Copyright1999 by the Genetics Society of America
Localization of Single- and Low-Copy Sequences on Tomato Synaptonemal
Complex Spreads Using Fluorescence
in Situ
Hybridization (FISH)
Daniel G. Peterson,* Nora L. V. Lapitan
†and Stephen M. Stack
‡*Department of Crop and Soil Sciences, University of Georgia, Athens, Georgia, 30602 and†Department of Soil and Crop Sciences and‡Department of Biology, Colorado State University, Fort Collins, Colorado 80523
Manuscript received November 13, 1998 Accepted for publication February 1, 1999
ABSTRACT
Fluorescence in situ hybridization (FISH) is a powerful means by which single- and low-copy DNA sequences can be localized on chromosomes. Compared to the mitotic metaphase chromosomes that are normally used in FISH, synaptonemal complex (SC) spreads (hypotonically spread pachytene chromo-somes) have several advantages. SC spreads (1) are comparatively free of debris that can interfere with probe penetration, (2) have relatively decondensed chromatin that is highly accessible to probes, and (3) are about ten times longer than their metaphase counterparts, which permits FISH mapping at higher resolution. To investigate the use of plant SC spreads as substrates for single-copy FISH, we probed spreads of tomato SCs with two single-copy sequences and one low-copy sequence (ca. 14 kb each) that are associated with restriction fragment length polymorphism (RFLP) markers on SC 11. Individual SCs were identified on the basis of relative length, arm ratio, and differential staining patterns after combined propidium iodide (PI) and 49,6-diamidino-2-phenylindole (DAPI) staining. In this first report of single-copy FISH to SC spreads, the probe sequences were unambiguously mapped on the long arm of tomato SC 11. Coupled with data from earlier studies, we determined the distance in micrometers, the number of base pairs, and the rates of crossing over between these three FISH markers. We also observed that the order of two of the FISH markers is reversed in relation to their order on the molecular linkage map. SC-FISH mapping permits superimposition of markers from molecular linkage maps directly on pachytene chromosomes and thereby contributes to our understanding of the relationship between chromosome structure, gene activity, and recombination.
F
OR most plant species, what is known about the Stack 1995). The most direct means of determining order of loci on chromosomes is based almost en- the location of genes on chromosomes is through the tirely on genetic linkage maps. Such linkage maps are use of fluorescence in situ hybridization (FISH). In this generated by producing multi-hybrid crosses and de- technique hapten-labeled DNA probes are hybridized termining the relative frequency of recombination be- to chromosomes that have been spread on glass micro-tween genes or molecular markers (Tanksley et al. scope slides, and antibodies or other affinity reagents 1989;Paterson1996). The physical location of genes conjugated to fluorochromes are used to detect directly on chromosomes is of great interest because chromo- or indirectly sites of hybridization (Trask1991). FISH some structure profoundly influences gene activity (see with repetitive sequences as probes has been widelyre-LoheandHilliker1995;WallrathandElgin1995; ported for both animal and plant chromosomes, and
ZuckerkandlandHennig1995 for reviews). However, single-copy FISH to mammalian chromosomes is fairly linkage maps cannot be superimposed on chromosomes routine (e.g.,Landegent et al. 1987; Lawrence et al.
because map distances are not proportional to physical 1988;Lichteret al. 1991; Henget al. 1992). Because
distances (SturtevantandBeadle1939;Khush and most techniques used to prepare plant chromosomes
Rick1968; Moore andSherman1974;Flavell et al. leave overlying debris that interferes with probe penetra-1985). The relative scarcity of genes and crossing over tion, single-copy FISH to plant chromosomes has proven in heterochromatin is partly responsible for this discrep- considerably more difficult (Lehfer et al. 1993;Jiang
ancy, but crossing over is not evenly distributed in et al. 1995). However, now there are a few laboratories
euchromatin either (Snape et al. 1985; Dooner 1986; that have overcome this obstacle and are successfully
TsujimotoandNoda1990;CurtisandLukaszewski localizing single-copy sequences on plant chromosomes 1991;Kotaet al. 1993;Stacket al. 1993;Shermanand by FISH (e.g., Leitch and Heslop-Harrison 1993;
Hansonet al. 1995;Jianget al. 1995;Go´ mezet al. 1997).
FISH is usually performed on mitotic metaphase chro-Corresponding author: Daniel G. Peterson, Plant Genome Mapping mosomes, but there are reasons to believe that
pachy-Laboratory, University of Georgia, Riverbend Research Center, Rm.
tene (meiotic) chromosomes may be better substrates.
162, 110 Riverbend Rd., Athens, GA 30602.
E-mail: [email protected] Each pachytene chromosome (bivalent) is composed of
and TG523 are single-copy markers, Southern blots of TG400 two homologous chromosomes that are joined along
indicate that it may be present in more than one copy per their entire length by a proteinaceous scaffold called
haploid genome (S. D.Tanksley, personal communication; the synaptonemal complex (SC;Moses1968). Because also see SolGenes web site, http://geneous.cit.cornell.edu/ each homologue contains two chromatids, there are solgenes/aboutsolgenes.html/). Transformation of the com-petent E. coli strain DH5a, plasmid isolations, and verification four closely associated copies of each locus available for
of RFLP marker size by restriction analysis were performed hybridization on a bivalent. In comparison, there are
using standard methods (Sambrooket al. 1989). The polymer-only two nearby copies of each locus available for FISH
ase chain reaction was used to concomitantly amplify and on a metaphase chromosome. In spreads of pachytene label RFLP markers with digoxigenin (DIG;Rashtchianand chromosomes that have been prepared to reveal SCs Mackey1992), and the resulting probes were used to screen a Clonetech (Palo Alto, CA) tomato lambda (l) genomic (SC spreads), chromatin extends as a diffuse cloud
library (cat. no. FL 1082d). Plating and filter preparation were around each SC. The loops of DNA extending from the
performed as described in the Clonetech Lambda Library SC (e.g., Weithand Traut1980) appear to be more
Protocol Handbook (PT1010-1). For each plate, an original accessible to FISH probes than the DNA of condensed and a duplicate filter were prepared. Saline sodium citrate metaphase chromosomes (MoensandPearlman1989, (SSC) solutions of various strengths were produced by diluting a 203SSC stock solution (3mNaCl, 0.5msodium citrate, pH 1990;Henget al. 1994;SolariandDresser1995), and
7.0) with deionized water. Filter hybridization and detection SC spreads can be prepared relatively free of overlying
of positive clones were performed according to the Boehringer debris. Additionally, pachytene chromosomes are 5–15 Mannheim (Indianapolis) DIG/Genius User’s Manual times longer than corresponding metaphase chromo- (see http://biochem.boehringer-mannheim.com/prod_inf/ somes (RamannaandPrakken 1967;Stack1984), so manuals/dig_man/dig_toc.htm/) with modifications as sug-gested by A. S. N.Reddyand I. S.Day(personal communica-many closely associated loci that are not resolvable by
tion). Briefly, nylon filters (Boehringer Mannheim) were FISH on metaphase chromosomes should be resolvable
placed in standard hybridization buffer [53 SSC, 0.1% N-on pachytene chromosomes. lauroylsarcosine, 0.02% sodium dodecyl sulfate (SDS), 1.0% Here we report high resolution localization of two w/v Boehringer Mannheim blocking reagent, 0.01mmaleic acid, 0.015mNaCl, pH 7.5] for 1 hr at 558, incubated overnight
single-copy sequences and one low-copy sequence on
at 558 with gentle shaking in standard hybridization buffer tomato SC 11 using FISH. This is the first report of
containing 0.01 mg/ml heat-denatured DIG-labeled probe, single-copy FISH to SC spreads and one of only a few washed twice in 23SSC containing 0.1% w/v SDS (558, 5 min studies in which FISH has been used to study the rela- each wash), washed twice in 0.53SSC containing 0.1% SDS tionship between genetic linkage and chromosome (558, 15 min each wash), rinsed for 1 min in washing buffer (0.1 mmaleic acid, 0.15 m NaCl, 0.3% Tween-20, pH 7.5), morphology in plants (e.g., Pedersen and
Linde-placed in blocking solution (0.1mmaleic acid, 0.15mNaCl,
Laursen1995;Pedersenet al. 1995). Tomato
(Lycopersi-1% w/v blocking reagent) for 1 hr at 208with gentle shaking,
con esculentum Mill., recently renamed Solanum
lycopersi-and incubated in a 1:2500 dilution of anti-DIG-alkaline
phos-cum L.) was used for this study because all 12 of its SCs phatase (750 units/ml, Boehringer Mannheim) in blocking
are identifiable on the basis of relative lengths and arm buffer for 45 min (208with gentle shaking). After incubation with the primary antibody, filters were washed twice in 500 ratios (Sherman andStack 1992, 1995) and because
ml washing buffer (15 min each wash), placed in detection tomato is a true diploid of agronomic importance (Rick
buffer [100 mmTris-(hydroxymethyl)-aminomethane, 100 mm
1991). Tomato SC 11 was chosen for hybridization be- NaCl, pH 9.5] for 2 min, and incubated in the dark without cause it is one of the shortest tomato chromosomes, agitation in 200 ml of detection buffer containing 900ml of and its corresponding linkage group contains 16 sepa- nitrobluetetrazolium solution (75 mg/ml in 70% dimethyl-formamide, Boehringer Mannheim) and 700ml of 5-bromo-rate RFLP loci, several of which are associated with
4-chloro-3-indoyl phosphate toludinium salt solution (50 mg/ known genes (Tanksleyet al. 1992). Combined
propid-ml in 100% dimethylformamide, Boehringer Mannheim). ium iodide and 49,6-d iamidino-2-phenylindole (CPD) Afterz20 min, positive plaques (small blue “o-shaped” rings) staining was employed to facilitate chromosome identi- began to appear on filters. Filters with a positive plaque in the same location on both the original and duplicate filters fication and aid in relating hybridization sites to
chro-were thoroughly washed in deionized water, and correspond-mosome structures. Our results suggest that FISH to SC
ing plaques were removed from petri plates as described by spreads (SC-FISH) can be used to construct
comprehen-Sambrooket al. (1989). Secondary and tertiary screens were
sive maps of single-copy sequences on pachytene chro- performed as described above. For each of the three starting mosomes. RFLP markers (TG46, TG400, and TG523), one correspond-ing lambda clone (designatedlTG46,lTG400, andlTG523, respectively) was selected for use in FISH.
Probe preparation:The QIAGEN (Valencia, CA) Lambda MATERIALS AND METHODS
Maxi Kit was used to isolate DNA fromlTG46,lTG400, and
Screening the lambda tomato genomic library:Plasmids con- lTG523. The DNA was digested with BamHI or EcoRI, and taining the tomato restriction fragment length polymorphism tomato insert DNA fragments were separated fromlarms by (RFLP) markers TG46, TG400, and TG523 were provided by gel electrophoresis. Bands containing tomato insert DNA were S. D. Tanksley (Cornell University). TG523 flanks the jointless excised from 1% w/v agarose gels, and the QIAGEN Qiaex gene (Winget al. 1994), and TG400 is linked to the hairless II Kit was used to isolate DNA from agarose. The GIBCO BRL gene (Tanksley et al. 1992). To date, TG46 has not been (Rockville, MD) BioNick Kit was used to label DNA with biotin. closely associated with a particular gene or phenotype. While Each biotin-labeled probe was placed in its own 1.5-ml
Tomato synaptonemal complex spreads:SC spreads were citric acid, 164.7 mmNa2HPO4, pH 7.0) containing 0.5mg/ml 49,6-diamidino-2-phenylindole (DAPI), 1 mg/ml propidium prepared as described byPetersonet al. (1996) with several
modifications. Briefly, tomato (cv. Cherry) anthers were iodide (PI), or 1mg/ml PI and 0.5mg/ml DAPI (i.e., CPD). Slides were washed for 30 sec in deionized water and allowed to placed in 200ml of sugar-salt medium [0.56 mmKH2PO4, 0.1
mmacid PIPES, 0.2% w/v potassium dextran sulfate, 1 mm air-dry for 5 min. Ten microliters of freshly prepared antifade medium (aqueous 50 mmTris, 50% glycerol, 1 mg/ml phenyl-CaCl2, 0.7 mmannitol, 1% w/v polyvinylpyrrolidone (Mr 5
10,000), pH 5.1] containing 3 mg desalted cytohelicase (Sepra- enediamine) was placed on each slide, and a 22-350-mm coverglass was added. Coverglasses were sealed onto slides with cor, Marlborough, MA). The upper tips of the anthers were
cut off, and anther bottoms were allowed to digest in the dark fingernail polish. Fluorescence and phase-contrast microscopy were performed using an Olympus Provis AX70 microscope for 10 min. Drops of 45% acetic acid were placed on new
microscope slides (Corning, Inc., Corning, NY), and slides equipped with an Olympus UM-51005 filter cube for simulta-neous visualization of PI and FITC, a U-MNV filter cube for were wiped dry with a Kimwipe. Slides then were
glow-dis-charged according to Dubochet et al. (1971) except that observation of DAPI and CPD staining, and a U-NG filter cube for visualizing PI alone. Photographs were taken using a PM-air rather than amylamine vapors was introduced into the
discharge chamber. Dissecting needles were used to squeeze C35DX camera and Kodak Royal Gold 400 film.
CPD karyotype: Both freshly fixed SC spreads and SC microsporocytes from anthers, and microsporocytes were then
allowed to digest for an additional 10–15 min. A 0.5-ml aliquot spreads that had been used in FISH were CPD stained, and complete (or nearly complete) late pachytene sets were photo-of the protoplast suspension was drawn into a siliconized
micro-pipette and gently blown into a 10-ml droplet of hypotonic graphed using phase-contrast and fluorescence microscopy. Phase-contrast and corresponding CPD photographs (magni-bursting medium (0.05% v/v Nonidet P-40 and 0.1% w/v
bovine serum albumin) suspended from the end of a 200-ml fication of prints32258) were scanned at a resolution of 300 dpi into a computer. The chromosome-measuring program pipette tip. The resulting droplet was immediately placed in
the center of a glow-discharged (hydrophilic) microscope Micromeasure 3.01 (available at http://www.colostate.edu/ Depts/Biology/Micromeasure/) was used to determine relative slide, and an additional 10ml of hypotonic bursting medium
was added. A hand-held nebulizer (Fullam, Latham, NY) was SC lengths and arm ratios from phase-contrast images. The SC karyotype ofShermanandStack(1992, 1995) was used used to immediately give the slide 30 puffs of 4% aqueous
formaldehyde (pH 8.5). The slide was air-dried, fixed in 4% to identify individual-SCs. Adobe Photoshop 4.0 was used to digitally merge phase-contrast and corresponding CPD im-aqueous formaldehyde for 10 min, rinsed twice without
agita-tion in aqueous 0.01% Photoflo 200 (Kodak, Rochester, NY), ages. To describe the position of each CPD band, the distance along a CPD-banded SC from the center of the CPD band to rinsed four times (20 sec each rinse) in distilled water, and
air-dried. the center of the kinetochore was measured and divided by the length of the entire SC arm. In this way, the position of
Fluorescencein situhybridization:FISH was performed
us-ing a combination of the protocols of Rayburn and Gill each CPD band was expressed as a decimal fraction of the arm length from the centromere. These fractions were (1985) andAnamthawat-Jo´ nssonet al. (1996) with
signifi-cant modifications. Slides with spread SCs were incubated in multiplied by the mean length of the SC arm (Shermanand
Stack1995) to give micrometer distances from bands to the
23SSC containing 100mg/ml RNase A (Sigma, St. Louis) at
378for 30 min, washed in two changes of 23 SSC (378, 10 centromere (Table 1). Likewise, the length of each CPD band was measured and divided by the length of its entire SC to min each wash), rapidly dehydrated in a graded ethanol series,
and air-dried. Chromosomal DNA was denatured by placing determine the relative proportion of chromosome length con-tained within the band (Table 1).
slides in 50 ml of 70% formamide in 23 SSC at 708for 2.5
min. Slides were rapidly dehydrated in an ethanol series at FISH data analysis:For each late pachytene SC spread (i.e., spreads with visible kinetochores) showing a FISH signal, a 2208 and air-dried. A cocktail containing 75 ml deionized
formamide, 30ml 50% w/v potassium dextran sulfate, 15ml phase-contrast and a PI/FITC photograph were taken. For most sets, a CPD image was also obtained. Phase-contrast and 203SSC, 7.5 ml of 10 mg/ml sheared herring sperm DNA
(GIBCO BRL), and 12ml of 25mg/ml biotin-labeled probe(s) FITC/PI photographs were scanned and digitally merged to allow accurate measurement of distances between FISH signals was heated at 978 for 10 min. Thirty microliters of the
heat-denatured cocktail was placed on a slide, and a 22-350-mm and kinetochores. In all instances, SC 11 was identified on the basis of relative length and arm ratio according to the SC coverglass was added. Slides were incubated in sealed humid
chambers at 378overnight. Coverglasses were gently washed karyotype data of Sherman and Stack (1992, 1995). In spreads where FISH labeling was observed and a correspond-from slides with 23 SSC at 378. Slides were washed twice in
23 SSC, once in 25% formamide in 23 SSC, twice in 23 ing CPD image was available, the pattern of CPD staining was used to confirm the identity of SC 11 (see results). SSC containing 2% Tween-20 (all washes 378, 10 min), and
incubated for 60 min at 378in blocking buffer (0.1mmaleic To determine the distance between an FITC focus and the kinetochore, the distance along the SC from the location of acid, 0.15mNaCl, 1% w/v Boehringer Mannheim blocking
reagent, pH 7.5). Slides were incubated for 1 hr at 378in 1mg/ the focus to the center of the kinetochore was measured and divided by the length of the entire SC arm to express the ml mouse anti-biotin (Boehringer Mannheim) in blocking
buffer, washed three times in 23SSC containing 2% Tween- focus location as a decimal fraction of the arm length from the centromere. Fraction distances were multiplied by the 20 (378, 10 min each wash), incubated for 1 hr at 378 in
10mg/ml biotinylated goat anti-mouse (Sigma) in blocking mean length of the SC arm to give micrometer distances from foci to the centromere. This is the same technique used by buffer, and washed three times in 23 SSC containing 2%
Tween-20 (378, 10 min each wash). Slides were then incubated ShermanandStack(1995) to describe the location of recom-bination nodules on tomato SCs. In cases where an FITC focus in 40 mg/ml mouse anti-biotin fluorescein isothiocyanate
(FITC; Sigma) or 4 mg/ml streptavidin-FITC conjugate in did not lie directly on the SC, a line was drawn from the center of the FITC focus perpendicular to the axis of the SC. The blocking buffer for 1 h (378). All slides were washed once in
23 SSC containing 2% Tween-20 (378, 10 min) and twice point of intersection between the line and the SC axis was marked as the location of that focus on the SC. In some in 23SSC at room temperature (10 min each wash).
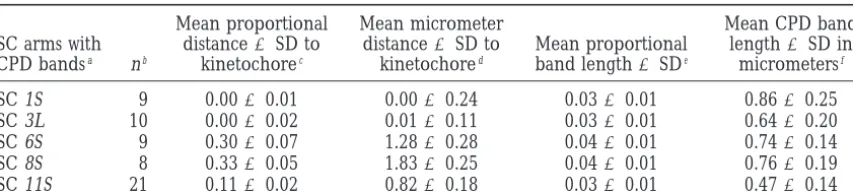
TABLE 1
Karyotype of the CPD-banded tomato SC complement
Mean proportional Mean micrometer Mean CPD band SC arms with distance6SD to distance6SD to Mean proportional length6SD in CPD bandsa nb kinetochorec kinetochored band length6SDe micrometersf
SC 1S 9 0.0060.01 0.0060.24 0.0360.01 0.8660.25 SC 3L 10 0.0060.02 0.0160.11 0.0360.01 0.6460.20 SC 6S 9 0.3060.07 1.2860.28 0.0460.01 0.7460.14 SC 8S 8 0.3360.05 1.8360.25 0.0460.01 0.7660.19 SC 11S 21 0.1160.02 0.8260.18 0.0360.01 0.4760.14
aS, short arm; L, long arm.
bn, number of SC spreads examined in which the CPD-banded SC could be positively identified on the basis
of its relative length and arm ratio.
cMean distance from the center of a specific CPD band to the center of the kinetochore expressed as a
proportion of the length of the SC arm in which the band is found. The center of the kinetochore has a value of 0 while the end of any particular arm has a value of 1.0.
dFor a particular CPD band, the proportional distance from the kinetochore multiplied by the mean length
of the SC arm in which the band is found gives a mean distance (mm) from the band to the kinetochore. The mean arm lengths (fromShermanandStack1995) are as follows: 1S, 7.5mm; 3L, 17.9mm; 6S, 4.3mm; 8S, 5.5mm; and 11S, 7.3mm.
eMean proportional length of the entire SC occupied by the band.
fFor a particular CPD band, the proportional length of a CPD band multiplied by the mean length of the
SC on which it is found. Mean SC lengths in micrometers (fromShermanandStack1995) are as follows: 1, 30; 3, 23.1; 6, 18.5; 8, 18.5; and 11, 16.2.
the end of the SC, a perpendicular line was drawn from the which suggests that kinetochores are highly infiltrated focus through the extended line, and the point of intersection with chromosomal DNA.
was marked as the location of the focus.
In PI-stained early pachytene SC spreads of tomato, Initially, all measurements were made from SC spreads
la-heterochromatic regions often appear wider than eu-beled with only one of the three probes. However, after it
chromatic regions (Figure 1B). Additionally, hetero-was clearly established that there hetero-was a considerable physical
distance betweenlTG523 andlTG46 and that there was no chromatin stains a bit more intensely than euchromatin. difficulty determining which signal belonged to which probe, In late pachytene spreads, PI-stained SCs have a rather measurements were made from these dual-labeled spreads as homogenous appearance with no noticeable staining well. No measurements were made from spreads probed with
difference between heterochromatin and euchromatin. any other combination of markers. Statistical analysis of the
Kinetochores stain with an intensity similar to that of distribution of FITC foci for the three probes was performed
using the computer program Instat 1.12a (GraphPad Software, nearby chromatin, again indicating the presence of
San Diego). DNA in kinetochores.
Dot blot verification of probe identity: To rule out the When early pachytene tomato SC spreads are stained unlikely possibility that the genomic probes used for FISH
with CPD and examined using a wide UV filter, euchro-were inadvertently switched or cross-contaminated before in
matin and heterochromatin are easily differentiated situ hybridization, each DIG-labeled RFLP marker was used
to probe a separate nylon membrane on which a 20-ml drop with heterochromatin fluorescing white and euchroma-(0.5 mg) of each of the biotin-labeled l insert DNAs had tin fluorescing blue (Figure 1C). The differential stain-been dotted. Filter hybridization conditions and colorimetric ing of heterochromatin and euchromatin seen after detection of probe hybridization were performed as described
CPD treatment is more striking than differences pro-above.
duced by PI staining alone. By late pachynema, hetero-chromatin and euhetero-chromatin do not exhibit a predict-able pattern of differential staining (Figure 1D). RESULTS
Throughout pachynema, the NOR (which comprises
DAPI, PI, and CPD staining of tomato SCs: When most of the short arm of chromosome 2) is easily identi-tomato SC spreads are stained with DAPI, pericentro- fied by its red fluorescence (Figure 1, C and D). As meric heterochromatin, euchromatin, and telomeres previously described, the NOR typically exhibits partial are not readily differentiated. However, some bivalents asynapsis (Shermanand Stack 1992; Peterson et al.
possess a single, highly localized region that does not 1996), and, as pachynema progresses, the NOR becomes fluoresce, i.e., a DAPI-negative band (Figure 1A). DAPI- more elongate (Figure 1D).
Figure1.—Staining of chro-matin associated with tomato SC spreads. (A) Early pachy-tene SC set stained with DAPI only. Several SCs possess a re-gion where DAPI staining is not visible, i.e., a DAPI-negative band (arrows). No other chromatin-based features are visible. (B) Early pachytene SC spread stained with PI only. Hetero-chromatic regions (e.g., arrow-heads) appear wider and stain a bit more intensely than the distal euchromatic regions (e.g., arrows). (C) CPD staining of an early pachytene SC spread. Pericentromeric heterochro-matin fluoresces white while distal euchromatin fluoresces blue. A red-fluorescing chro-matin band (CPD band) is visi-ble within the pericentromeric heterochromatin of 5 of toma-to’s 12 SCs (arrows). These bands most likely correspond to the DAPI-negative bands seen in DAPI-stained tomato SC spreads. The NOR near the terminus of the short arm of SC 2 fluoresces red and usually exhibits partial or complete asynapsis (e.g., diamond-head arrows). (D) Late pachytene CPD-stained SC spread. Kinet-ochores are visible as thick-enings along SCs (e.g., white boxes). On two CPD-banded SCs, the kinetochores are lo-cated directly over the CPD band (arrowheads). However, on the remaining three CPD-banded SCs a red band is visi-ble outside of the region en-compassed by the kinetochore (arrows). The elongate, asyn-apsed NOR fluoresces red (dia-mond-head arrows). (E) Phase-contrast image of a complete spread of late pachytene to-mato SCs. A kinetochore is visi-ble on each SC (black arrows). One SC is broken into two pieces (white arrow). Likewise, the NOR region has been lost from the end of SC 2 (white arrowhead). (F) CPD staining of the same SC spread shown in E. CPD bands are visible on five of the SCs (arrows). (G) Diagram showing the relative position of CPD bands and kinetochores. Blue lines represent SCs, yellow dots represent kinetochores, and red bars represent CPD bands. The identity (chromosome number) of each CPD-banded chromosome is shown next to an arrow pointing to its kinetochore. (H) Idiogram of the tomato SC complement showing the distribution of CPD bands. SCs 1-12 are shown in consecutive order from left to right. Euchromatin is blue, heterochromatin is white, kinetochores are orange, and CPD bands are red. The NOR in the short arm of SC 2, which appears red after CPD staining, is colored pink in this idiogram. A constricted area within the NOR marks the general region where SC formation does not occur. Frames that share a common bar: A and B; C and D; and E–G. Bars, 10mm.
location of CPD bands suggest that they are equivalent and Stack (1992), to construct a CPD karyotype for tomato. CPD bands are found on SCs 1, 3, 6, 8, and 11 to the DAPI-negative bands seen in tomato SC spreads
stained with DAPI only (Figure 1A). We used CPD stain- (Figure 1, E–G and Table 1). The CPD bands on SCs
1 and 3 lie essentially at centromeric positions while the
bands on SCs 6, 8, and 11 are located at subcentromeric from several experiments, some experiments involved use of only one probe at a time, and probes used in all positions in the short arms of their respective
chromo-somes (Figure 1G). An idiogram showing the distribu- FISH experiments were removed from the same three tubes from which samples were taken for dot blot probe tion of CPD bands on the tomato SC complement is
shown in Figure 1H. verification (seematerials and methods).
SCsvs.“prefixed” pachytene chromosomes:In initial
Fluorescencein situhybridization:Probes containing
single-/low-copy sequences were obtained by screening attempts to localize specific DNA sequences on meiotic chromosomes, we used pachytene chromosomes fixed a tomato l genomic library with the chromosome
11-associated RFLP markers TG46, TG400, and TG523 before spreading in 1:3 acetic ethanol, i.e., “prefixed pachytene chromosomes,” as substrates for FISH rather (Tanksleyet al. 1992). For each RFLP marker, a
corre-sponding positive genomic clone was selected, i.e., than SC spreads. Although, when repetitive sequence probes were used, prefixed pachytene chromosomes
lTG46,lTG400, andlTG523. Tomato insert DNA from
each clone was isolated (lengths of tomato inserts: were adequate substrates for FISH (e.g., Peterson et al. 1998), localization of single-copy probes on these lTG46, 16,550 bp;lTG400, 10,200 bp;lTG523, 14,000
bp) and labeled with biotin using nick translation. La- chromosomes proved difficult because of high levels of background fluorescence and a scarcity of chromosome beled insert DNA was hybridized to spreads of tomato
SCs on microscope slides. Slides were treated with sets in which each bivalent could be distinguished, i.e., less than one completely interpretable set per slide (data mouse anti-biotin followed by incubation in biotinylated
goat anti-mouse. Sites of probe hybridization were de- not shown). When SC spreads were substituted for pre-fixed pachytene chromosomes, significant problems tected using anti-biotin-FITC or streptavidin-FITC
con-jugates. SC 11 was identified on the basis of its relative with background fluorescence were eliminated and chromosome identification was facilitated because each length and arm ratio. Additionally, CPD banding
analy-sis was used to confirm the identity of SC 11. The Micro- slide possessed 10 or more SC sets where all 12 tomato SCs were clearly identifiable (e.g., Figure 1, E–G). measure chromosome measuring program was used to
determine the distance from the centromere to each FITC focus.
DISCUSSION Each of the three genomic probes hybridized
exclu-sively to euchromatin on the long arm of SC 11 (Figure CPD staining:If tomato SC spreads are stained with a combination of PI and DAPI, i.e., CPD stained, and 2). Little or no background FITC fluorescence was
ob-served, and the efficiency of hybridization (i.e., the per- illuminated with broad-band UV-visible light, structural features associated with differential chromatin conden-centage of SC spreads where specific labeling of SC 11
could be seen) was high for all three probes (ca. 70%). sation and/or DNA sequence can be visualized. These features either are not visible by DAPI or PI staining On the basis of the relative locations of FITC foci, the
three probes were positioned in relationship to the cen- alone or are more readily differentiated by CPD stain-ing. Although a variety of dye concentrations were tromere and to one another (Table 2 and Figure 2,
N–P). We refer to the diagram showing the position of tested, best results were obtained when chromosomes were stained with 1mg/ml PI and 0.5mg/ml DAPI (data each locus on SC 11 as an SC-FISH map (Figure 2O).
One-way analysis of variance (ANOVA) indicates that not shown).
Euchromatin and heterochromatin can be differenti-the difference of differenti-the group means for differenti-the probes is
highly significant (F5146.9, P,0.0001). Comparison ated by CPD staining in early pachytene SC spreads, but by late pachynema, the two types of chromatin are not of the data for any two individual probes also indicates
that the loci are clearly separated (Bonferroni P value readily distinguished. The explanation for this may be that the difference in relative condensation of hetero-for any two probes is,0.001).lTG523 is the most distal,
i.e., farthest from the centromere, of the loci with a near chromatin and euchromatin is decreased during the transition from early to late pachynema. On the other terminal location. The probeslTG46 andlTG400 are
located more proximally on SC 11 withlTG400 being hand, CPD bands and NORs remain differentially stained throughout pachynema (and probably through-closer to the centromere thanlTG46. This contradicts
the molecular linkage map of chromosome 11 on which out meiosis).
Likewise, a combination of PI and DAPI has been TG400 is located between TG523 and TG46 (Tanksley
et al. 1992; compare Figure 2N and 2O with 2P). To shown to differentially stain NORs in mitotic and mei-otic metaphase chromosomes of cotton (Hansonet al.
rule out the possibility that the probes lTG46 and
lTG400 had been inadvertently switched before FISH, 1995; Ji et al. 1997), mitotic metaphase chromosomes
of tomato (T. P. V. Hartman, personal communica-we performed a series of dot blot experiments. These
experiments verified that each biotin-labeled FISH tion), and SC spreads from maize and potato (our un-published observations). Consequently, it seems likely probe hybridized exclusively with the RFLP marker
orig-inally used to isolate it. It is unlikely that probes were that the NORs of many, if not all, plants can be visualized using CPD staining.
Figure2.—Single-/low-copy FISH to tomato SC 11. (A) Phase-contrast image of a late pachytene SC 11. The kinetochore is clearly visible (arrow-head). (B) Hybridization oflTG523 to the long arm of the same SC. Probe hybridization is visible as yellow-green (FITC) foci (arrow). The chromo-some appears red because of the PI counterstain. The kinetochore is marked by an arrowhead. (C) CPD image of the same SC. As predicted by the CPD karyotype, this chromosome possesses a red CPD band (arrow) in the short arm near the kinetochore (arrowhead). (D) Phase-contrast im-age of SC 11. The kinetochore is indi-cated by an arrowhead. (E)lTG523 hybridization on the SC shown in D. The signal is localized near the end of the chromosome (arrow). Note that there is little background FITC fluo-rescence, and there are no ectopic sites of hybridization. (F and G) Phase-contrast and PI/FITC image of a chromosome 11 showing the kinet-ochore (black arrowhead) and lTG400 hybridization (white arrow), respectively. (H and I) Phase-contrast and PI/FITC images of the same chromosome 11. In I, simultaneous detection of lTG523 (arrowhead) andlTG46 (arrow) is shown. ( J–M) Centromere placement and probe positioning on an early pachytene to-mato SC 11. ( J) Phase-contrast image of an SC 11. No kinetochore is visible. (K) lTG523 hybridization (arrow). (L) CPD staining. Heterochromatin appears white and euchromatin blue. A distinct CPD band (arrowhead) is observed within the pericentromeric heterochromatin of the bivalent. (M) Diagram of chromatin features and hybridization sites superimposed on the phase-contrast image of the SC. In the diagram, euchromatin is blue, heterochromatin is green, the CPD band is red, and FITC foci are yellow. Although a kinetochore is not visi-bleon this SC, the consistent relation-ship between the location of CPD bands and kinetochores allows accurate placement of a “kinetochore” on the SC (black band;
seediscussion). (N–P) SC-FISH and molecular linkage maps of tomato chromosome 11. In all diagrams,lTG400,lTG46, and
TABLE 2
Locations of three probes on the long arm of tomato SC11
Number of Mean locus position6SD Distance (mm) from Probes na observed FITC foci from kinetochoreb kinetochorec6SD
lTG46 16 38 0.7660.03 6.7860.28
lTG400 12 31 0.6260.04 5.5460.37
lTG523 21 58 0.9760.04 8.6160.37
aNumber of SC sets from which data were collected for a particular probe. In all sets from which measurements
were made, SC 11 could be positively identified and FISH label was observed only on this SC.
bMean locus position is expressed as a decimal fraction of the long arm from the kinetochore. The center
of the kinetochore has a decimal fraction value of 0, the terminus of the long arm has a value of 1, and foci distal to the terminus have values.1.
cThese values were obtained by multiplying the average locus position by 8.9mm, which is the mean length
of the long arm of SC 11 (ShermanandStack1995).
Insight into the probable mechanisms by which CPD CPD bands may be equivalent to certain C-bands. A C-band karyotype based on 1:3 acetic ethanol-fixed staining differentially stains NORs and CPD bands may
be gained by considering the mechanisms by which PI tomato (Moneymaker) pachytene chromosomes shows prominent centromeric/subcentromeric C-bands on and DAPI interact with DNA. PI intercalates between
bases of single- or double-stranded nucleic acid mole- chromosomes 1, 4, 6, 8, and 11 and staining of the NOR on chromosome 2 (M. S.Ramannaand L. P.Pijnacker, cules without regard to nitrogenous base composition,
and consequently PI can be used as a quantitative nu- unpublished data). The relative location of major C-bands on chromosomes 1, 6, 8, and 11 corresponds cleic acid dye (Heslop-HarrisonandSchwarzacher
1996). DAPI is a double-stranded DNA-specific dye that exactly with locations of CPD bands on these chromo-somes. However, the CPD- and C-band karyotypes differ interacts with DNA by at least two different mechanisms
(see Kapuscinski 1995 for review). In regions where in three ways: (1) There is no corresponding CPD band for the major C-band near the centromere of chromo-three or four AT base pairs are located in tandem, DAPI
binds to the minor groove of the DNA, and this results in some 4. (2) A C-band is found near the centromere of chromosome 3, but it is very small and probably does a highly fluorescent compound. DAPI also intercalates
between bases, which results in a nonfluorescent com- not account for the relatively large CPD-band we observe on this chromosome. And (3) there are several small pound. The former reaction is energetically favored in
AT-rich regions while the latter is favored in GC-rich C-bands for which corresponding CPD bands were not observed. These discrepancies may be caused by differ-areas along a DNA molecule. As an intercalator, the
binding strength of DAPI (in GC-rich areas) is roughly ences in the heterochromatin of the cultivars Cherry and Moneymaker. Also the mechanism of C-banding equivalent to that of PI. NORs, i.e., rDNA sequences,
are GC-rich compared to most other DNA regions (Mac- (Sumner1990) differs from the proposed mechanism of CPD staining, so it is not surprising that some CPD
gregor and Kezer 1971; Yasmineh and Yunis 1971;
Ingleet al. 1975). As a result, in CPD-stained spreads, bands coincide with C-bands while others do not (and
vice versa).
minor groove binding of DAPI to rDNA sequences
would be limited. Presumably, DAPI and PI would com- SCs as substrates for single-copyin situhybridization:
Although FISH has been used to localize repetitive se-pete for intercalation sites in the GC-rich rDNA, but
only PI intercalation would result in fluorescence. In quences on spreads of vertebrate SCs (Moens and
Pearlman 1989, 1990; Heng et al. 1994; Solari and addition, the PI concentration in the CPD staining
solu-tion is double that of the DAPI concentrasolu-tion, and thus Dresser1995) and on SC spreads of two plant species (Hasenkampf1991;AlbiniandSchwarzacher1992), PI intercalation would be favored. Consequently, red
(PI) fluorescence predominates in NORs. Similarly, to our knowledge, there have been no reports of single-copy FISH to SCs of any species. Here we demonstrate DAPI-negative bands (seen in DAPI-stained SC spreads)
and CPD bands probably represent other chromosomal that spreads of tomato SCs are well suited for the rigor-ous requirements of in situ detection of single-copy se-regions rich in GC base pairs. The presence of GC-rich
regions in heterochromatin (including GC-rich satellite quences. Because comparable SC spreads can be pre-pared for a variety of plants as well as animals and fungi, sequences) has been reported for other species (see
Sumner1990 for review). Differential staining between it is likely that this technique can be generally applied. In our experience, background fluorescence is mark-heterochromatin and euchromatin observed in early
pect that this phenomenon reflects the time at which matin, tomato should be particularly well suited for gene tagging and chromosome walking (Petersonet al. 1998).
chromosomes are fixed. Treatment of cells with
acetic-ethanol results in rapid protein crosslinking and fixa- Single-vs.low-copy number sequences:Molecular evi-dence indicates that TG523 and TG46 are located at tion of chromosome structure (Sharma and Sharma
1980). Consequently, in prefixed preparations nucleo- single loci on tomato chromosome 11 (Tanksleyet al.
1992; SolGenes web site, http://geneous.cit.cornell.edu/ plasm may be fixed onto chromosomes before squashes/
spreads are made. During FISH, probes and fluoro- solgenes/aboutsolgenes.html/). Our observation that
lTG523 andlTG46 each bind exclusively to a single, chrome-labeled affinity reagents may become trapped
within nucleoplasm surrounding the chromosomes to highly localized region on SC 11 supports this premise. On the other hand, blotting evidence suggests that produce high background fluorescence. While
diges-tion of chromosomes with a protease(s) before FISH can TG400 may occur at more than one locus in the tomato genome (S. D.Tanksley, personal communication; Sol-be used to remove at least some of the contaminating
nucleoplasm from prefixed pachytene chromosomes Genes web site). However, we observed only one highly localized site oflTG400 probe hybridization on tomato (Jianget al. 1995), protease treatment also results in
significant changes in chromosome structure (our expe- SC spreads, and this hybridization site is on the long arm of SC 11. If there is indeed more than one copy rience). In contrast, SCs are spread before fixation.
Dur-ing the spreadDur-ing process, nucleoplasm around SCs is of TG400 in the tomato genome, this result might be explained if the copies of TG400 are so closely associated dispersed and chromatin is decondensed, which allows
better access of single-copy probes to chromosomal DNA. that they are not resolvable using our FISH method. Alternatively, it is possible that thelTG400 FISH clone Although we believe that SC spreads possess some
features that make them better suited for single-copy hybridized to a TG400 site on SC 11 and did not recog-nize TG400 loci elsewhere.
FISH than prefixed pachytene chromosomes, it should
be noted that other investigators have successfully local- Comparison of the SC-FISH and molecular genetic maps: While linkage maps show the relative order of ized single-copy sequences on prefixed pachytene
chro-mosomes by in situ hybridization (Shenet al. 1987;Jiang genes along each chromosome, they provide little in-sight into the relationship between genes and
chromo-et al. 1995).
CPD banding and FISH: In all cases where single- some structure. This is evident in a comparison of the SC-FISH map (Figure 2O) with the molecular linkage copy FISH was detected and a CPD photograph was
taken, a CPD band was observed in a position predicted map of chromosome 11 (Figure 2P). On the molecular linkage map, TG523 is closer to the center of the map by the CPD band karyotype of SC 11. Although SC 11
can be readily identified on the basis of relative length than it is to the terminus, but on the SC-FISH map,
lTG523 is located near the end of the long arm of SC and arm ratio, FISH and CPD results each
indepen-dently indicate that SC 11 was correctly identified in 11. Because TG523 is not the most distal locus on the
molecular linkage map, it is likely that there are other this study.
Nature of the tomato genome:Because it is possible genes/markers clustered within the more distal subtelo-meric region of the chromosome. With respect to dis-that genomic clones isolated by screening a library with
RFLP markers might contain repetitive elements in ad- tances between markers, all three SC-FISH markers are contained within a chromosomal region comprising dition to single-copy DNA, genomic probes are often
“prehybridized” with unlabeled repetitive DNA se- only 19% of the length of SC 11. However, the three RFLP markers span a region encompassing 35% of the quences before they are placed on chromosomes. Such
chromosomal in situ suppression (CISS) hybridization molecular linkage map (Tanksleyet al. 1992).
On the SC-FISH map, the order of lTG46 and effectively prevents highly repetitive sequences in
probes from participating in hybridization with chromo- lTG400 is reversed in relation to their order on the molecular linkage map (Figure 2, N–P). This disagree-somal DNA (Landegentet al. 1987;Lichteret al. 1988,
1990;Jianget al. 1995, 1996;Zwicket al. 1997). How- ment is not caused by an inability to resolve these mark-ers cytologically, because (with the exception of a single ever, in earlier studies we determined that only 23% of
the tomato genome is composed of repetitive sequences FITC focus) the ranges of FITC foci for the markers do not overlap (Figure 2N). While somewhat unexpected, and that the vast majority of these repetitive sequences
are localized in pericentromeric heterochromatin (see disagreement between the order of loci on tomato maps is not without precedent (e.g.,Schumacheret al. 1995).
Peterson et al. 1996, 1998). Consequently, because
there was reason to believe that our genomic probes Relative order of TG46 and TG400:There are several possible explanations for the discrepancy between the might not contain repetitive sequences, we tried FISH
without CISS hybridization, and indeed, we found that relative positions of TG46 and TG400 on the SC-FISH and molecular genetic maps. For example, it is possible each of the three genomic probes hybridized exclusively
to a different highly localized region on SC 11 in the that (1) thelTG400 probe recognizes a locus on SC 11 different from the TG400 locus positioned on the link-absence of suppression sequences. With a relatively
incor-TABLE 3
Tomato DNA per micrometer of SC in euchromatin and heterochromatin
Proportion of DNA concentration
genome contenta DNA content (bp)b SC length (mm)c (bp/mm)
Euchromatin 0.23 1.913108 136.6 1.403106
Heterochromatin 0.77 6.393108 76.4 8.363106
aData fromPetersonet al. (1996).
bDetermined by multiplying the fraction of the genome occupied by each type of chromatin by the 1C
genome size (8.303108bp;Petersonet al. 1998) for tomato.
cData fromShermanandStack(1995).
rectly positioned on the molecular linkage map; or (3) of the SC-FISH markers and between the SC-FISH mark-ers themselves (Table 4). On the basis of the data in the order of loci in cv. Cherry tomato differs from the
gene order in the cultivar that was used to prepare the Table 4 and the average positions for the three probes shown in Figure 2O, we estimate that SC-FISH easily molecular linkage map (i.e., VF36-Tm2a; Tanksley et
al. 1992). permits resolution of loci separated by as few as 1.7
million bp with every indication that much closer loci The first of these possibilities seems unlikely because
the lTG400 probe would have to consistently fail to could be resolved. For comparison,Trask(1991) esti-mated that FISH probes can be mapped to human meta-hybridize to the “RFLP-mapped” copy of TG400 on
chro-mosome 11, yet succeed in hybridizing to a second copy phase chromosomes at a resolution ofz3 million bp. On the other hand, substrates other than condensed of TG400 on chromosome 11 overlooked during RFLP
mapping. The homology between the two TG400 copies chromosomes can be used to improve resolution. For example, FISH has been used to resolve single-copy (even if limited to the 1200 bp of the RFLP marker
itself ) would probably be sufficient to produce some sequences separated by as little as 50 kb in the decon-densed chromatin of interphase nuclei (Lawrence et
positive hybridization at both sites. Consequently, a less
defined distribution oflTG400 FITC foci encompassing al. 1988; Trask et al. 1991). Additionally,Heng et al.
(1992) resolved genes as close as 20 kb by FISH to free the range of lTG46 hybridization sites would be
ex-pected, but this is not observed (Figure 2N). chromatin, andFranszet al. (1996) resolved sequences
separated by ,1 kb using stretched, naked DNA as a In reference to the second possibility, construction
of linkage maps involves sexual crosses and sometimes FISH substrate. Unfortunately, with these latter tech-niques individual chromosomes and structural features difficult and/or complex scoring of recombination
events that could lead to a mistake. However, because such as heterochromatin, centromeres, and telomeres are not readily distinguishable.
TG46, TG400, and TG523 have LOD scores of at least
3.0 (Tanksleyet al. 1992), it is unlikely that the markers Consolidation of the SC-FISH and recombination nodule maps:Because of the discrepancy between the have been positioned incorrectly on the molecular
link-age map (seeTamarin1999). SC-FISH and RFLP maps for tomato chromosome 11, a meaningful discussion of the relationship between Perhaps the most likely explanation for the
discrep-ancy between the SC-FISH and RFLP map is a difference molecular map distances and the number of base pairs between the three SC-FISH marker loci is not possible. in locus order between cv. Cherry and cv. VF36-Tm2a
because of a small inversion in chromosome 11. In However, the SC-FISH and recombination nodule maps for SC 11 (ShermanandStack1995) can be compared crosses between our Cherry tomato line and
character-ized translocation lines of cv. Early Fire Ball, no inver- to determine map distances between SC-FISH markers. [Recombination nodules (RNs) are ellipsoidal struc-sion loops were observed in pachytene SC spreads from
the F1progeny (unpublished observations made during tures associated with SCs during late pachynema, and there is a 1:1 correlation between RNs and crossover research of Herickhoff et al. 1993). Consequently, if
cv. Cherry contains an inversion in chromosome 11 events (Carpenter 1975; Sherman and Stack 1995;
Andersonet al. 1997).] Comparison of the maps
indi-relative to cv. Early Fire Ball, either the inversion is
not large enough to produce an inversion loop in the cates that the rate of crossing over per megabase pair (106bp) is similar in the interval flanked bylTG400 and heterozygote or cv. Cherry and cv. Early Fire Ball have
the same inversion. lTG46 and the interval flanked bylTG46 andlTG523 (Table 5). Thus, at least at the resolution of this analysis,
Base pair distances between SC-FISH markers:Data
from previous investigations (Peterson et al. 1996, there is no obvious recombination hot/cold spot in one of these intervals compared to the other (seeSegalet
1998) were used to calculate DNA densities (1C) in
heterochromatic and euchromatic regions along to- al. 1992 for an example of a recombination hot spot in
tomato euchromatin). mato SCs (Table 3). This permitted an estimate of the
pachy-TABLE 4
Calculation of base pairs between different molecular and cytological markers
SC in euchromatin SC in heterochromatin Base pairs Markers between markers (mm)a between markers (mm)a between markersb
lTG46 and centromere 2.98 3.80 3.603107
lTG400 and centromere 1.74 3.80 3.423107
lTG523 and centromere 4.81 3.80 3.853107
lTG523 andlTG46 1.83 0 2.533106
lTG523 andlTG400 3.07 0 4.303106
lTG46 andlTG400 1.24 0 1.743106
aFromShermanandStack(1995), the long arm of SC 11 (8.9mm mean total length) has an average of
3.8 mm of its length in pericentromeric heterochromatin. To determine the length of the long arm in euchromatin between a probe and the centromere, 3.8 mm was subtracted from the distance between the probe and the centromere (see Table 2). Regions between probes are euchromatic.
bBase pairs per micrometer of SC in euchromatin and heterochromatin (as calculated in Table 3) are 1.403
106and 8.363106, respectively. The number of base pairs between two markers was calculated by multiplying the micrometers in euchromatin between the markers by 1.40 3 106 bp, multiplying the micrometers in heterochromatin between the two markers by 8.363106bp, and adding the resulting values together.
tene SCs:Because heterochromatin and euchromatin arm to the kinetochore, i.e., the length of the short arm, is [5.8mm 4 (1.02 0.112) 5] 6.5mm, and the long can be differentiated in early pachytene SCs, SC-FISH
to early spreads may be useful in determining the chro- arm is (15.0mm26.5mm5) 8.5mm in length. Conse-quently, in Figure 2M the centromere (purple band) matin background in which a gene/transgene is found.
However, it is difficult to position hybridization sites has been placed 6.5mm from the end of the short arm. Once the centromere has been positioned, the locations on early pachytene SCs because they do not possess
cytologically discernible centromeres/kinetochores at of FISH hybridization signals can be expressed as frac-tions of the length of the chromosome arm to the cen-the light microscopic level (Stack and Anderson
1986). This problem can be overcome for chromosomes tromere. For example, the twolTG523 FITC foci near the terminus of the long arm of SC 11 (Figure 2, K and with a characterized CPD band(s) because the band can
be used to position the centromere. An example of M) are 7.8 and 7.9mm from the centromere, respectively. Because the length of the long arm is 8.5mm (see above), centromere placement in relation to a CPD band is
shown in Figure 2, J–M. The SC 11 in these frames is the two foci lie at points [100 3 (7.8 mm 4 8.5 mm)
5] 92.0% and [100 3(7.9mm4 8.5mm) 5] 93.0% 15.0mm in length. The distance from the end of the
short arm (i.e., the arm lacking alTG523 FISH hybrid- of the long arm from the centromere.
The potential of single-copy FISH to SCs:In this study ization signal) to the CPD band is 5.8mm. From Table
1, the relative distance between the CPD band and the we demonstrated that single-copy sequences can be reli-ably detected on SC spreads, discovered a discrepancy kinetochore of SC 11 is 11.2% of the length of the short
arm. Thus the distance from the terminus of the short between our SC-FISH map and the molecular genetic
TABLE 5
Map distances between markers determined by integrating the data from the recombination nodule map of tomato with the SC-FISH data
Total number of RNs Map units Map units per 106bp of Markers between markersa between markersb DNA between markersc
lTG46 and centromere 219 25.9 0.7
lTG400 and centromere 112 13.3 0.4
lTG523 and centromere 363.5 43.1 1.1
lTG523 andlTG46 144.5 17.1 6.8
lTG523 andlTG400 251.5 29.8 6.9
lTG46 andlTG400 107 12.7 7.3
aThe mean number of RNs observed between two markers based on an analysis of 422 SC 11s (Table A1
inShermanandStack1995).
bThe map units (percentage crossing over) between two markers were determined by dividing the total
number of observed RNs in the interval by the total number of SC 11s from which data were obtained (i.e., 422) and multiplying the quotient by 50 map units (seeShermanandStack1995 for more information).
heterologous DNA inserts on the mouse meiotic chromosome
map, estimated the distance between molecular markers
core. Chromosoma 103: 401–407.
in base pairs, and integrated the SC-FISH map with the Herickhoff, L., S. StackandJ. Sherman, 1993 The relationship
between synapsis, recombination nodules and chiasmata in
to-preexisting RN map for tomato. Additionally, our data
mato translocation heterozygotes. Heredity 71: 373–385.
provide further evidence that there are few repeated
Heslop-Harrison, J. S., andT. Schwarzacher, 1996 Flow
cytome-sequences in tomato euchromatin. In broader terms, try and chromosome sorting, pp. 85–106 in Plant Chromosomes— Laboratory Methods, edited byK. FukuiandS. Nakayama. CRC
SC-FISH mapping has considerable potential as a means
Press, Boca Raton, FL.
of relating genes to chromosome morphology
(espe-Ingle, J., J. N. TimmisandJ. Sinclair, 1975 The relationship
be-cially when coupled with differential chromatin-staining tween satellite deoxyribonucleic acid, ribosomal ribonucleic acid
gene redundancy, and genome size in plants. Plant Physiol. 55:
techniques such as CPD staining), studying the
relation-496–501.
ship between chromatin organization and gene
expres-Ji, Y., D. A. Raska, T. D. McKnight, M. N. Islam-Faridi, C. F. Crane sion, relating linkage distances to chromosome struc- et al. 1997 Use of meiotic FISH for identification of a new
monosome in Gossypium hirsutum L. Genome 40: 34–40.
ture, and locating sites of transgene insertions. Because
Jiang, J., B. S. Gill, G.-L. Wang, P. C. RonaldandD. C. Ward,
SC spreads have been prepared for many plants,
ani-1995 Metaphase and interphase fluorescence in situ
hybridiza-mals, and a few fungi, it is probable that SC-FISH map- tion mapping of the rice genome with bacterial artificial
chromo-somes. Proc. Natl. Acad. Sci. USA 92: 4487–4491.
ping can be used to investigate the genomes of a wide
Jiang, J., S. H. Hulbert, B. S. GillandD. C. Ward, 1996 Interphase
variety of organisms.
fluorescence in situ hybridization mapping: a physical mapping strategy for plant species with large complex genomes. Mol. Gen. This research was funded in part by the United States Department
Genet. 252: 497–502. of Agriculture grant 95-37300-1570 to S.M.S. and N.L.V. and Colorado
Kapuscinski, J., 1995 DAPI: a DNA-specific fluorescent probe. Bio-Agricultural Experiment Station grant 1878-670 to S.M.S.
technic Histochem. 70: 220–233.
Khush, G. S., andC. M. Rick, 1968 Cytogenetic analysis of the tomato genome by means of induced deficiencies. Chromosoma
23:452–484.
LITERATURE CITED Kota, R. S., K. S. GillandB. S. Gill, 1993 A cytogenetically based physical map of chromosome 1B in common wheat. Genome 36:
Albini, S. M.,andT. Schwarzacher, 1992 In situ localization of
467–475.
two repetitive DNA sequences to surface-spread pachytene chro- Landegent, J. E., N. Jansen In De Wal, R. W. Dirks, F. Bassand mosomes of rye. Genome 35: 551–559.
M. Van Der Ploeg, 1987 Use of whole cosmid cloned genomic
Anamthawat-Jo´ nsson, K., J. S. Heslop-HarrisonandT. Schwarz- sequences for chromosomal localization by non-radioactive in acher, 1996 Genomic in situ hybridization for whole chromo- situ hybridization. Hum. Genet. 77: 366–370.
some and genome analysis, pp. 1–23 in In Situ Hybridization, edited Lawrence, J. B., C. A. VillnaveandR. H. Singer, 1988 Sensitive, byM. Clark. Chapman and Hall, New York. high resolution chromatin and chromosome mapping in situ:
Anderson, L. K., H. H. Offenberg, W. M. H. C. Verkuijlenand Presence and orientation of two closely integrated copies of EBV
C. Heyting, 1997 RecA-like proteins are components of early in a lymphoma line. Cell 52: 51–61.
meiotic nodules in lily. Proc. Natl. Acad. Sci. USA 94: 6868–6873. Lehfer, H., W. Busch, R. MartinandR. G. Herrmann, 1993
Local-Carpenter, A. T. C.1975 Electron microscopy of meiosis in Drosoph- ization of the B-hordein locus on barley chromosomes using
ila melanogaster females. II. The recombination nodule—a recom- fluorescence in situ hybridization. Chromosoma 102: 428–432.
bination-associated structure at pachytene? Proc. Natl. Acad. Sci. Leitch, I. J., andJ. S. Heslop-Harrison, 1993 Physical mapping USA 72: 3186–3189. of four sites of 5S rDNA sequences and one site of thea
-amy-Curtis, C. A., andA. J. Lukaszewski, 1991 Genetic linkage between lase-2 gene in barley (Hordeum vulgare). Genome 36: 517–523. C-bands and storage protein genes in chromosome 1B of tetra- Lichter, P., T. Cremer, J. Borden, L. ManuelidisandD. C. Ward, ploid wheat. Theor. Appl. Genet. 81: 245–252. 1988 Delineation of individual human chromosomes in
meta-Dooner, H. K.1986 Genetic fine structure of the Bronze locus in phase and interphase cells by in situ suppression hybridization maize. Genetics 113: 1021–1036. using recombinant DNA libraries. Hum. Genet. 80: 224–234.
Dubochet, J., M. Ducommun, M. ZollingerandE. Kellenberger, Lichter, P., C. J. Chang, K. Call, G. Hermanson, G. A. Evanset 1971 A new preparation method for dark-field electron micros- al. 1990 High-resolution mapping of human chromosome 11 copy of biomacromolecules. J. Ultrastructure Res. 35: 147–167. by in situ hybridization with cosmid clones. Science 247: 64–69.
Flavell, R., D. SmithandW. Thompson, 1985 The distribution Lichter, P., A. L. Boyle, T. CremerandD. C. Ward, 1991 Analysis of recombination sites, the structure of ribosomal DNA loci and of genes and chromosomes by nonisotopic in situ hybridization. the multiplicity of sequences containing inverted repeats, pp. Genet. Anal. Tech. Appl. 8: 24–35.
1–14 in Molecular Form and Function of the Plant Genome, edited by Lohe, A. R., andA. J. Hilliker, 1995 Return of the H-word
(hetero-L. van Vloten-Doting, G. Grootand T. C. Hall. Plenum chromatin). Curr. Opin. Genet. Dev. 5: 746–755.
Press, New York. Macgregor, H. C., andJ. Kezer, 1971 The chromosomal
localiza-Fransz, P. F., C. Alonso-Blanco, T. B. Liharska, A. J. M. Peeters, tion of a heavy satellite DNA in the testis of Plethodon c. cinereus.
P. Zabelet al. 1996 High-resolution physical mapping in Arabi- Chromosoma 33: 167–182.
dopsis thaliana and tomato by fluorescence in situ hybridization Moens, P. B., andR. E. Pearlman, 1989 Satellite DNA I in chromatin to extended DNA fibres. Plant J. 9: 421–430. loops of rat pachytene chromosomes and in spermatids.
Chro-Go´ mez, M. I., M. N. Islam-Faridi, S.-S. Woo, K. F. Shertz, D. mosoma 98: 287–294.
Czeschin, Jr.et al. 1997 FISH of a maize sh2-selected BAC to Moens, P. B., andR. E. Pearlman, 1990 In situ DNA sequence
chromosomes of Sorghum bicolor. Genome 40: 475–478. mapping with surface-spread mouse pachytene chromosomes.
Hanson, R. E., M. S. Zwick, S. Choi, M. N. Islam-Faridi, T. Cytogenet. Cell Genet. 53: 219–220.
McKnight et al. 1995 Fluorescent in situ hybridization of a Moore, C., andF. Sherman, 1974 Lack of correspondence between bacterial artificial chromosome. Genome 38: 646–651. genetic and physical distances in the iso-1-cytochrome c gene of
Hasenkampf, C. A., 1991 In situ hybridization of Lilium whole mount yeast, pp. 295–306 in Mechanisms In Recombination, edited byR.
synaptonemal complex chromosomal preparations. Biotech. His- Grell. Plenum Press, New York.
tochem. 1: 210–215. Moses, M., 1968 Synaptinemal complex. Annu. Rev. Genet. 2: 363–
Heng, H. H. Q., J. Squire andL.-C. Tsui, 1992 High-resolution 412.
mapping of mammalian genes by in situ hybridization to free Paterson, A. H., 1996 Making genetic maps, pp. 23–39 in Genome chromatin. Proc. Natl. Acad. Sci. USA 89: 9509–9513. Mapping in Plants, edited byA. H. Paterson. Academic Press,
San Diego.
Pedersen, C., and I. Linde-Laursen, 1995 The relationship be- region relative to three marker loci on chromosome 1B of wheat (Triticum aestivum). Theor. Appl. Genet. 69: 263–270.
tween physical and genetic distances at the Hor1 and Hor2 loci
Solari, A. J., andM. E. Dresser, 1995 High-resolution cytological of barley estimated by two-colour fluorescent in situ hybridization.
localization of the XhoI and EcoRI repeat sequence in the pachy-Theor. Appl. Genet. 91: 941–946.
tene ZW bivalent of the chicken. Chromosome Res. 3: 87–93.
Pedersen, C., H. GieseandI. Linde-Laursen, 1995 Towards an
Stack, S. M., 1984 Heterochromatin, the synaptonemal complex, integration of the physical and the genetic chromosome maps
and crossing over. J. Cell Sci. 71: 159–176. of barley by in situ hybridization. Hereditas 123: 77–88.
Stack, S. M., andL. K. Anderson, 1986 Two-dimensional spreads
Peterson, D. G., H. J. Price, J. S. JohnstonandS. M. Stack, 1996
of synaptonemal complexes from solanaceous plants. II. Synapsis DNA content of heterochromatin and euchromatin in tomato
in Lycopersicon esculentum (tomato). Am. J. Bot. 73: 264–281. (Lycopersicon esculentum) pachytene chromosomes. Genome 39:
Stack, S. M., J. D. Sherman, L. K. Anderson, andL. S. Herickhoff, 77–82.
1993 Meiotic nodules in vascular plants, pp. 301–312 in
Chromo-Peterson, D. G., W. R. PearsonandS. M. Stack, 1998
Characteriza-somes Today, edited byA. T. SumnerandA. C. Chandley. Chap-tion of the tomato (Lycopersicon esculentum) genome using in vitro
man and Hall, New York. and in situ DNA reassociation. Genome 41: 346–356.
Sturtevant, A., andG. Beadle, 1939 An Introduction to Genetics. Ramanna, M. S., andR. Prakken, 1967 Structure of and homology
W. B. Saunders, Philadelphia. between pachytene and somatic metaphase chromosomes of
to-Sumner, A. T., 1990 Chromosome Banding. Unwin Hyman, London.
mato. Genetica 38: 115–133. Tamarin, R. H., 1999 Principles of Genetics. McGraw-Hill, Boston.
Rashtchian, A., andJ. Mackey, 1992 Efficient synthesis of biotinyl- Tanksley, S. D., N. D. Young, A. H. PatersonandM. W. Bonierbale, ated DNA probes using polymerase chain reaction. Focus 14: 1989 RFLP mapping in plant breeding: new tools for an old
64–65. science. BioTechnology 7: 257–264.
Rayburn, A. L., andB. S. Gill, 1985 Use of biotin-labeled probes Tanksley, S. D., M. W. Ganal, M. C. Prince, M. C. De Vicente, to map specific DNA sequences on wheat chromosomes. J. Hered. M. W. Bonierbaleet al., 1992 High density molecular linkage
76:78–81. maps of the tomato and potato genomes. Genetics 132: 1141–
Rick, C. M., 1991 Tomato paste: A concentrated review of genetic 1160.
highlights from the beginnings to the advent of molecular genet- Trask, B. J., 1991 Fluorescence in situ hybridization. Trends Genet.
ics. Genetics 128: 1–5. 7:149–154.
Sambrook, J., E. F. FritschandT. Maniatis, 1989 Molecular Clon- Trask, B. J., H. Massa, S. KenwrickandJ. Gitschier, 1991
Map-ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press, ping of human chromosome Xq28 by two-color fluorescence in Cold Spring Harbor, NY. situ hybridization of DNA sequences to interphase cell nuclei. Schumacher, K., M. GanalandK. Theres, 1995 Genetic and physi- Am. J. Hum. Genet. 48: 1–15.
Tsujimoto, H., andK. Noda, 1990 Deletion mapping by gametoci-cal mapping of the lateral suppressor (ls) locus in tomato. Mol.
dal genes in common wheat: position of speltoid suppression Gen. Genet. 246: 761–766.
(Q) and beta-amylase (beta-Amy-A2) genes on chromosome 5A.
Segal, G., M. Sarfatti, M. A. Schaffer, N. Ori, D. Zamiret al.
Genome 33: 850–853. 1992 Correlation of genetic and physical structure in the region
Wallrath, L. L., andS. C. R. Elgin, 1995 Position effect variegation surrounding the I2Fusarium oxysporum resistance locus in tomato.
in Drosophila is associated with an altered chromatin structure. Mol. Gen. Genet. 231: 179–185.
Genes Dev. 9: 1263–1277.
Sharma, A. K., andA. Sharma, 1980 Chromosome Techniques—Theory
Weith, A., andW. Traut, 1980 Synaptonemal complexes with
asso-and Practice. Butterworth & Company, London.
ciated chromatin in a moth, Ephestia kuehniella Z. Chromosoma
Shen, D., Z. WangandM. Wu, 1987 Gene mapping on maize
pachy-78:275–291. tene chromosomes by in situ hybridization. Chromosoma 95:
Wing, R. A., H.-B. Zhangand S. D. Tanksley, 1994 Map-based 311–314.
cloning in crop plants. Tomato as a model system: I. Genetic and
Sherman, J. D., andS. M. Stack, 1992 Two-dimensional spreads of
physical mapping of jointless. Mol. Gen. Genet. 242: 681–688. synaptonemal complexes from solanaceous plants. V. Tomato
Yasmineh, W. G., andJ. J. Yunis, 1971 Satellite DNA in calf hetero-(Lycopersicon esculentum) karyotype and idiogram. Genome 35:
chromatin. Exp. Cell Res. 64: 41–48. 354–359.
Zuckerkandl, E., andW. Hennig, 1995 Tracking heterochromatin.
Sherman, J. D., andS. M. Stack, 1995 Two-dimensional spreads Chromosoma 104: 75–83.
of synaptonemal complexes from solanaceous plants. VI. High- Zwick, M. S., R. E. Hanson, T. D. McKnight, M. N. Islam-Faridi, resolution recombination nodule map for tomato (Lycopersicon D. M. Stellyet al. 1997 A rapid procedure for the isolation of
esculentum). Genetics 141: 683–708. C
ot-l DNA from plants. Genome 40: 138–142. Snape, J. W., R. B. Flavell, M. O’Dell, W. G. HughesandP. I. Payne,