Copyright Q 1992 by the Genetics Society of America
Biased Gene Conversion, Copy Number, and Apparent Mutation Rate
Differences Within Chloroplast and Bacterial Genomes
C.
William Birky,
Jr.,*and
J.Bruce
Walsht
*Department of Molecular Genetics, The Ohio State University, Columbus, Ohio 43210, and +Department ofEcology and Evolutionary Biology, The University of Arizona, Tucson, Arizona 85721
Manuscript received July 12, 199 1
Accepted for publication November 30, 199 1
ABSTRACT
We investigate the possibility that differences between synonymous substitution rates of organelle and bacterial genes differing only in copy number may be due to conversion bias. We find that the rather large observed difference in the synonymous rates between genes in the single copy and inverted-repeat regions of chloroplasts can be accounted for by a very small bias against new mutants. More generally, differences in the within-organelle fixation probability result in different apparent mutation rates as measured by the expected rate of appearance of cells homoplasmic for new mutants. Thus, differences in intracellular population parameters rather than molecular mechanisms can account for some variation in the apparent mutation rates of organelle genes, and possibly in other systems with variable numbers of gene copies. On the other hand, our analysis suggests that conversion bias is not a likely explanation for relatively low mutation rates observed near the replication origin
of bacterial chromosomes.
,
-I
N their analysis of the relative rates of nucleotide substitution among plant genomes, WOLFE, LI and SHARP (1987) and WOLFE (199 1) observed that the synonymous substitution rate in the chloroplast ge- nome is lower for genes in the inverted repeat (IR) than for genes in the single-copy regions (SC). They suggested that the mutation rate may be lower in the IR. This is conceivable but ad hoc in the absence of a biological precedent or known molecular mechanism that would cause mutagen sensitivity or replication fidelity of a sequence to vary with its copy number. Alternatively, they noted that there is a correction mechanism that maintains the identity of the two copies of the IR and suggest that this correction mechanism may be biased in favor of wild-type alleles. We believe these two hypotheses are the same for most purposes, because the effective mutation rate u for organelle genes is measured as the rate of appear- ance of mutant homoplasmic cells. u is the rate meas- ured after new mutations have been fixed within a cell, and any correction mechanism influences this process of intracellular fixation or loss. T h e likelymolecular mechanisms for correction of the IR are the same as those for concerted evolution of repeated sequences in general: crossing over between sister molecules, and gene conversion. In this note we point out that if gene conversion were biased on average, either toward wild-type or mutant alleles, it would have different effects on genomes or regions of ge- nomes with different copy numbers, as is the case with the IR and SC. If conversion is biased, on average, against mutant alleles, the result would be a lower
Genetics 1 3 0 677-683 (March, 1992)
value of u for sites within an IR relative to sites in the
SC region with the same bias and mutation rate. An evolutionary argument has been made that conversion is likely to be biased against mutant alleles by NAGY-
LAKI and PETITS (1982) and WALSH (1983), who noted that the repair of heteroduplexes is often biased in favor of one base or the other, and hence of one allele or the other, at a given site. A favored base (and allele) is more likely to be fixed in the population. Thus the wild type alleles in a population are likely to be en- riched in such favored bases and thus will tend to be favored over mutant alleles in conversion events.
Another potential example of the effects of biased conversion in an intracellular population of genomes is seen in the data of SHARP et al. (1989) and SHARP (1991), who found that the synonymous substitution rate in enterobacterial genes is lower in the vicinity of the replication origin (oriC) than elsewhere. Noting that the gene copy number is greater in the vicinity of oriC, they hypothesize that postreplication (recom- bination) repair of lesions might be more effective near oriC because the copy number is higher, which in turn might lower the mutation rate and hence the synonymous substitution rate. We examine whether biologically realistic values of bias can account for these two examples of observed copy-number-depend- ent mutation rates.
SUBSTITUTION RATES FOR ORGANELLE GENES WITH BIASED CONVERSION AND
INTRACELLULAR RANDOM DRIFT
contains multiple (one to hundreds) genomes. Thus for a mutant to become fixed in a cell, it must be fixed in an organelle and that organelle subsequently fixed within the cell. It is this fixation within the cell that concerns us. T h e fate of a new mutation within an organelle/cell is determined by two general classes of mechanisms, analogous to random drift and selection in Mendelian populations (BACKER and BIRKY 1985;
BIRKY 1991). Intracellular random drift is the term applied to random changes in allele frequencies within individual cells or organelles. One major mechanism of intracellular random drift is random replication,
i.e., random choice of organelle DNA molecules for replication, perhaps coupled with random degrada- tion (BIRKY 1983). Another mechanism is random pairing of molecules and subsequent gene conversion (BIRKY and SKAVARIL 1976). T h e second factor de- termining the fate of organelle mutations is intracel- lular selection, which causes the frequency of a mutant allele to decrease or, rarely to increase, in the cell (BACKER and BIRKY 1985). Similarly, any bias in gene conversion also acts as a directional force. If both of these directional forces are weak, they can be over- come by the effects of intracellular drift.
Assume that the expected time to fix a new mutant destined to become fixed (whether within the organ- elle, cell, or population) is short relative to the ex- pected waiting time for such fortunate mutants to appear. Under this assumption, the expected substi- tution rate R of alleles at a site approximately equals the rate of appearance of successful new mutants (the number of new mutations per generation times the probability of fixation of each new mutant). With organelle genes, we can decompose the probability of fixation into the probability that the mutant becomes fixed within a cell and the subsequent probability that it is then fixed in the entire population. Our focus here is on mutants with no fitness effects on the replication of molecules, organelles, cells or orga- nisms. Once such a mutant becomes fixed within a cell, it subsequently behaves in a completely neutral fashion, provided organelle inheritance is uniparental. With biparental inheritance intracellular directional forces can further increase the probability of fixa- tion throughout the population beyond their role in generating the initial homoplasmic mutant cell (TAKAHATA 1984; WALSH 1992), so that the ex- pected substitution rate is equal to v , the per gen- eration rate of appearance of cells homoplasmic for a new mutant.
To obtain u, consider a cell consisting of m organ- elles each of which contains ng genomes (DNA mole- cules). We assume there is no genetic exchange be- tween the individual organelles so that a mutation must first become fixed within an organelle, and the homoplasmic organelle must then be fixed in the cell.
AS above, we assume that the expected time until the
loss or fixation of a new mutation within an organelle (and also fixing an organelle
type
within a cell) is small relative to the expected waiting time between success- ful mutations. Let p be the mutation rate per genome per generation at the site being followed, so that within each organelle prig new mutant alleles arise each generation, and let Uo(x) be the probability of fixing a new mutant allele within an organelle given that its initial frequency is x. Thus the expected num- ber of new mutations destined to be fixed within an organelle that arise each generation is pn,V,(l/ng).Since there are no organelles in each cell, the rate of appearance of mutant homoplasmic organelles within a cell is no[pngUo(l/ng)], giving the expected rate of mutant homoplasmic cells as
u = ( n o [PflgVo( 1 /ng)l)~c( 1 / n o ) (1)
where Uc( l/no) is the probability of fixing an organelle (starting as a single copy) within a cell. To facilitate matters, we assume throughout that the mutant ge- nome has no effect on the replication and/or trans- mission of the organelle within a cell, so that fixation of a given organelle type throughout a cell is by drift alone, giving Uc(x) = x and
= P gU o ( 1 /.g) (2)
which is simply the expected number of mutants des- tined to become fixed in a single organelle each gen- eration.
BIRKY, MARUYAMA and FUERST (1983) pointed out that for completely neutral mutations the probability of fixation within an cell is equal to the initial fre- quency of a mutant allele in the cell, i e . , to the reciprocal of the total number of gene copies in the cell. Consequently, the observed mutation rate, i.e.,
the rate of appearance of homoplasmic mutant cells,
is u = (nongp)(l/nong) = p, the mutation rate of an
individual organelle genome. Hence in the absence of any intraorganelle directional effects, the observed mutation rate is independent of the number of
ge-
nomes in the organelle.
Directional effects, such as intraorganelle selection biased on differences in replication rates (TAKAHATA
1984), make v sensitive to copy number (BIRKY 199 1).
To see how biased gene conversion influences u, as- sume that there are ng genomes per organelle and let
b be the probability that a wild-type
+
allele is con- verted to a mutant m allele during a conversion event that changes allelic type. Similarly let 1-
b be the probability that m is converted to+.
If b>
1/2, the mutant allele is favored by conversion, while the wild- type allele is favored if b<
1/2. NAGYLAKI and PETES(1 982) found that the probability of fixation (starting
Gene Conversion and Mutation Rates 679
a gene family within a single chromosomal lineage is 1
-
r' l - bT i =
-
where r =-
b ' (3) 1
-
r"'A more detailed analysis on the probability of fixation of a new organelle mutant considering simultaneous directional selection due to genotypic-specific differ- ences in replications rates (both genomes within or- ganelles and organelles within cells), biased gene con- version, biparental inheritance, and some exchange between organelle) has been developed and will be given elsewhere (WALSH 1992). If we assume that the fixation of one allele (either mutant or wildtype) is due primarily to gene conversion as opposed to ran- dom replication and partitioning, this result can be directly applied to biased conversion between the n copies of a gene in an organelle. Note that for genes in the SC, n = nIrl while for genes in the IR, n =
272,.
Suppose bias 1s small, so that b = (1/2)
+
6 whereI 6
I
<<
1. In this caseUsing this and the approximation (1
-
x)" k: exp(-nx) whenI
xI <<
1, the probability of fixation within an organelle starting from a single new mutant is1 - r 46
1
-
r" 1-
exp(-46n)LJo(l/n) =
-
x (4)giving the rate of appearance of newly arisen mutant homoplasmic organelles as
l - r 46
u = p n c - F = pnl
-
exp(-46n)* (5) If mutants differ in the amount of conversion bias, the expected rate is given by taking the expectation of ( 5 ) over the distribution of 6 values in new mutants. Under our assumption that mutants have no effect on the replication rates of genomes, organelles, cells, or organisms, the substitution rate of alleles at this site in the population is u. Thus when conversion bias is present, u depends on copy number, and the substi- tution rate varies in organelles with different numbers of genomes. In particular, if mutant alleles have the same amount of bias, the ratio of substitution rates for organelles with n and n' genomes is""
u' n' 1
-
r" n' 1-
exp(-46n) u n 1-
P' n 1-
exp(-46nf)'-
x - (6)Perhaps more interestingly (as we show below), when bias is present, genes within the same genome differ- ing in copy number will also show differences in the substitution rates.
A N APPLICATION TO PLANT PLASTIDS
As an application of the above theory, we consider how conversion bias can influence intraorganelle fix-
ation probabilities in chloroplasts (plastids). In plants, chloroplasts are often inherited from only one parent, frequently because plastids from the other parent are excluded from the zygote (HAGEMANN and SCHRODER
1989; NEALE, MARSHALL and SEDEROFF 1989). More- over, recombination of plastid genes has not been detected in plants showing biparental inheritance
[R. A. E. TILNEY-BASSETT (personal communication) and CHIU and SEARS (1985) and references therein). When plant protoplasts were fused and plants regen- erated with stringent selection for recombinant plastid types, only one recombinant plant was found (MED- CYESY, FEJES and MALICA 1985), but it had a plastid genome that resulted from numerous recombination events (FEJES, ENGLER and MALICA 1990). This sug- gests that plant plastids have the enzymatic machinery for recombination, but that plastids having different genotypes rarely fuse. Further evidence for intermo- lecular recombination in plant chloroplasts comes from experiments in which plastid genomes were sta- bly transformed with fragments of plastid DNA (SVAB, HAJDUKIEWICZ amd MALICA 1990), a process that almost certainly involves homologous recombination such as has been demonstrated for transformation in Chlamydomonas (NEWMAN et al. 1990). Therefore we can focus on events that lead to fixation or loss of a mutation within the plastid where it occurred or in a product of the division of that plastid. Once a muta- tion is fixed in a plastid, we assume that it has no effect on rates of plastid or cell replication and that inheritance is uniparental so that the population sub- stitution rate is given by u computed from (2). Let V I R
be the expected rate of appearance of plastids that are homoplasmic for mutations at a site in the inverted repeat and USC be the rate for a site in the single-copy
region. As above, we assume that the actual time to fixation is short relative to the time between appear- ance of mutants destined to be fixed.
and
( 1 ) Conversion within inverted repeats is very rapid
W
(2) Rapid within-organelle conversion and slower between-organelle conversion
rn-m
W
+ +
U
A
In- + I - b
<:+
- w
conversion
h
1 -c mhm+ + n
rnL+ I - b
\+-+ +-+ ( I - b ) ( l - c )
w w
FIGURE 1.-Model I for intraorganelle drift assumes that con- version between sites in the IR within a genome is sufficiently rapid that only m/m and
+/+
genomes exist in the organelle.conversion occurs primarily within molecules, vs.
equal rates of conversion within and between mole- cules. Different combinations of these extreme as- sumptions are used in three different models.
Model I assumes that there is very rapid correction between variant members of the IR of a single ge- nome-when a mutation to allele m occurs within a site in the IR, it is very rapidly corrected so that the IR remains symmetric (Figure 1). Under this model, all genomes within an organelle are either completely mutant m/m or wild type
+/+.
Following the appear- ance of a mutation at one site in the IR, with proba- bility b =(1/2)
+
6 the IR is corrected to give two mutant copies (mlm), while with probability 1-
b it is converted back to wild-type (+/+). Hence if p is the per-site mutation rate, the rate of appearance of mu- tant ( m / m ) genomes within a plastid is 2pbn = (1+26)pn. Model I further assumes that intraorganelle drift and the fixation of alleles within an organelle is due to random replication and partitioning of DNA molecules rather than to gene conversion. Then Uo(x)
= x, and the expected substitution rate at a site in the IR (for alleles with no effect on organelle or cell replication rates) is uIR = (1
+
26)pn, while the ex-pected rate within a single copy region is usc = pn,
giving
Model I1 also assumes that intramolecular IR cor- rection is essentially instantaneous, but that biased conversion, rather than random replication and par-
titioning of DNA molecules, is the primary cause of
the fixation of alleles within organelles. As shown in Figure 1 , we can easily allow for the possibility of differences in the bias between the intragenomic and intergenomic conversion processes. Here c = (1 /2)
+
t measures the between-genome bias for the mutant
allele and b = (1/2)
+
6 measures the within-genome bias. From Figure 1, for sites within the IR the ratioof the probability of conversion losing a m/m genome to the probability of gaining a m/m genome is ( 1
-
c)(l-
b ) / ( c b ) , while for single copy sites, this ratio is (1-
c)/c. Using results from the theory of birth and death processes, the probability that all genomes within an organelle are fixed for a mutant (given that we start with i mutant genomes) is given by Equation3 using
r=-- 1 - 6 1 - C
b c
=
1-
4(6+
e ) (84for sites within the IR and
for single copy sites. Thus for single copy sites
U o ( l / n )
=
461
-
exp(-4tng) ( 9 4while for sites within the IR
For single copy sites the mutation rate is prig, while for sites within the IR the mutation rate (to m/m genomes) is pn(1
+
26). Combining these with (sa) and (9b) gives the ratio of substitution rates asVIR
-
=
( 1+
26)-
t+
6 1-
exp(-4tng)t 1
-
exp(-4(t+
6)ng)' (1 0 4usc
Assuming that the within- and between-genome biases are identical (t = 6),
-
VIR=
2(1+
26) 1-
exp(-46ng)usc 1
-
exp(-86ng)' (1 Ob)Model 111 assumes that (i) conversion rates within and between genomes in a plastid are roughly equal, and (ii) the fixation of alleles within an organelle is due to conversion. With biased gene conversion, the proba- bility of fixation within an organelle is given by (4) where n = 2ng for sites in the IR (since each genome contains two copies), while n = ng for sites in the SC region. Consequently, from (6) the ratio of substitu- tion rates in this case is
-
VIR 1-
exp(-46ng)Gene Conversion and Mutation Rates 68 1
B
.
YL
I
c .dY
B
5 k - 23
+
+
k - 4 k - 88
I
4
*.
0 0
3
.I
9
2 10
-0.4 -0.3 -0.2 -0.1 0.0 0.1 0.2 0.3 0.4
6
FIGURE 2.-The ratio of substitution rates within the IR us. SC regions, uIR/uSC as a function of x- 4nb where A is the copy number
and 6 is the gene conversion bias (based on Equation 11).
For small 6, this is essentially identical to (lob). As Figure 2 shows, the ratio uIR/uSC ranges between 0
when n6 is small and negative (conversion favors the wild-type allele) and 2 when n6 is large and positive (conversion favors the mutant allele).
Can gene conversion, biased against new mutations, account for the lower synonymous substitution rate in the inverted repeat of plant chloroplast DNA? T h e number of genomes in a meristematic cell proplastid, ng, is unknown and probably highly variable; we as- sume it is on the order of 10 to lo2, similar to or smaller than the number in mature chloroplasts (order of 10 to 10'; reviewed by ROSE 1988). T h e weighted mean of the ratio UIR/USC for all the sequence compar-
isons made by WOLFE, LI and SHARP (1991) is 0.26, while a more recent calculation by WOLFE (1991) based on more sequences gives 0.1 9. Under the as- sumption that alleles are fixed in organelles mainly by random replication and partitioning (model I and Equation
7),
uIR/usc = 0.19 = 1+
26 giving 6 = -0.405, an extremely large and biologically unrealistic conver- sion bias. Under the assumption that conversion is the primary force that makes organelles homoplasmic (models I1 and I11 and Equations 10b and 11; we use Equation 1 1 since (1 Ob) and (1 1) are essentially equiv- alent whenI
6I
C< 0), uIR/uSC = 0.19 if 6 = -0.056and ng = 10, or if 6 = -0.0056 and ng = 100. Biased conversion of this magnitude, especially with the sec- ond value, would probably not be detected in experi- ments performed to date. In the absence of tetrad analysis, a conversion bias would be detectable only in plants with biparental inheritance of plastid genes, where it would be seen as an bias in favor of wild type alleles, averaged over reciprocal crosses involving many different newly arisen mutant alleles and many different nuclear and plastid genetic backgrounds.
Large-scale crosses would be technically difficult to perform with synonymous substitutions, and if the crosses were done with nonsynonymous mutations, any observed bias would be attributed to selection at the level of organisms.
We conclude that gene conversion with a bias against mutant alleles can potentially explain the dif- ference in synonymous substitution rate between IR and SC regions of plant chloroplast genomes. A com- parison of model I to models I1 and I11 also shows that it is necessary to have substantial biased gene conversion between molecules in an organelle, while the organelles are still heteroplasmic. Alleles must be fixed in organelles primarily by conversion as opposed to random replication and partitioning. Our compar- ison of models 11 and I11 also suggests that possible differences in the rates of conversion between IRs within and between molecules would be of little im- portance in this context.
An interesting possibility is that differential effects of gene conversion in the IR us. SC may be accen- tuated by differential recombination frequencies. THOMAS and ROTHSTEIN (1 99 1) review a substantial body of evidence that transcription promotes recom- bination, including gene conversion. T h e IR contains the rRNA genes, which may be transcribed more
frequently than genes in the SC. LEMIEUX, TURMEL and LEMIEUX (1 990) found that recombination junc- tions occurred more frequently in the IR than in the SC in Chlamydomonas crosses. If conversion events are initiated more frequently in the IR than in the SC, the population dynamics of the IR would follow model I while those in the SC would follow model 11. This in turn would enhance the differential effects of biased gene conversion.
A N APPLICATION TO BACTERIAL GENOMES T o apply these ideas to the case of the bacterial chromosomes studied by SHARP et al. (1 989), we limit discussion to conversion within the cell in which the mutant originally occurred. Further opportunities for conversion between mutant and wild type alleles may arise due to sexual reproduction, but the frequency of bacterial sex in nature is not well known. Within the cell, random drift of allele frequencies is due to gene conversion alone. If n is the number of copies of a site near the origin, then (5) implies
uOR = pn
-
1 - r1
-
r"giving the relative substitution rate of sites near the origin with n copies compared to a single copy site within the same cell (such as near the terminator of
replication) as
W. Birky
-4 -3 -2 -1 0 1 2 3 4
X
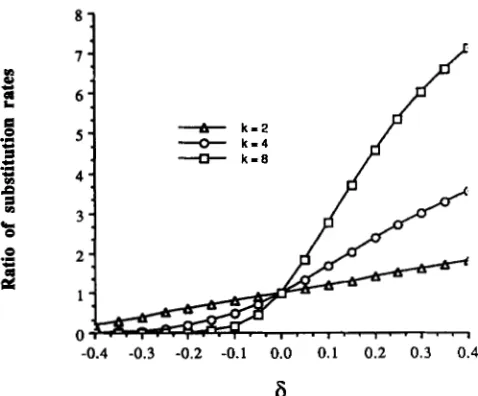
FIGURE 3.-Ratio of substitution rates between overreplicated regions (with k copies) near the origin of replication and single-copy genomes within a single cell uOR/u when mutant alleles have conver- sion bias 6.
Figure 3 plots this ratio as a function of 6 and n. T o explain a twofold difference in mutation rates between sites near the origin and near the terminator, such as observed by SHARP et al. (1989) and SHARP
(1991), solving (12b) for u O R / U = 0.5 gives 6 = -0.250,
-0.098 or -0.043 for n = 2, 4 or 8 (respectively). While the exact number of gene copies per cell in nature is unknown (since it depends on temperature and nutritional factors), strong bias is required in any of these cases and bias as an explanation for the observed difference seems unreasonable.
DISCUSSION
Mutation rates are usually measured in terms of the rate of appearance of phenotypically mutant cells. It is not always appreciated that when cells have more than one copy of a genome, as is the case with mito- chondria and chloroplasts, what is first produced is a cell with one mutant and many wild type alleles, which is unlikely to have a mutant phenotype. A cell will not be counted as mutant until the number of mutant alleles has increased to the point where the cell is phenotypically mutant. In many experimental proto- cols, such as the fluctuation test, a cell will not count as mutant until the mutant phenotype is thereafter inherited clonally; this requires that the mutant allele be fixed in the cell. Thus the observed mutation rates are determined not only by mutation processes at the molecular level but also by intracellular population dynamics that control fixation or loss of the mutant allele. Intracellular random drift plays an important role, but it is not directional, i.e., it does not favor the fixation of mutant or wild type alleles, hence it does not affect the observed mutation rate. Intracellular selection is a directional force that affects the fixation probability and hence the observed mutation rate. Here we show that biased gene conversion has the
same kind of effect. We further show that the number of gene copies in a cell or organelle influences the conversion dynamics and hence influences the ob- served mutation rate. For the specific case of plant chloroplast DNA, we show that a very small bias in favor of wild-type alleles can explain the observed lower mutation rate in the inverted repeat, provided that the fixation of alleles within individual plastids is due primarily to gene conversion rather than to ran- dom replication and partitioning. We note that such a bias might result from the long-range evolutionary tendency to fix and retain bases that are favored in the repair of mismatched base pairs during gene con- version, as suggested by NAGYLAKI and PETES (1982) and WALSH (1983). This argument is analogous to the explanation of why new mutations are more often detrimental than advantageous, just as biased conver- sion is formally analogous to selection. T h e mutation rate difference between IR and SC regions in cpDNA can be considered indirect evidence for such a bias, which would be difficult to measure directly.
Let us briefly consider some alternative hypotheses that might explain the observed copy number effects. One possibility is that DNA repair mechanisms may be more efficient at higher copy number. Most of the classes of repair mechanisms known to operate in bacteria and known or suspected to operate in eukar- yotes (reviewed in LEWIN 1990) involve only one molecule at a time and hence are not dependent on copy number; these include damage reversal by spe- cific enzymes, excision repair, mismatch repair, SOS
repair and error-prone repair. Recombination repair, which was suggested by SHARP et al. (1989) as a possible explanation for their observations on enter- obacteria, appears to involve the donation of a single strand by an undamaged molecule to fill a gap in a damaged molecule. The process also requires gap- filling DNA synthesis which will produce single base pair substitutions if it is error-prone. Recombination repair might be more efficient when there are more copies of a region to serve as donors, but this will serve to increase rather than decrease the observed mutation rate because a molecule with an unrepaired gap may be lost from the population rather than being detected as mutant. Some repair processes are in- duced by damaged DNA; for these, a higher copy number may result in more efficient repair. This will not explain differences in mutation rates between different genomic regions in a single cell or organelle, but it might result in rate differences among organ- isms or cells with different numbers of genomes. Higher copy numbers could result in either higher or lower mutation rates, depending on whether mole- cules with unrepaired damage were lethal or viable and mutant.
Gene Conversion and Mutation Rates 683
count for observed differences in mutation rates be- tween regions at varying distances from the replica- tion origin of bacterial chromosomes. But in this case the necessary conversion bias is extremely large, and probably unrealistic. There is a plausible alternative mechanism: hemi-methylation of newly replicated DNA favors the repair of mismatched base pairs (rep- lication errors) to re-create the wild type allele (PUK- KILA 1987). CAMPBELL and KLECKNER (1 990) showed
that genes closer to oriC remain hemimethylated longer, which might permit more methyl-directed re- pair and a lower mutation rate.
Our most important conclusion is that biased gene conversion can cause systems with different numbers of genome copies to have different realized mutation rates. Besides the repeated vs. single copy regions in chloroplast genomes and the oriC effects discussed here, this might apply to repeats in mitochondria DNA; to dispersed repeats in nuclear genomes; to species with different ploidy levels of nuclear DNA; to plasmids with different intracellular copy numbers; and to intracellular symbionts. We modeled the case of plant chloroplasts in which there is little or no fusion between organelles, but biased conversion will also affect chloroplast and mitochondrial mutation rates in organisms where the organelles regularly ex- change genomes. However, the population parame- ters and kinetics of conversion and fixation, and hence the magnitude of the effect of biased conversion, may differ.
We thank WEN-HSIUNG LI, PAUL SHARP, KEN WOLFE and ANDY CLARK for useful comments on the manuscript. This work was partially supported by National Institutes of Heath grant GM37912
(to J.B.W.).
LITERATURE CITED
BACKER, J. S., and C. W. BIRKY, JR., 1985 The origins of mutant cells: mechanisms by which Saccharromyces cerevisiae produces cells homoplasmic for new mitochondrial mutants. Curr. Ge- net. 9 627-640.
BIRKY, C. W., JR., 1983 Relaxed cellular controls and organelle heredity. Science 222: 468-475.
BIRKY, C. W., JR., 1991 Evolution and population genetics of organelle genes: mechanisms and models, pp. 112-1 34 in Eu- olution at the Molecular Level, edited by R. K. SELANDER, A. G .
CLARK and T. S. WHTTAM. Sinauer Associates, Sunderland, Mass.
BIRKY, C. W., JR., T. MARUYAMA and P. FUERST, 1983 An a p proach to population and evolutionary genetic theory for genes in mitochondria and chloroplasts and some results. Genetics
BIRKY, C. W., JR., and R. V. SKAVARIL, 1976 Maintenance of genetic homogeneity in systems with multiple genomes. Genet. Res. 27: 249-265.
CAMPBELL, J. L., and N. KLECKNER, 1990 E. coli orjc and the
dnaA gene promoter are sequestered from dam methyltrans- f e m e following the passage of the chromosomal replication fork. Cell 62: 967-979.
103: 513-527.
CHIU, W.-L., and B. B. SEARS, 1985 Recombination between chloroplast DNAs does not occur in sexual crosses of Oenothera.
Mol. Gen. Genet. 198: 525-528.
FEJES, E., D. ENGLER and P. MALIGA, 1990 Extensive homologous chloroplast DNA recombination in the pt14 Nicotiana somatic hybrid. Theor. Appl. Genet. 7 9 28-32.
HAGEMANN, R., and M.-B. SCHRODER, 1989 The cytological basis of the plastid inheritance in angiosperms. Protoplasma 152:
57-64.
LEWIN, B., 1990 Genes N. Oxford University Press, New York. LEMIEUX, B., M. TURMEL and C. LEMIEUX, 1990 Recombination
of Chlamydomonas chloroplast DNA occurs more frequently in the large inverted repeat sequence than in the single-copy regions. Theor. Appl. Genet. 79: 17-27.
MEDGYESY, P., E. FEJES and P. MALIGA, 1985 Interspecific chlo- roplast recombination in a Nicotiana somatic hybrid. Proc. Natl. Acad. Sci. USA 82: 6960-6964.
NAGYLAKI, T., and T. D. PETES, 1982 Intrachromosomal gene conversion and the maintenance of sequence homogeneity among repeated genes. Genetics 1 0 0 315-337.
NEALE, D. B., K. MARSHALL and R. R. SEDEROFF,
1989 Chloroplast and mitochondrial DNA are paternally in- herited in Sequoia sempewirens D. Don Endl. Proc. Natl. Acad. Sci. USA 8 6 9347-9349.
NEWMAN, S. M., J. E. BOYNTON, N. W. GILLHAM, B. L. RANDOLPH- ANDERSON, A. M. JOHNSON and E. H. HARRIS,
1990 Transformation of chloroplast ribosomal RNA genes in Chlamydomonas: Molecular and genetic characterization of integration events. Genetics 126: 875-888.
PUKKILA, P. J., 1987 Telling right from wrong: a role for DNA methylation. Trends Genet. 3: 1-2.
ROSE, R. J., 1988 The role of membranes in the segregation of plastid DNA, pp. 1 7 1 - 195 in The Division and Segregation of
Organelles, edited by S. A. BOFFEY and D. LLOYD. Cambridge University Press, Cambridge.
SHARP, P. M., 1991 Determinants of DNA sequence divergence between Escherichia coli and Salmonella typhimurium: codon usage, map position, and concerted evolution. J. Mol. Evol. 33:
SHARP, P. M., D. C. SHIELDS, K . H. WOLFE and W.-H. LI,
1989 Chromosomal location and evolutionary rate variation in enterobacterial genes. Science 246 808-8 10.
SVAB, Z., P. HAJDUKIEWICA and P. MALIGA, 1990 Stable transfor- mation of plastids in higher plants. Proc. Natl. Acad. Sci. USA
TAKAHATA, N., 1984 A model of extranuclear genomes and the substitution rate under within-generation selection. Genet. Res.
THOMAS, B. J., and R. ROTHSTEIN, 1991 Sex, maps, and imprint- ing. Cell 64: 1-3.
WALSH, J. B., 1983 Role of biased gene conversion in one-locus neutral theory and genome evolution. Genetics 105 461-468.
WALSH, J. B., 1992 Intracellular selection, conversion bias, and the expected substitution rate of organelle genes. Genetics (in press).
WOLFE, K. H., 1991 Protein-coding genes in chloroplast DNA: compilation of nucleotide sequences, data base entries, and rates of molecular evolution, pp. 467-482 in Cell Culture and Somatic Cell Genetics of Plants, Vol. 7 8 . The Photosynthetic Appa- ratus: Molecular Biology and Operation, edited by L. BOGORAD
and I. K. VASIL. Academic Press, New York.
WOLFE, K. H., W.-H. LI and P. M. SHARP, 1987 Rates of nucleo- tide substitution vary greatly among plant mitochondrial, chlo- roplast and nuclear DNAs. Proc. Natl. Acad. Sci. USA 84:
23-33.
87: 8526-8530.
44: 109-1 16.
9054-9058.