rev. Genet. 17: 315-344.
FEDOROFF, N., P. MASSON, J. BANKS and J. KINGSBURY,
1988 Positive and negative regulation of the Suppressor-mu- tator elements. pp. 1-15. In: Plant Transposable Elements, Ed- ited by 0. E. Nelson, Jr. Plenum Press, New York.
MCCLINTOCK, B., 1951 Chromosome organization and genic
expression. Cold Spring Harbor Symp. Quant. Biol. 16: 13- 47.
MCCLINTOCK, B., 1978 Mechanisms that rapidly reorganize the genome. Stadler Genet. Symp. 1 0 25-48.
MCCLINTOCK, B., 1984 T h e significance of responses of the ge- nome to challenge. Science 226: 792-801.
MCLAUGHLIN, M., AND V. WALBOT, 1987 Cloning of a mutable
bz2 allele of maize by transposon tagging and differential hybridization. Genetics 117: 771-776.
MESSING, J., 1983 New M13 vectors for cloning. Methods En- zymol. 101: 20-78.
MIZUSAWA, S., S. NISHIMURA and F. SEELA, 1986 Improvement of the dideoxy chain termination method of DNA sequencing by use of deoxy-7-deazaguanosine triphosphate in place of dGTP. Nucleic Acids Res. 14: 13 19- 1324.
O'REILLY, C., N. S. SHEPHERD, A. PEREIRA, 2. SCHWARZ-SOMMER, I. BERTRAM, D. S. ROBERTSON, P. A. PETERSON and H. SAE-
DLER, 1985 Molecular cloning of the a 1 locus of Zea mays
using the transposable elements En and M u l . EMBO J. 4: 877- 882.
PESCHKE, V. M., R. L. PHILLIPS and B. G. GENGENBACH,
1987 Discovery of transposable element activity among prog- eny of tissue culture-derived maize plants. Science 238: 804- 806.
RABOY, V., J. W. SCHIEFELBEIN and 0. E. NELSON, 1988 DNA modification and the timing of defective Spm excision from the
br-m13 allele in maize. pp. 376-377. In: Plant Transposable Elements, Edited by 0. E. NELSON, JR. Plenum Press, New York.
RIVIN, C. J., E. A. ZIMMER and V. WALBOT, 1982 Isolation of DNA and DNA recombinants from maize. pp. 161-164. In:
Maize f o r Biological Research, Edited by W. F. SHERIDAN. Plant Molecular Biology Association, Charlottesville, Va.
ROBERTS, D., B. C. HOOPES, W. R. MCCLURE and N. KLECKNER,
1985 IS10 transposition is regulated by DNA adenine meth- ylation. Cell 43: 1 17-1 30.
ROBERTSON, D. S., 1978 Characterization of a Mutator system in maize. Mutat. Res. 51: 21-28.
ROBERTSON, D. S., 1986 Genetic studies on the loss of M u mutator activity in maize. Genetics 113: 765-773.
SANGER, F., S. NICKLEN and A. R. COULSON, 1977 DNA sequenc- ing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 74: 5463-5467.
SCHMIDT, R. J., F. A. BURR and B. BURR, 1987 Transposon tagging and molecular analysis of the maize regulator locus
SCHWARTZ, D., and E. DENNIS, 1986 Transposase activity of the Ac controlling element in maize is regulated by its degree of
methylation. Mol. Gen. Genet. 205: 476-482.
SHAPIRO, J., 1983 Mobile Genetic Elements. Academic Press, New York.
TALBERT, L. E., and V. L. CHANDLER, 1988 Characterization of a highly conserved sequence related to Mutator transposable elements in maize. Mol. Biol. and Evol., In press.
TAYLOR, L. P., A N D V. WALBOT, 1987 Isolation and characteriza- tion of a 1.7 Kb transposable element from a Mutator line of maize. Genetics 117: 297-307.
TAYLOR, L., V. CHANDLER and V. WALBOT, 1986 Insertion of
1.4 Kb and 1.7 Kb M u elements into the bronze-1 gene of Zea mays L. Maydica 31: 31-45.
VIERRA, J., A N D J. MESSING, 1982 T h e p U C plasmids, an M13mp7-derived system for insertion mutagenesis and se- quencing with synthetic universal primers. Gene 19: 259-268. WALBOT, V., 1986 Inheritance of mutator activity in Zea mays as
assayed by somatic instability of the br2-mu1 allele. Genetics WALBOT, V., V. CHANDLER and L. TAYLOR, 1985 Alterations in the mutator transposable element family of Zea mays. UCLA Symp. Mol. Cell. Biol. 35: 333-342.
WESSLER, S. R., D. FLEENOR and M. VARAGONA, 1988 T h e splic- ing of transposable elements and DNA insertions from Wx
mRNA. pp. 293-303. In: Plant Transposable Elements, Edited by 0. E. NELSON, JR. Plenum Press, New York.
114: 1293-1312.