Copyright 0 199 1 by the Genetics Society of America
Genetic Exchange Across
a Paracentric Inversion of the Mouse t Complex
M. F.
Hammer,’ S. Bliss and L. M. Silver
Department of Molecular Biology, Princeton University, Princeton, New Jersey 08544 Manuscript received January 15, 199 1
Accepted April 27, 199 1
ABSTRACT
Mouse t haplotypes are distinguished from wild-type forms of chromosome 17 by four non-
overlapping paracentric inversions which span a genetic distance of 20 cM. These inversion polymor- phisms are responsible for a 100-200-fold suppression of recombination which maintains the integrity of complete t haplotypes and has led to their divergence from the wild-type chromosomes of four species of house mice within which t haplotypes reside. As evidence for the long period of recombi- national isolation, alleles that distinguish all t haplotypes from all wild-type chromosomes have been established at a number of loci spread across the 20-cM variant region. However, a more complex picture emerges upon analysis of other t-associated loci. In particular, “mosaic haplotypes” have been identified that carry a mixture of wild-type and t-specific alleles. T o investigate the genetic basis for
mosaic chromosomes, we conducted a comprehensive analysis of eight t complex loci within 76 animals
representing 10 taxa in the genus Mus, and including 23 previously characterized t haplotypes. Higher resolution restriction mapping and sequence analysis was also performed for alleles at the Hba-ps4 locus. The results indicate that a short tract of D N A was transferred relatively recently across an inversion from a t haplotype allele of Hba-ps4 to the corresponding locus on a wild-type homolog leading to the creation of a new hybrid allele. Several classes of wild-type Hba-ps4 alleles, including
the most common form in inbred strains, appear to be derived from this hybrid allele. The accumulated
data suggest that a common form of genetic exchange across one of the four t-associated inversions is
gene conversion at isolated loci that do not play a role in the transmission ratio distortion phenotype required for t haplotype propagation. The implications of the results pose questions concerning the evolutionary stability of gene complexes within large paracentric inversions and suggest that recom- binational isolation may be best established for loci residing within a short distance from inversion breakpoints.
V
ARIANT forms of the proximal portion of chro-mosome 17 known as t haplotypes are polymor-
phic in natural populations of house mice (SILVER
1985; KLEIN 1986). These chromosomal segments
carry mutations that result in the violation of Mendel’s first law. I n a process known as transmission ratio
distortion (TRD), males heterozygous for a t haplo-
type and a wild-type chromosome can transmit the t
haplotype to over 95% of their offspring, whereas
transmission in females is n o r m a l . T R D is a special
case of meiotic drive because both sperm types are
produced in Mendelian proportions, but those carry- ing the t haplotype have a relative advantage at fertil- ization (OLDS-CLARKE 1986).
t haplotypes share a number of properties with
o t h e r systems of meiotic drive, such as SD a n d SR in
Drosophila (CROW 1988; WU a n d HAMMER 1991),
and represent informative examples of coadapted
gene complexes (DOBZHANSKY 1 9 5 1). In particular,
every meiotic drive system that has been analyzed in
detail involves interactions among closely linked
genes, and chromosomal rearrangements that sup-
’
Present address: Division of Biotechnology, Biosciences West, University of Arizona, Tucson, Arizona 85721.Genetics 128: 799-8 12 (August, 199 1)
press recombination between these loci. In fact, the- oretical studies predict that recombination suppres-
sors such as inversions will be favored by selection
(THOMSON a n d FELDMAN 1974; CHARLESWORTH a n d
HARTL 1978). In the case of t haplotypes, five loci
that are required for the expression of T R D have
been mapped across the 20-cM t region of chromo-
some I7 (LYON 1984; SILVER and REMIS 198’7). Re-
combination between t haplotype and wild-type alleles
of these loci is suppressed by a series of four non- overlapping paracentric inversions (ARTZT, SHIN and
BENNETT 1982; HERRMANN et al. 1986; SARVETNICK
et al. 1986; HAMMER, SCHIMENTI a n d SILVER 1989).
These inversions have probably been selected for
because only t haplotypes with a complete set of T R D alleles are transmitted at high ratios, and only high
ratio t haplotypes survive for significant periods of
time in natural populations (LEWONTIN and DUNN 1960).
TRD and recombination suppression should lead to
the rapid fixation of t haplotypes in mouse popula-
tions. However, they have been found only at fre-
quencies between 5 and 15% in most populations of
800 M. F. Hammer, S. Bliss and L. M. Silver
ern European house mouse M u s musculus, and the Far
Eastern species M u s castaneus a n d Mus molossinus
(KLEIN, SIPOS, a n d FICUEROA 1984; KANEDA et al.
1989). Two factors appear to play a role in the pre-
vention of fixation. First, most t haplotypes carry
recessive lethal mutations, with at least 16 different
complementation groups identified in wild mice
(KLEIN, SIPOS, a n d FIGUEROA 1984; KLEIN 1986).
Second, all t haplotypes cause male sterility in animals
with two complementing lethal forms or homozygous for a nonlethal t haplotype.
There is evidence that the origin of t haplotypes predates the divergence of t h e species in which they are found, and that they have existed for at least 2-4
million years (DELARBRE et al. 1988; HAMMER, SCHI-
MENTI and SILVER 1989). Along with the preservation
of allelic combinations at T R D loci, the presence of
the inversions also preserves combinations of variant
alleles of other loci within the region of recombination
suppression. T h e long-term recombinational isolation
has led to DNA sequence divergence and the estab- lishment of alleles that distinguish wild-type chromo-
somes and t haplotypes. For example, DNA sequences
from the noncoding regions of t complex protein-1
( T c p - I ) , DNA segment Lehrach 122 (DI7Leh122),
and a-globin pseudogene-4 (Hba-fd) demonstrate a
wild-typelt haplotype divergence of 1-2% (FRISCHAUF
1985; WILLISON, DUDLEY a n d POTTER 1986; M. F.
HAMMER and L. M. SILVER, unpublished results). In
addition, a number of protein-coding and anonymous DNA loci have alleles that are associated exclusively with t haplotypes (SILVER et al. 1983, 1987; Fox et al.
1985). However, there are a number of exceptions to
the patterns just described. At certain loci, alleles have
been found that are shared between t haplotypes and
wild-type chromosomes (DEMBIC, SINGER a n d KLEIN
1984; SKOW et al. 1987; NADEAU a n d PHILLIPS 1987).
These exceptions present interesting evolutionary
problems. Some authors have interpreted the exist-
ence of mosaic chromosomes (those carrying a mix- ture of wild-type alleles a n d t-like alleles) as evidence
for the persistence of ancestral polymorphism that was
present in the population that gave rise to t haplotypes
(FIGUEROA et al. 1988; ERHART et al. 1989). Alterna-
tively, mosaic chromosomes may be the result of rare
recombination between contemporary t haplotypes
a n d wild-type chromosomes (ERHART et al. 1989).
This would imply that t haplotypes could be disassem- bled and would raise questions concerning the stability
of coadapted gene complexes in evolution.
H e r e we present an investigation of mosaicism a t t h e Hba-ps4 locus mapping to the most distal inversion o f t haplotypes. The data provide evidence for a recent genetic exchange between t haplotypes and wild-type
chromosomes as the cause of mosaicism at this locus.
MATERIALS A N D METHODS
Mice: Table 1 lists the geographic origins and sources of
76 animals representing 10 taxa from 29 localities. The survey includes 11 laboratory strains (see Table 1 legend), 1 1 mice trapped in the wild, and 3 1 laboratory descendants
of separate lines of wild-caught mice. In addition, 23 lines
of mice carrying independent t haplotypes were examined.
Genomic DNA purification and analysis: DNA was prepared from tissues that were kept at -20" until used.
High molecular weight DNA was isolated according to
standard procedures. Two micrograms of genomic DNA
was digested to completion with Tag1 restriction endonucle-
ase (New England Biolabs), the fragments were size fraction- ated by 0.8% agarose gel electrophoresis and transferred to nylon membranes (GeneScreen, New England Nuclear) and baked according to the manufacturer's specifications. The DNA was covalently bound to the nylon by exposure to 254 nm light.
DNA probes and hybridization: The following recom- binant plasmids were used as molecular probes to study selected chromosome I 7 loci. Three obtained by microdis-
section-Tu48, Tu1 19, Tu89-define the loci Dl 7Leh48,
Dl7Lehll9, DI7Leh89, respectively (ROHME et al. 1984). A
cosmid subclone (Bb-40) is used to define Dl 7Leh66b (SCHI-
MENTI et al. 1987). A genomic fragment 0.7 kilobase (kb) upstream to Tcp-I gene (probe 29X) was kindly provided by K. WILLISON (WILLISON, DUDLEY and POTTER 1986). A
0.92-kb BamHI fragment containing portions of exon 5 and
6 and the intervening intron of the Pim-1 gene was provided by A. BERNS (CUYPERS et al. 1984).
T h e a-globin pseudogene-4 (Hba-ps4) and the a-A crystallin
(Crya-I) genes were isolated from a homozygous genomic
library derived from cell line DNA and kindly provided by
JOHN SCHIMENTI. Lambda clones containing these sequences
were isolated, digested with EcoRI and BamHI for subclon-
ing into pBR322. Four Hba-ps4 region subclones of 1.6, 1.3, 1 and 0.6 kb are shown in Figure 3 and referred to as Hba-6, -7, -9 and -1 1, respectively. Hba-6 and Hba-9 span a
2.6-kb region within and immediately surrounding the Hba-
ps4 pseudogene. Hba-7 and Hba-1 1 define a region of DNA
that lies 3-6 kb downstream from the pseudogene. This
entire region will be considered as a single locus. A Crya-2
Barn HI-Bam HI subclone of 0.6 kb was used in this study and is referred to as Cry-1 0.
Radioactive probes were produced by polymerization
from a mixture of random oligonucleotide primers on tem-
plates of denatured DNA (FEINBERG and VOGELSTEIN
1983). Hybridization to nylon membranes was performed
according to the procedure of CHURCH and GILBERT (1 984).
DNA sequencing: The 2.6-kb insert of the BALB/cJ Hba- ps4-containing plasmid (LEDER et al. 1981) and the sub-
cloned fragments Hba-9 and -6 were annealed to gene
specific oligonucleotide primers spaced at 200-400-bp in- tervals and sequenced using the double stranded sequencing method of BARTLETT, GAILLARD, andJoKLIK (1 986). A 1.4- kb region of PCR-amplified D N A from the DEAl allele of Hba-ps4 was also sequenced using this method. Y . NISHIOKA
initially provided 1.3 kb of the unpublished BALB/cJ Hba-
ps4 sequence.
High resolution restriction analysis: Three micrograms of genomic DNA were digested to completion with 1 of 1 1
Genetic Exchange in the t Complex
TABLE 1
Sources of mice subjected to restriction enzyme analysis
80 1
Mouse number and kind Geographic origin Source"
M. domesticus 1 2 :3 4 5 6-8 9-1 0
1 1 12-13 14-16 17 18 19 20-2 1
22-23 24-29 :30-3 1 32
M . musculus
3 3
3 4
3 5
36 37 38 39 M. castaneus 40 41 42 43 44
M . molossinu.!
M. bactrianw M. abbotta 45 46 BALB/cJ CSH/HeJ 1 29/sv C57BllO/J
S M/J DME DMA I)I H I CJ
D1J D I T WMP/PAS DPL DEA DEA DES DEG DEW PWK/PAS MCB MCS MYB MA1 MPW MDS CASA/RK C T T
MOLC/RK BI0.BACI BlO.BAC2 XBS AYG Laboratory Laboratory Laboratory Laboratory Laboratory Morocco, Erfoud Morocco, Azrou Israel, Haifa Israel, Jerusalem Israel, Jerusalem Italy, Tirano Tunisia, Monastir Peru, Lima Egypt, Abu Rawash Egypt, Abu Rawash Egypt, Saqqara
Egypt, Giza, near Bashtil Egypt, Wadi El Naturn
C7echoslavakia, Prague Czechoslavakia,
Bratislava Czechoslavakia,
Studenec. Yugoslavia, Belgrade Austria, Illmitz Poland, Warsaw Denmark, Skive
Thailand, Thon Buri Thailand, Thon Buri
Japan, Kyushu
Pakistan, Lahore Pakistan, Lahore
Bulgaria, Slantchev Briag Yugoslavia, Gradsko
1 1 1 1 1 2 2, 3 4 4 2, 4 2 5 3 2 7 7 7 7 5 2 2 4 2 2 2 1 3 1 12 12 4 3
-
" Inbred laboratory strains were provided by T h e Jackson Laboratory, Bar Harbor Maine (1) and J. L. GUENET ( 5 ) ; laboratory raised
descendents of wild-caught individuals were provided by MICHAEL POTTER (2), R. D. SAGE ( 3 ) , V . CHAPMAN (4). U . RITTE ( 6 ) ; and wild- caught mice were provided by I,. D'HOOSTELAERE (7). t haplotypes were from Princeton (8) o r provided by J. KLEIN (9). H. WINKING ( I O ) , J. FOREIJT (1 1). T h e congenic inbred strains from M . bactrianus were provided by K. FISCHER-LINDAHL (1 2). All t haplotypes were maintained a s congenic strains except tTU"" a n d t"' which were t l t . For further details on the original collection of mice see FERRIS et al. (1983) and
POTTER, NADEAU and CANCRO (1986).
M . hortulanus
47 HAH
48 HYP
49 SSC
5 0 SSG
51 SMA
52 C v T
5 ?l COT
54 t "
55 56 57
58 t ( I
59 t" 2 60 t" 5 61 y l ?
6 2 t"'3? M. spretus
M . cervicolor
M. caroli
t haplotypes t l . h l
lu6 J
I lrr<,X
6 3
64
t ~ r l , , c ' 7 ( ~
I !,X 27
65
66 to4
67 68 69 70
71 tl>,h4 72
73
74 tl""U
75 t'l""'l I 76
l-aW2Y
ru,m
t"7'
l - u l l z h
I,," 20
l u h 5 t l t < h h
I i w 2 4
Austria, Halbturn Yugoslavia, Pancevo
Spain, Cadiz Spain, Granada Morocco, Azrou
Thailand, Chon Buri
Thailand, Loei
Laboratory Italy, Alpic Drobic Italy, Ardenno Egypt, Giza France, Paris USA, Philadelphia USA, New York USA, California USA, Montana USA, Michigan Federal Republic of Ger- many, Dudelhof Federal Republic of Ger- many, Erpenhausen Czechoslavakia, Lhotka Czechoslavakia, Brno
Denmark, S. Jutland
Poland, Bialowieza USSR, Ryazan,
Astrachan Italy, Cremona Italy, Caneto, Pavese Italy, Luino Italy, Calcinato Chile, Buin
Federal Republic of Ger- many, Langenargen
2 3 3 4 2 2 2 8
I 0 10 9 8 8 8 8 8 9 9 9 11 9 8 9 9 10 10 10 10 9 9
analyzed by polyacrylamide gel electrophoresis (KREITMAN
and ACUADE 1986). Each fragment pattern produced by a given enz me was assigned a capital letter, with A being reserved
?
or the pattern appearing in the known BALB/cJ Hba-ps4 sequence. Cleavage maps for each enz me were constructed by the sequence comparison m e t h o J i n which one maps the sites needed to account for each fragment pattern by comparison with the known sequence (FERRIS et al. 1983).RESULTS
Variation at t complex loci: DNA probes for eight loci mapping to the t complex were used in a survey
of TaqI restriction sites in 23 previously described,
independently obtained t haplotypes, 9 inbred labo- ratory strains (listed in the legend to Table l ) ,
2
congenic lines containing a segment of chromosome17 derived from Mus bactrianus, 3 2 wild-derived and wild-caught M. domesticus and M. musculus, as well as individuals representing
7
other species in the genus Mus (Table 1). T h e positions occupied by these loci within the t complex are shown in Figure 1. The results of the survey are shown in the APPENDIX (Table802 M. F. Hammer, S. Bliss and L. M. Silver
Wild-type
t haplotype
T R D b c i : D l D4 R D3 D2
CM: 2 4 6 8 10 12 14 16 18
FIGURE 1 .-Positions occupied by eight loci and four inversions within the mouse t complex. The centromere is depicted by the solid circle and the four chromosomal inversions by the shaded boxes (relative orientation on wild-type and t haplotypes indicated by slanted lines). The eight loci examined in this survey and their positions within the t complex are shown. The symbols 48, 119,66, Hba, Pim, Cry and 89 represent D l 7 L e h 4 8 , D I 7 L e h l l 9 , D 1 7 L e h 6 6 ,
Hba-ps4, Pim-1, Crya-1 and Dl 7Leh89, respectively. Also shown are
the five loci involved in transmission ratio distortion and the scale in centiMorgans.
characterized by at least one restriction fragment var- iant between wild-type and t haplotypes (APPENDIX, Table 4), and these variants define separate restriction fragment length polymorphism (RFLP) alleles. Alleles found in previously characterized t haplotypes and absent from all wild-type chromosomes are called t- specific.
Five patterns of variation are detected among the 8 loci: (1) at D17Leh48, D17Leh119, D17Leh89, and
Hba-ps4 (detected with probes Hba-7 and Hba-1 l ) , a
single t-specific allele is shared by all t haplotypes; (2) at the D17Leh66 locus (detected with the Bb40 probe), two t-specific alleles are carried by different t haplo- types; (3) at Pim-1, a single t-specific allele is present in a subset o f t haplotypes, and all others have an allele shared by some wild-type chromosomes; (4) at Tcp-1
(and at Hba-ps4 as detected with probes Hba-6 and Hba-9), an allele shared by all t haplotypes is present on some wild-type chromosomes; and (5) at Cry-1, three alleles present in t haplotypes are all found in wild-type chromosomes.
Six DNA probes, Tu48, Tu1 19, Tu89, Hba-7, Hba- 11 and Bb-40, identify RFLP alleles that distinguish all t haplotypes from all wild-type chromosomes of the house mice examined in this survey. Only these probes are “informative” in detecting t-specific alleles and determining whether chromosomes collected from wild-mice are complete (with all four inversions) or partial t haplotypes (missing one or more inversions).
The probes used to detect Tcp-I and Crya-I alleles failed to identify t-specific variants (Figure 2). All t
haplotypes examined carry the Tcp-I allele producing pattern 1 which was found in 80% of M . domesticus mice. T h e Tcp-I allele producing pattern 2 was iden- tified in 20% of M . domesticus and 100% of M . mus- culus mice. This contrasts with the results of two- dimensional protein electrophoresis (SILVER et al.
1983) and restriction fragment patterns analyzed with Tcp-1 cDNA (WILLISON, DUDLEY and POTTER 1986) where all t haplotypes were found to be associated
40 119 Tcpl 66 Hbae Hba7 Plm Cry 89
I 1 1
*’
1 I 1 2 4 5 7 II 1 I 2 1234 1 2 3 I 2 2 4 7 I 2
1 1 1 1 1 1 2 4 1
1 1 2 2 1 2 2 4 1
1 1 2 5 3 2 1 4 5 3 4 5 %
1 h l p l o t w 3
M. d o m l i c u s
D U l
x
S M l J
M. musculus
M. castamus
-
’’
M. m/ossiflur
1 1 1 6 7 2 2 1 3 7
1 1 2 . 5 3 2 1 5 4
Ourgroups
2 I 2 2 3 4 8910 3 2 1 3 1258 9101112
FIGURE 2.--Summary of the Tag1 restriction patterns for eight t-complex loci. Abbreviations are the same as used in Figure 1 . Each box represents the pattern obtained with a separate probe hybrid- izing in the vicinity of a particular locus. The type of filling repre- sents restriction fragment patterns which are assigned numbers
(APPENDIX, Table 3). The results for t w o non-overlapping portions of the Hba-ps4 region are shown separately. Solid boxes indicate a restriction pattern uniquely associated with t haplotypes whereas other types of filled boxes represent restriction patterns associated with wild-type chromosomes.
with one variant, and all wild-type chromosomes were associated with another. t haplotypes carry three al- leles of Crya-I producing fragments 4 (46%),
7
(42%) and 5 (12%).
The majority of M . domesticus animals have the allele producing fragment 4 (94%), whereas 6% have the allele producing fragment 7. For M .musculus, equal numbers of mice were found with alleles producing fragments 4 and 5.
T h e conclusions of SKOW et al. (1987) regarding the Crya-1 locus differ in some regards from this report. Using a cDNA probe and several restriction enzymes, they found the majority of t haploptypes surveyed to be associated with an allele they consid- ered to be t-specific (Crya-1‘). We have reexamined three t haplotypes with the “Crya-1‘” allele (t“’, tm3’,
and t”’7’) and have found that all three carry the allele
producing fragment 7 which is also present in 6% of wild-type M . domesticus chromosomes. Thus the Crya- I‘ allele can no longer be considered t-specific. T h e Crya-I” allele corresponds to the class of chromosomes that carry the allele producing fragment 4 in our survey.
Genetic Exchange in the t Complex 803
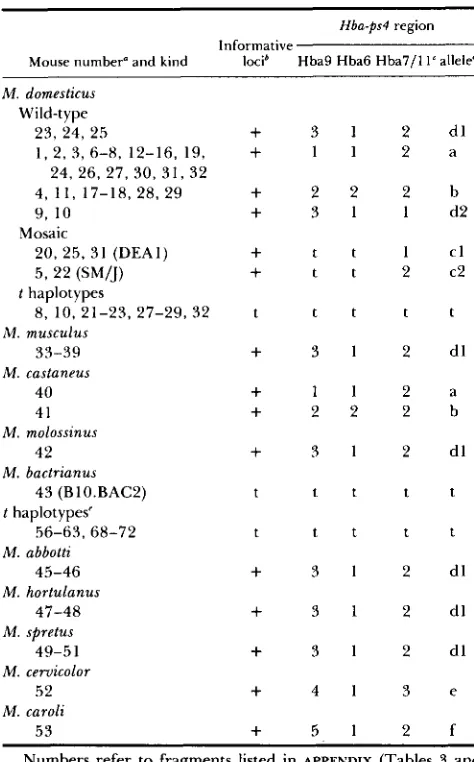
TABLE 2
Tag1 alleles and restriction patterns for the Hba-ps4 region and
four informative loci
Hba-ps4 region
Informative
Mouse number” and kind loci* Hba9 Hba6 Hba7/1 1‘ alleled
M . domesticus
Wild-type
2 5 , 24, 25
1 , 2, 3, 6-8, 12-16, 19, 24, 26, 27, 30, 3 1 , 32 4 , 1 1 , 17-18, 28, 29 9, 10
20, 25, 31 (DEA1) 5 , 22 (SWJ) Mosaic
t haplotypes
8, 10,21-23, 27-29,32
33-39 M . musculus
M . castaneus
40 41
M . molossinus
42
43 (BlO.BAC2) M . bactrianus
t haplotypes‘
M . abbotti
56-63,68-72
45-46
47-48
M . spretus
49-51
M . cervicolor
52 M . hortulanus
M . caroli
53
+
3 1 2 d l+
1 1 2 a+
2 2 2 b+
3 1 1 d2+
t t 1 c l+
t t 2 c 2t t t t t
+
3 12 d l+
1 1 2 a+
2 2 2 b+
3 21 d lt t t t t
t t t t t
+
3 1 2 d l+
3 1 2 d l+
3 1 2 d l+
4 1 3 e+
5 1 2 f4).
Numbers refer to fragments listed in APPENDIX (Tables 3 and
a Individuals listed twice are heterozygous.
The loci D l 7Leh48, Dl 7Lehl19, D17Leh66 and D l 7Leh89 have alleles that distinguish all characterized t haplotypes from wild-type chromosomes. t represents t-specific alleles at all four loci and
+
represents wild-type alleles at all four loci (see APPENDIX, Table 3, and text).‘ Hba-7 and Hba-1 1 results are shown in a single column because they span adjacent regions and hybridize the same restriction frag- ments.
Alleles named on basis of RFLPs with probes Hba6, Hba9, Hba7/11 (see Figure 3).
Previously characterized t haplotypes.
al. (1988) speculated that they had recovered a t haplotype from this subspecies based on the presence of the 5.2-kb TaqI allele of Hba-ps4.
Mosaic chromosomes: Chromosomes associated with both t-specific and wild-type alleles have been referred to as mosaic (ERHART et al. 1989). A subset of these chromosomes may be equivalent to partial t
haplotypes, which are generated by rare crossing over between complete t haplotypes and wild-type chro- mosomes. However, it is difficult to explain the origin
of some mosaic chromosomes by simple recombina- tion events alone when wild-type and t-specific alleles are found to be intermingled. Mosaic chromosomes of the latter type were identified with the use of DNA probes hybridizing to two loci, Pim-1 and Hba-ps4
(Figure 2). Seven of 23 t haplotypes were found to carry a Pim-I restriction fragment pattern indistin- guishable from that associated with all wild-type M .
domesticus chromosomes. These results agree with the
findings of NADEAU and PHILLIPS (1 987).
Previous studies on Hba-ps4 identified a series of
TaqI alleles associated with wild-type chromosomes
and showed that a 5.2-kb TaqI fragment was shared by all t haplotypes (FOX, SILVER and MARTIN 1984; SILVER et al. 1987). Three distinct wild-type Hba-ps4
alleles have been identified with the use of the restric- tion enzyme TaqI (D’EUSTACHIO et al. 1984; FOX, SILVER and MARTIN 1984; DELARBRE et al. 1988);
Hba-ps4“ is associated with 3.8- and 1.4-kb TaqI frag-
ments, Hba-ps4‘ is associated with a 3.4-kb TaqI frag- ment, and a third allele is associated with 2.0- and 1.4-
kb Tag1 fragments (defined as H b ~ - p s 4 ~ ; Table
2).
In the present study, the 5.2-kb Hba-ps4‘ was found in association with all 23 t haplotypes examined. How- ever, this allele was also found in the inbred wild-type strain SM/J (mouse number 5) as well as four wild- derived mice that carried no other t-specific alleles (mice numbers 20, 22, 25 and 31: DEA1, DEA3, DES2 and DEG2; see Table 2 and Figure
2).
By the definition of ERHART et al. (1989), all five of these mice carry mosaic chromosomes. These results may be interpreted in two ways. First, it is possible that a single point mutation(s) occurred recently in one or more wild-type chromosomes to give rise to a 5.2-kbTaqI fragment independent o f t haplotypes. This iden-
tity by “coincidence” can easily be accounted for by any mutation which eliminates the TaqI site located between the 3.8- and 1.4-kb fragments characteristic of the wild-type Hba-ps4” allele (Figure 3). Alterna- tively, the DNA sequences surrounding this site could be identical by descent and indicative of the presence of t-like regions within otherwise wild-type chromo- somes. It is possible to distinguish between these two interpretations through an analysis of additional re- striction site polymorphisms in the vicinity of the TaqI
sites responsible for the 5.2-kb fragment. In a previous study, we used such an analysis to demonstrate a single point mutation as the likely explanation for the pres- ence of the 5.2 kb Tag1 fragment in loose linkage with the partial t haplotype tTUw3* (SILVER et al. 1987).
Tag1 site variation in the vicinity of Hba-ps4: The
804 M. F. Hammer, S. Bliss and L. M. Silver
2 1 I 1 1 1 . I
,
a3 1 I c I 1 . I I b
4 1 1 I
I .
. I I d2Mosaic
5 1
. .
1 . I , 1 c16 1
.
1 1 .I 1 c2I haplotype
7 1
.
I , 1 1 I 11 1 1 1 1 1 1 1 1 I I 1 1
0 2 4 6 8 10 12
kilnbases
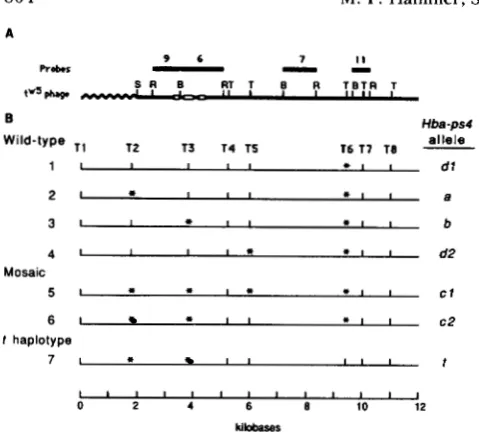
FIGURE 3.-DNA probes and restriction sites in the vicinity of
the Hbu-ps4 locus. (A) Restriction map of the tu' genomic clone. All
Eco RI (R), BamHI (B) and TuqI (T) sites within a 12-kb region
surrounding the Hbu-ps4 locus are shown. The small open boxes
represent the three open reading frames of Hbu-ps4. The solid
black bars, numbered 9, 6, 7 and 11, represent four genomic
subclones-Hba9, Hba6, Hba7 and Hba 1 1-that were used as probes.
(B) TaqI map of the Hbu-ps4 region derived from Southern analysis
of genomic DNA. TuqI sites are numbered T1-T8 and indicated
by vertical lines; an asterisk indicates the absence of a TuqI site.
The seven Tag1 haplotypes are referred to as alleles with the
designated letters and numbers (shown on right). The relationships between allele names shown here and specific restriction fragment sizes listed in APPENDIX (Table 4) are indicated in Table 2.
3). Haplotype #1 represents a subset of the H b q "
class and is found in all taxa including the more distantly related species Mus abbotti, Mus hortulanus and Mus spretus. This haplotype is considered to be ancestral and will be referred to as Hba-ps#'. Haplo- type #2 (Hba-psC), #3 (Hba-ps4') and #4 (named Hba-
p ~ 4 ~ ' here) could all be derived from H b ~ - p s 4 ~ ' by a
single site loss ( T 2 , T 3 and T 5 , respectively; Figure 3). Haplotype
#7
(Hba-ps4') was found uniquely in association with all previously characterized t haplo- types, and differs from all other haplotypes by a site gain at T6. The mosaic chromosomes, defined above on the basis of the 5.2-kb "t-fragment," are divided into two groups-haplotypes #5 (Hba-ps4"') and #6 (Hba-ps4")-based on polymorphisms in TaqI sites de- tected by probes Hba7 and Hbal l.High resolution restriction maps: Eleven restric- tion enzymes were used to obtain high resolution maps (HRM) of a 2.6-kb EcoRI fragment of DNA (spanning probes Hba-9 and Hba-6) within and immediately surrounding the Hba-ps4 locus from 14 mice (APPEN- DIX, Table 5). A total of 8 different cleavage patterns were identified and analyzed in conjunction with the nucleotide sequence for BALB/cJ. Restriction maps were inferred for over 100 sites within each genomic DNA sample (data not shown). A comparison of Hba- ps4 restriction maps obtained with a wild-type BALB/
CJ chromosome (Hba-ps4"), a complete tw5 haplotype
(Hba-ps4')), and the five mosaic chromosomes (Hba-
Ps4" and Hba-ps4") is shown in Figure 4A. There are
a total of 15 variable sites (1 1 restriction enzyme differences and 4 length polymorphisms) among the maps obtained. Fourteen of these sites differ between t"' and BALB/cJ. There are two classes of HRM for
Hba-ps4 alleles on mosaic chromosomes that are con-
sistent with the allelic designation on the basis of TaqI
sites: the two mosaic chromosomes with an Hbu-ps4"
allele (SM/J and DEA3) share the same HRM, as do the three chromosomes with the Hba-psC' allele (DEA1, DES2 and DEG2). These two HRMs are distinguished from one another at 4 sites. In addition, these maps are distinguishable from that of Hba-ps4'
by
7
sites (Hba-ps4") and 5 sites (Hba-ps4").Further examination of the HRMs reveals the ex- istence of nested sets of shared sites among the maps
of Hba-ps4", H ~ u - P s ~ ' ~ and tw5 in the 3' region (indi-
cated by the open lines in Figure 4A), and a similar sharing of sites occurs among the maps of Hba-ps4",
H ~ U - P S ~ ' ~ and BALB/cJ in the 5' region (indicated by
shaded lines in Figure 4A).
M. domesticus chromosomes previously classified with the most common wild-type allele ( b ) can be subdivided into three closely related groups based on HRMs (Figure 4B). All three maps share a 3' cluster of restriction sites with each other, with the maps of the mosaic chromosomes, and with that of tW5. In fact, the HRM of the wild-type inbred strain B10 is indis- tinguishable from the HRM of the mosaic chromo- some SM/J. T h e sharing of clusters of sites is sugges- tive of one or more recent genetic exchange events between progenitors to the various chromosomes ana- lyzed here.
DNA sequence comparisons between wild-type, t haplotype and mosaic chromosomes: T o further in- vestigate the possibility of genetic exchange events, DNA sequences were determined for the Hba-ps4
locus present in BALB/cJ, tw5, and the mosaic chro- mosome DEAl (data not shown). T h e wild-type (BALB/cJ) and t haplotype sequences differ at 18 positions in a 1072 bp region (Figure 5A). When the DEAl Hba-ps4 sequence is compared to the others, a boundary is observed between nucleotides 5 10 and 675. In the region 5' to this boundary, DEAl is identical to BALB/cJ at 5 of the 6 sites that distinguish BALB/cJ from tw5; in the region 3' to this boundary, DEAl is identical to tW' at 11 of the 12 sites that distinguish wild-type and t chromosomes (see boxed- in regions in Figure 5A). This boundary contains the putative 5' breakpoint of a genetic exchange between wild-type sequences and t haplotypes.
Genetic Exchange in the t Complex 805
~ I I I I I I ~ I I I I I I I I I I ”
1 2 3 4 5 6 7 8 9 1011 12 13 14 15 FIGURE 4.-High resolution re-
A striction maps of the Hba-ps4 locus
n for BALB/cJ, and mosaic chro-
mosomes. T h e locations of polymor- phic restriction sites detected in ge-
nomic DNAs with eleven enzymes
are shown with a vertical line and a letter or number designating the en- zyme Hue111 (H), Sau3AI (3). Hphl
(P). Tag1 (T), S c r f f (S), Ddel (D), RsaI
(R). HinfI (F), Ah1 (A). Length vari- ation is indicated with inverted tri- angles. FAch variable site is given a number (1-15; site 8 is equivalent to T 3 in Figure 3B). Regions with ho-
mology to BALB/cJ are indicated
with shaded lines, and regions with homology to 1”” are indicated with open lines. (A) Restriction maps for
BALB/cJ. tu” and mosaic chromo-
somes with a 5.2-kb Tag1 fragment. T h e allelic designation is shown in parentheses. (B) Restriction maps for alleles with the 3.4-kb TaqI fragment
(Hba-ps4’). ( C ) Restriction maps for
two alleles of M . castaneus.
tended in the 3’ direction through analysis with the probes Hba7 and Hbal 1 (APPENDIX, Table 4A). Ho-
mology between DEAl and t”” continues through two informative sites to the 5-kb position in Figure 5B. At position 10 kb in Figure 5B, the situation reverses with identity between DEAl and BALB/cJ at all four informative sites. Therefore, the length of the region of homology between DEA 1 and tu’’ measures between 2 and 6 kb.
DISCUSSION
Wild-type M . domesticus chromosomes can carry a t-like form of Hba-ps4: Wild-type chromosomes recovered from the inbred strain SM/J and mice living in several Egyptian localities carry the 5.2-kb TaqI
allele of Hba-ps4 that is characteristic of all well- defined complete t haplotypes (Hba-ps4‘). All of the other loci examined on these Egyptian chromosomes have non-t haplotype alleles (Figure 2). Previous stud- ies demonstrated that the 5.2-kb Tag1 fragment could arise independently of t haplotypes (SILVER et al. 1987). T o determine the relative relationship of an Egyptian Hba-ps4 allele (from the DEAl strain; the allele is named Hba-ps4’) to wild-type and t-alleles at this locus, these three sequences were compared by high resolution restriction mapping and DNA se- quencing. T h e results demonstrate a strong homology between Hba-ps4“ and Hba-ps4‘ that is restricted to a region of DNA of 2-6 kb in length (Figure 5). Se- quences flanking both sides of this t-homologous re- gion are much more homologous to that of the stand- ard wild-type chromosome from the inbred strain
BALB/cJ. Two other Egyptian mice recovered from
nearby localities (DES2 and DEG2, Table 1) carry the same Hba-ps4” allele by the criteria of HRM. A num- ber of other wild-derived and inbred mice carry Hba- ps4 alleles that are very similar to Hba-ps4“ with homology to Hba-ps4‘ over a somewhat shorter (by 400-500 bp) length of DNA. T h e true mosaic nature of all of these otherwise wild-type chromosomes is demonstrated by the existence of short DNA segments with multiple informative sites showing strong homol- ogy to t haplotypes. These chromosomes-the only class of “wild-type” mosaic chromosome identified in our survey-represent 14% of the 22 chromosomes sampled directly from wild mice. This differs from the study of ERHART et al. (1989) in which several classes of mosaic chromosome were found and repre- sented 45% of all chromosomes recovered from wild mice.
Genetic exchange between t haplotypes and their wild-type homologs: There are two explanations for the presence of mosaic chromosomes in populations of wild mice. t-like alleles on wild-type chromosomes could represent persisting polymorphisms that existed in the common population that gave rise to t haplo- types and present-day wild-type chromosomes. Assum- ing that barriers to recombination did not preexist in this ancestral population, such alleles would be ex- pected to be shared by the progenitors to both wild- type chromosomes and t haplotypes. An alternative explanation is that recombination between complete t
haplotypes and their wild-type homologues has oc- curred relatively recently, well after the appearance of the inversion polymorphism.
M. F. Hammer, S. Bliss and L. M. Silver
806
A
BALB/cJ
DEAl
t w 5
B
C G G C T T A GA A T GGTAA
G$G G C C C G -2 AG T C A A C G
I I I I
A A C G T T C G -2 AG T CAACGC
5' exchange poinl 3' exchange point
C
I
BALB/cJ DEAl
L_.___._..._....__._ m U I
R A: R
R A T
1 w 5 ... : T A
DEAl
S M / J p
DES5
-
1 ~ 1 1 1 ~ 1 1 1 1 1
2 4 6 8 10
kilobases
FIGURE 5.-DNA sequence and restriction site comparisons of the Hba-ps4 region among BALB/cJ, tW5 and the mosaic chromo- some from DEAl. (A) Variable nucleotide positions are shown for 1072 bp (scale in base pairs shown at top) including the 5' flanking region and open reading frames 1 and 2 of Hba-ps4. T h e first box
shows a region where DEAl and BALB/cJ are identical at five out
of six nucleotide positions and the second box shows a region where
DEAl and tU5 are identical at ten out of eleven nucleotide positions. (B) Variable restriction sites in the 3' flanking region. T h e se- quenced region and the two boxes from (A) are shown under the bracketed line. T h e region of homology between DEAl and t W 5 is extended by the sharing of RsaI (R) and AluI (A) sites. A region of undetermined homology is indicated with dotted lines and includes a Taql site (T) that is absent in DEAl and shared between BALBc/ J and tW5. A region of shared restriction sites between DEAl and
BALB/cJ is indicated with a box on the far right of the diagram.
( C ) t-homologous regions carried by DEA 1, SM/J and DES5 chro-
mosomes. T h e line indicates wild-type regions and the solid box indicates homology with t haplotypes. T h e open portion of the box indicates regions of undetermined homology. T h e scale for (B) and (C) is shown at the bottom in kilobases.
predictions of the former model. First, the mosaic chromosomes do not carry a mixture of wild-type and t-specific alleles at several loci as might be expected if they were descended directly from the population giving rise to t haplotypes. Second, shared ancestral alleles would be expected to diverge such that base substitutions would occur randomly throughout the Hba-ps4 region. In contrast, the results demonstrate a clustering of shared enzyme sites between tw5 and the mosaic chromosomes. This clustering is highly signif- icant at the level of DNA sequence as shown in Figure 5A, and provides strong support for the model of a recent genetic exchange as an explanation for the mosaic chromosomes reported here. T h e low level of
sequence divergence between DEAl and within the region of shared sites (0.02%) compared to the
1.8% sequence divergence between tw5 and BALB/cJ gives further evidence that the genetic exchange event occurred recently relative to the event giving rise to the inversion.
Gene conversion: T h e position of Hba-ps4 within the distal inversion (Figure 1) makes the possibility of recovering the products of single crossover events in the vicinity of the pseudogene extremely unlikely. Single crossovers within paracentric inversions lead to inviable zygotes through the production of acentric and dicentric chromosomes. Exchanges of genetic ma- terial across inverted regions of t haplotypes are more likely due to double recombination or gene conver- sion.
Two lines of evidence suggest that the Hba-ps4" allele was created by gene conversion rather than by double crossing over. First, the length of the DNA exchanged between a t haplotype and the progenitor of the DEAl chromosome is between 2 and 6 kb. The probability of two crossover events occurring within this length of DNA is practically zero because strong positive interference will act over such short distances (KING et al. 1989). On the other hand, gene conver- sion events typically involve short tracts of DNA. T h e average length of conversion tracts at the rosy locus of D. melanogaster were shown to be 1208 f 790 bp (CURTIS et al. 1989). This is similar to measurements of the average meiotic conversion tract lengths in yeast which range from 0.7 kb to over 2 kb (BORTS and HABER 1989).
Second, there is evidence from experiments with D. melanogaster that the majority of recombinants from inversion heterozygotes are the products of gene con- versions rather than the classical double crossovers. Using a selective system to examine about 5 x IO6 zygotes, CHOVNICK (1 973) examined recombination between rosy mutant alleles in the center of a paracen- tric inversion heterozygote, and compared the results with experiments carried out in standard chromosome homozygotes. He found gene conversion frequencies were of the same order of magnitude in the inversion heterozygotes as in the standard arrangement homo- zygotes.
Origin of Hba-ps4 alleles: A single gene conversion between a t haplotype and a wild-type chromosome may explain the presence of mosaic chromosomes like DEAl in Egyptian mouse populations. All three mice bearing chromosomes with Hba-ps4" come from the geographical region close to Giza (and the pyramids). Therefore, it is very likely that these chromosomes are identical by descent. The other mosaic chromo- somes identified in this study ( i e . , Figure 4, restriction maps 2, 5-7) may have resulted from separate con- version events or secondary recombination events in-
Genetic Exchange in the t Conlplex
T 2 1 3 5 7T39 11 13 15 T5 T6 T7 T 2 1 3 5 7 T 3 9 11 13 15 T5 T6
A " A
807
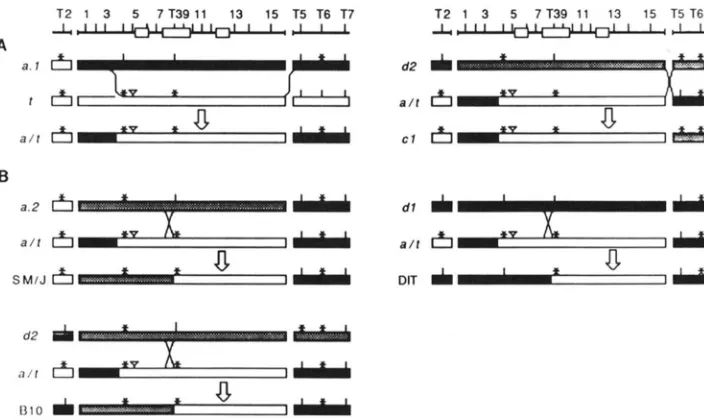
FIGURE 6.-Hypothetical scenario for the origin of mosaic haplotypes from known wild-type and f haplotype alleles. The positions of Tag1
(T2, T5-7) and other enzyme sites (1-15: site 8 is equivalent to T 3 ) are shown on the top of the diagram. Informative sites are indicated with vertical lines (presence) and asterisks (absence). (A) Two-step process in the formation of the DEA 1 Hba-ps4" allele. (Left) An ancestral
1 haplotype converts a wild-type chromosome with an allele similar to that carried by BALB/cJ (Hba-ps4""). creating an intermediate (Hba-
ps4"") with t-like sites 4-15, and BALB/cJ-like sites T5-T7. (Right) Recombination between sites 15 and T 5 w i t h Hba-ps4". (B) Formation
of Hba-ps4" (SM/J) and Hba-ps4'alIeles (haplotypes 5-7, Figure 48). The alleles carried by SM/J, BI 0 and DIT are formed by recombination
between sites 7 and 8 (T3) of Hba-ps4"" and three different wild-type alleles, Hba-psP', Hba-pdd2, and Hba-ps4". respectively.
the rosy locus between inverted regions of
D.
mela-nogaster was 1 O-4events per meiosis (CHOVNICK 1973),
it seems likely that Hba-psC'-related alleles are the result of intra-allelic recombination with a wild-type allele.
A hypothetical scenario for the origin of all mosaic haplotypes from known wild-type chromosomes and a hypothetical ancestral t haplotype is shown in Figure 6. This scenario, only one of many that are possible, is appealing because it involves the least number of total steps and emphasizes the greater probability of intra-allelic recombination over multiple inter-chro- mosomal conversion events. T h e origin of the Hba-
ps4" allele is envisioned as a two-step process where an ancestral t haplotype converts a wild-type chro- mosome similar to BALB/cJ (Hba-ps4".') thereby cre- ating an intermediate allele (which we call Hba-ps4"")
with t-sites 4 through 15, and BALB/cJ sites T 2 , 1-3 and T5-T7 (Figure 6A). This allele is now carried on a wild-type chromosome and can freely recombine with other wild-type alleles. Recombination with a wild-type H b ~ - p s 4 ~ ' allele, between sites 15 and T5, would create the arrangement of enzyme sites ob- served in Hba-ps4*'.
T h e origin of all other mosaic chromosomes can be explained by recombination between sites 7 and 8 of the Hba-ps4""interrnediate and one of three different closely related wild-type chromosomes (Figure 6B). This scenario implies that there is a recombination hotspot in which 3 intra-allelic recombination events have occurred within a 219-bp segment of the pseu- dogene (Figure 4, bases 91 1-1 130).
A separate conversion in
M.
castaneus? A similar region of t homology is also observed in one of two Hba-ps4 alleles of a wild-type chromosome from M .castaneus (Figure 4C). This may suggest that an inde-
pendent recombination event occurred between a M .
castaneus wild-type allele and one similar to Hba-ps4"'
'.
However, in the upstream region of both C T T and CASA alleles, there is a mixture of t-specific and wild- type sites. Two explanations can be offered. First, it is possible that these alleles are the product of a transfer of genetic information from a M . castaneus thaplotype (that differs from the t haplotypes examined from M . domesticus and M . musculus) to a M . castaneus wild-type chromosome. Recombination with wild-type alleles could then scatter t-specific sites onto different chromosomes.
A second explanation is that the alleles present in M . castaneus did not arise by recombination with a t haplotype, but are closely related to the alleles present in the ancestral population where t haplotypes origi- nated. For example, it is possible that the distal inver- sion that is now carried by all t haplotypes, originated in the range of M . castaneus. T h e recombination suppression that resulted from the fixation of the inversion chromosome would preserve the similarity of the t form of the Hba-ps4 allele with those present in M . castaneus.
808 M . F. Hammer, S. Bliss and L. M . Silver
populations of M . domesticus, one of which (Hba-ps4') is the most common allele among inbred laboratory strains (D'EUSTACHIO et al. 1984; FOX, SILVER and MARTIN 1984; DELARBRE et al. 1988). If this process has been recurrent, the same pattern of variation should occur for other loci. In fact, evidence for such patterns has been observed in surveys of other t com- plex loci. Nearly one third of the complete t haplo- types examined in this survey carry an allele of Pim-1 with TaqI sites identical to those of an allele carried by all M . domesticus chromosomes examined; a similar result was obtained by NADEAU and PHILLIPS (1 987). Other examples of putative genetic exchange between wild-type and t haplotypes have been reported at the Crya-1 locus (SKOW et al. 1987) and the H-2Ea locus (DEMBIC, SINGER and KLEIN 1984). All of these loci reside within the largest of the four inversion poly- morphisms-In( 17)2-that distinguish t haplotypes from wild-type chromosomes. It is possible that this inverted region is more likely to undergo the pairing necessary for exchange events to occur.
Conversion at t distorter loci would create mosaic t haplotypes with reduced transmission ratios and would be selected against. This means that Pim-1 and Crya-1 cannot be involved in TRD. Interestingly, there is the possibility that all TRD loci reside near the eight inversion breakpoints (Committee for Chro- mosome 17 et al. 199 l ) , whose adjacent regions might be less likely to undergo exchange events (CHOVNICK 1973), although this appears not to be the case for regions adjacent to insertion breakpoints in the Dro- sophila rosy locus (CLARK, HILLIKER and CHOVNICK
1988).
We thank Y. NISHIOKA for providing the unpublished BALB/cJ
Hba-ps4 sequence. This research was supported by a grant from the
National Institutes of Health to L. M. S. (HD20275) and by a
postdoctoral fellowship from the National Institutes of Health to M. F. H.
L I T E R A T U R E C I T E D
ARTZT, K., H-S. SHIN and D. BENNETT, 1982 Gene mapping
within the T/t-complex of the mouse. 11. Anomalous position of the H - 2 complex in t- haplotypes. Cell 28: 471-476.
BARTLETT, J. A,, R. K. GAILLARD and W. K. JOKLIK,
1986 Sequencing of supercoiled plasmid DNA. Biotechniques
BORTS, R. H., and J. E. HABER, 1989 Length and distribution of
meiotic gene conversion tracts and crossovers in Saccharomyces
cereuisiae. Genetics 123: 69-80.
CHARLESWORTH, B., and D. HARTL, 1978 Population dynamics
of the segregation distorter polymorphism of Drosophila mel-
anogaster. Genetics 89: 17 1- 192.
CHOVNICK, A. , 1973 Gene conversion and transfer of genetic
information within the inverted region of inversion heterozy- gotes. Genetics 75: 123-1 3 1.
CHURCH, G. M., and W. GILBERT, 1984 Genomic sequencing.
Proc. Natl. Acad. Sci. USA 81: 1991-1995.
CLARK, S . H., A. J. HILLIKER and A. CHOVNICK,
1988 Recombination can initiate and terminate at a large
4: 209-2 10.
number of sites within the rosy locus of Drosophila melanogaster. Genetics 118: 261-266.
Committee for Chromosome 17, K. ARTZT, D. BARLOW, W. F.
DOVE, K. FISCHER-LINDAHL, J. KLEIN, M. LYON and L. M.
SILVER, 1991 Maps of mouse chromosome 17: first report.
Mamm. Genome 1: 5-29.
CROW, J. F. , 1988 The ultraselfish gene. Genetics 118: 389-391.
CURTIS, D., S. H. CLARK, A. CHOVNICK and W. BENDER,
1989 Molecular analysis of recombination events in Drosoph- ila. Genetics 122: 653-66 1 .
CUYPERS, H. T., G. SELTEN, W . QUINT, M. ZIJLSTRA, E. R. MAAN-
DAG, w . BOELENS, P. VAN WEZENBECK, C. MELIEF and A.
BERNS, 1984 Murine leukemia virus-induced T-cell lym-
phomagenesis: integration of proviruses in a distinct chromo- somal region. Cell 37: 14 1 - 150.
D'EUSTACHIO, P., B. FEIN, J. MICHAELSON and B. TAYLOR,,
1984 The alpha-globin pseudogene on mouse chromosome
17 is closely linked to H - 2 . J. Exp. Med. 1 5 9 958-963.
DELARBRE, C., Y. KASHI, P. BOURSOT, J. S. BECKMAN, P. KOURLISKY,
F. BONHOMME and G . GACHELIN, 1988 Phylogenetic distri-
bution in the genusMus of t-complex-specific DNA and protein
markers: inferences on the origin of t-haplotypes. Mol. Biol. Evol. 5: 120-133.
DEMBIC, Z., A. SINGER and J. KLEIN, 1984 Eo : a history of a
mutation. EMBO J. 3: 1647-1654.
DOBZHANSKY, T. , 195 1 Genetics and the Origin ofSpecies. Columbia University, New York.
ERHART, M. A,, S. J. PHILLIPS, F. BONHOMME, P. BOURSOT, E. M.
WAKELAND and J. H . NADEAU, 1989 Haplotypes that are
mosaic for wild-type and t complex-specific alleles in wild mice.
Genetics 123: 405-415.
FEINBERG, A. P., and B. VOGELSTEIN, 1983 A technique for
radiolabeling DNA restriction endonuclease fragments to high
specific activity. Anal. Biochem. 132: 6-1 3.
FERRIS, S. D., R. D. SAGE, E. M. PRAGER, U. RITTE and A. C.
WILSON, 1983 Mitochondrial DNA evolution in mice. Ge-
netics 105: 681-721.
FIGUEROA, F., E. NEUFIELD, U. RITTE and J. KLEIN, 1988 t-
Specific DNA polymorphisms among wild mice from Israel and
Spain. Genetics 119: 157-160.
FOX, H., L. M. SILVER and G. R. MARTIN, 1984 An a-globin
pseudogene is located within the mouse t complex. Immuno-
genetics 19: 125-130.
Fox, H. S., G . R. MARTIN, M. F. LYON, B. HERRMANN, A,".
FRISCHAUF, H . LEHRACH and L. M. SILVER, 1985 Molecular
probes define different regions of the mouse t complex. Cell
4 0 63-69.
FRISCHAUF, A,"., 1985 T h e T / t complex of the mouse. Trends
HAMMER, M. F., J. SCHIMENTI and L. M. SILVER, 1989 The
evolution of mouse chromosome 17 and the origin o f t complex inversions. Proc. Natl. Acad. Sci. USA 86: 3261-3265.
HERRMANN, B., M. BUCAN, P. E. MAINS, A.-M. FRISCHAUF, L. M.
SILVER and H. LEHRACH, 1986 Genetic analysis of the prox-
imal portion of the mouse t complex: evidence for a second inversion within t haplotypes. Cell 44: 469-476.
KANEDA, H., Y . Y. MAEDA, K. MORIWAKI, M. SAKAIZUMI, C. TAYA,
S . WATENABE and H. YONEKAWA, 1989 t chromosome found in East Asiatic wild mice, M. m. molossinus , M . m. castaneus.
Mouse News Lett. 84: 119.
KING, T . R., W. F. DOVE, B. HERRMANN, A. R. MOSER and A.
SHEDLOVSKY, 1989 Mapping to molecular resolution in the
T to H-2 region of the mouse genome with a nested set of
meiotic recombinants. Proc. Natl. Acad. Sci. USA 86: 222-
226.
KLEIN, J. , 1986 Natural History of the Major Histocompatibility Complex. John Wiley & Sons, New York.
KLEIN, J., P. SIPOS and F. FIGUEROA, 1984 Polymorphism of t-
Genetic t h e E x c h a n g e i n t C o m p l e x 809
complex genes in European wild mice. Genet. Res. 3 0 93-
106.
KREITMAN, M., and M. AGUADE, 1986 Genetic uniformity in two
populations of Drosofhila melanogaster as revealed by filter hybridization of four-nucleotide-recognizing restriction en-
zyme digests. Proc. Natl. Acad. Sci. USA 83: 3562-3566.
LEDER, A,, D. SWAN, F. RUDDLE, D'EUSTACHIO and P. LEDER, 1981 Dispersion of alpha-like genes of the mouse to three
different chromosomes. Nature 293: 196-200.
LEWONTIN, R. C., and L. C. DUNN, 1960 T h e evolutionary dy-
namics of a polymorphism in the house mouse. Genetics 45:
LYON, M. F. , 1984 Transmission ratio distortion in mouse t- haplotypes is due to multiple distorter genes acting on a re-
sponder locus. Cell 37: 621-628.
NADEAU, J. H., and S. J. PHILLIPS, 1987 T h e putative oncogene
Pim-I in the mouse: its linkage and variation among t haplo-
types. Genetics 117: 533- 541.
OLDS-CLARKE, P., 1986 Motility characteristics of sperm from the
uterus and oviducts of female mice after mating to congenic males differing in sperm transport and fertility. Biol. Reprod.
POTTER, M.,J. H. NADEAuand M. P. CANCRo(Editors), 1986 T h e
wild mouse in immunology. Curr. Top. Microbiol. Immunol.
ROHME, D., H. Fox, B. HERRMANN, A,". FRISCHAUF, J.-E. ED-
STROM, P. MAINS, L. M. SILVER and H. LEHRACH,
1984 Molecular clones of the mouse t complex derived from
microdissected metaphase chromosomes. Cell 36: 783-788.
SARVETNICK, N., H. Fox, E. MANN, P. MAINS, R. ELLIOT and L. M.
SILVER, 1986 Nonhomologous pairing in mice heterozygous
for a t haplotype can produce recombinant chromosomes with
duplications and deletions. Genetics 113: 723-734.
SCHIMENTI, J., L. VOLD, D. SOCOLOW and L. M. SILVER, 1987 An
unstable family of large DNA elements in the center of the mouse t complex. J. Mol. Biol. 194: 583-594.
SILVER, L. M. , 1985 Mouse t haplotypes. Annu. Rev. Genet. 19:
705-722.
34: 453-467.
127: 373-395.
179-208.
SILVER, L. M., and D. REMIS, 1987 Five of the nine genetically defined regions of mouse t haplotypes are involved in trans- mission ratio distortion. Genet. Res. 49: 51-56.
SILVER, L. M., J. UMAN, J. DANSKA and J. 1. CARRELS, 1983 A
diversified set of testicular cell proteins specified by genes within the mouse t complex. Cell 35: 35-45.
SILVER, L. M., M . HAMMER, H. Fox, J. CARRELS, M. BUCAN, B.
HERRMANN, A.- M. FRISCHAUF, H. LEHRACH, H. WINKING, F.
FIGUEROA and J. KLEIN, 1987 Molecular evidence for the
rapid propagation of mouse t haplotypes from a single, recent,
ancestral chromosome. Mol. Biol. Evol. 4: 473-482.
SKOW, L. C., J. N. NADEAU, J. C. AHN, H.3. SHIN, K. ARTZT and
D. BENNETT, 1987 Polymorphism and linkage of the alpha-A
crystallin gene in t haplotypes of the mouse. Genetics 116: 107- 1 1 1 .
THOMSON, G. J., and M. W. FELDMAN, 1974 Population genetics
of modifiers of meiotic drive 11. Linkage modification in the
segregation distortion system. Theor. Popul. Biol. 5: 155-162.
WILLISON, K. R., K. DUDLEY and J. POTTER, 1986 Molecular
cloning and sequence analysis of a haploid expressed gene
encoding t complex polypeptide 1 . Cell 44: 727-738.
WU, C.-I., and M. F. HAMMER, 1991 Molecular evolution of
ultraselfish genes of meiotic drive systems, pp. 177-203 in
Evolution at the Molecular Level, edited by R. K. SELANDER, A.
G. CLARK and T. S. WHITTAM. Sinaur Associates, Sunderland,
Mass.
Communicating editor: A. CHOVNICK
APPENDIX
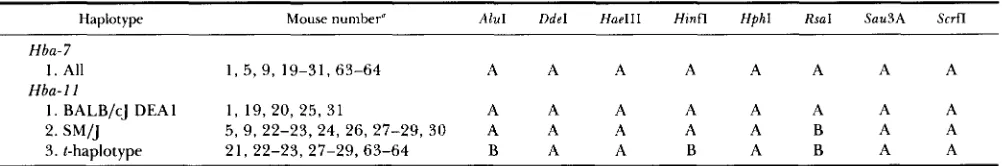
TuqI restriction fragment patterns for 8 loci and
fragment sizes for 11 t complex probes are given in Tables 3 and 4, respectively. Fragment patterns for
810 M . F. Hammer, S. Bliss and L. M. Silver
TABLE 3
TaqI restriction fragment patterns for 8 loci in the t complex
Tcp- 1 66 Hba
Mouse number and kind 4 8 119 29X Bb40 9 6 7 11 la 1 0 89
Pim Crya
M . domesticus 1 BALB/cJ 2 C3H/HeJ 3 129/sv 4 C57B11 O/J 5 SM/J 6 DMEl 7 DME2
8 DME3
9 DMAl
10 DMA2
11 DIH 12 lCJl 13 1CJ2 14 DIJl 15 DIJ2 16 DIJ3 17 D I T 18 WMP/PAS 19 DPL 20 DEAl 21 2
22 3 23 4
24 DES1
25 2
26 3
27 4
28 5
29 6
30 DEGl 31 DEG2 32 DEW M . musculus
33 PWK/PAS 34 MCB 35 MCS 36 MYB 37 MA1 38 MPW 39 MDS M . castaneus
40 CASA/RK 41 C T T
M . molossinus
42 MOLC/RK
43 BIO.BACI 44 BlO.BAC2
12.I. bactrianus
M. abbotti
45 XBS 46 AYG
M . hortulanus
47 HAH
48 HYP
M . spretus
49 ssc
50 SSG
51 SMA
M. cervicolorlcaroli
52 CVT 53 C K T
2 3 2 2 2 2 2 t*/2 t*/1 1 1 1 1 1 1 1 3 2 2 1 t t*/2 t*/2 1 1 4 t/ 1
t’/l t’/l 4 2 t‘/l
1 1
1 1
1 1
2 2
t t
1 1
1 1
t / l t / l
3 1
t/3 t/l
2 2
1 1
1 1
1 1
1 I
1 1
2 2 2 2
1 1
t t
t t
t/t t/t
t / 3 t / l
t/3 t/l
t / l t / l
t/2 t/2 t/2 t/2
t / l t / l
t / l t / l 113 1
1 1
1 1
2 2 2 2
2 2
2 2
2 2 2
2
t/2 -
1 2
t / l -
2 2
2 2
2 2 2
2 2 2 2
2 2
2 1 2
-
- --
- -t t
t/2 t/2
t/2 t/2
2 2
112 2
2 2
t/2 t/2
t/2
-
112 -
2 2
112
-
t/2 -
t/2 2
2 1
2 1
2 1 2 1 2 1 2 1 2 1
4 5
5 5
4 4
5 3
4 4 5 5
5 4
3 1
3 1
3 1
3 1
3 1
3 1
3 1
2
1
6 7
1 1
2 2
2 2
2
2 1
- 3
3 7
-
1 1
1 1
2 1 5 4
1 1 2 5 3 1 2
314 118
5 t
1 t 1 1 2 t*
2 2
t t
2
t
2 2
t t
1 t
3 1
3 1
2 2
2 1
2 1
5 9
5 9 2 2 1 1 2 2 8 8
2 1
2 1
8 10
8 10
2 2 1 1 3 3 9 8
3 1
3 1
2 2
2 3 2 3
2 3
2 11 2 11
1 1 1
2 2 2 2 2 2 3 3 3 I4
10 10 10
3 1
3 1
3 1
2 2 2
11 12
4 1
5 1
3 2
6 12
6 13
3 4
5 5
3 1
2 1