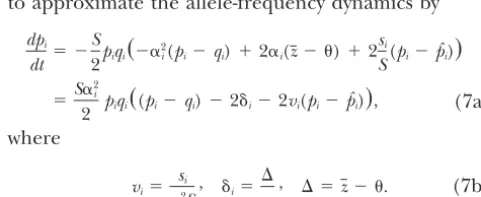Polygenic Variation Maintained by Balancing Selection: Pleiotropy,
Sex-Dependent Allelic Effects and
G
⫻
E
Interactions
Michael Turelli*
,1and N. H. Barton
†*Section of Evolution and Ecology and Center for Population Biology, University of California, Davis, California 95616 and
†Institute of Cell, Animal and Population Biology, University of Edinburgh, Edinburgh EH9 3JT, United Kingdom
Manuscript received May 29, 2003 Accepted for publication October 17, 2003
ABSTRACT
We investigate three alternative selection-based scenarios proposed to maintain polygenic variation:
pleiotropic balancing selection,G⫻E interactions (with spatial or temporal variation in allelic effects),
and sex-dependent allelic effects. Each analysis assumes an additive polygenic trait withndiallelic loci
under stabilizing selection. We allow loci to have different effects and consider equilibria at which the population mean departs from the stabilizing-selection optimum. Under weak selection, each model pro-duces essentially identical, approximate allele-frequency dynamics. Variation is maintained under pleio-tropic balancing selection only at loci for which the strength of balancing selection exceeds the effective strength of stabilizing selection. In addition, for all models, polymorphism requires that the population mean be close enough to the optimum that directional selection does not overwhelm balancing selection. This balance allows many simultaneously stable equilibria, and we explore their properties numerically.
Both spatial and temporalG ⫻ Ecan maintain variation at loci for which the coefficient of variation
(across environments) of the effect of a substitution exceeds a critical value greater than one. The criti-cal value depends on the correlation between substitution effects at different loci. For large positive
correlations (e.g.,2
ij⬎3/4), even extreme fluctuations in allelic effects cannot maintain variation.
Surpris-ingly, this constraint on correlations implies that sex-dependent allelic effects cannot maintain polygenic variation. We present numerical results that support our analytical approximations and discuss our results in connection to relevant data and alternative variance-maintaining mechanisms.
I
T remains a challenge for evolutionary geneticists to lelic effects (treating spatial and temporal heterogene-understand the additive genetic variance observed ity separately), and sex-dependent allelic effects. The for most traits in most populations. Given the ubiquity thread that unites these scenarios is that, under weak of additive genetic variation, it is natural to seek an selection, each produces very similar allele-frequency explanation in terms of ubiquitous forces.Lande(1975) dynamics and polymorphism conditions. An empirical proposed mutation-selection balance. However, over motivation for these analyses is that alleles of intermedi-the past 25 years, attempts to explain standing levels ate frequency seem to contribute to phenotypic varia-of quantitative genetic variation in terms varia-of mutation- tion in natural populations (e.g.,MackayandLangley selection balance have been at best only partially success- 1990;Longet al.2000). Such polymorphisms are incom-ful (e.g., Caballero and Keightley 1994; Charles- patible with mutation-selection balance for plausible worth and Hughes 2000; but see Zhang and Hill levels of selection and mutation.2002). One alternative is that some form of balancing The mathematical motivation for our analyses is selection, unconnected to the trait of interest, may ac- Wright’s (1935) demonstration that stabilizing selec-count for persistent polymorphism at the underlying tion tends to eliminate polygenic variation. Using a weak-loci (e.g., Robertson 1965; Bulmer 1973; Gillespie selection approximation, he showed that at most one 1984;Barton1990). In contrast to such pleiotropic ex- locus is expected to remain polymorphic at equilibrium. planations, balancing selection might arise from varia- More recent analyses of strong selection (Nagylaki tion in the effects of alleles that contribute to the trait, 1989; Bu¨ rgerandGimelfarb 1999) have found that for instance, through genotype-by-environment (G⫻E) two-locus polymorphisms can be stably maintained with interactions. Here we explore four scenarios in which sufficiently strong selection and sufficient interlocus variance-depleting stabilizing selection interacts with pleio- variation in allelic effects. We provide new simulations tropic balancing selection, environment-dependent al- that further illustrate the restrictive conditions needed to maintain even stable two-locus polymorphisms for ad-ditive traits under stabilizing selection and loose linkage. 1Corresponding author:Section of Evolution and Ecology, University
Robertson (1965) proposed that additive variation of California, 1 Shields Ave., Davis, CA 95616.
E-mail: [email protected] may be maintained by pleiotropically induced
inant selection, which counteracts the effects of stabiliz- that sex-dependent allelic effects do not stably maintain polygenic variation for additive traits.
ing selection. His conjecture was explored analytically
All of our analyses assume that selection is weak by Bulmer (1973) for diallelic loci and extended to
enough relative to recombination that linkage disequi-multiple alleles by Gillespie (1984). Both assumed
librium is negligible. We also assume diallelic loci. It is equal allelic effects across loci, symmetric
overdomi-not clear to us how restrictive this assumption is. In nance of equal intensity at all loci, and that the
popula-models of mutation-selection balance, two-allele and tion mean at equilibrium coincided with the optimal
continuum-of-allele models give similar results provided trait value. They found lower bounds on the intensity
that the alleles responsible for variation are rare ( Tur-of overdominant selection required to maintain stable
elli 1984;Slatkin andFrank1990). However, when multilocus polymorphisms. Our diallelic analyses
gener-loci are highly polymorphic (as might occur under bal-alize theirs and that of Zhivotovsky andGavrilets
ancing selection), continuum-of-allele models can give (1992), by allowing for unequal allelic effects and
arbi-qualitatively different results (Bu¨ rger1999; Waxman trary overdominance across loci and by considering the
and Peck 1999; Waxman 2003). Nevertheless, we be-simultaneous stability of alternative equilibria at which
lieve that models with two alleles are a better approxi-the population mean can depart from approxi-the optimum.
mation to reality (where there will usually be a few GillespieandTurelli(1989) showed how balancing
discrete alleles) than are a continuum of alleles, particu-selection could arise at individual loci by averaging over
larly when pleiotropy is considered, because it takes an randomly fluctuating allelic effects. In their symmetric
extraordinary number of discrete alleles to approxi-model ofG⫻Einteractions, all alleles have essentially
mate even a two-dimensional continuum (Turelli1985; the same mean and variance of effects. With this
ex-Wagner 1989). Models with discrete alleles also pre-treme symmetry assumption, even slight fluctuations can
clude a particular multilocus genotype that produces maintain indefinitely many alleles at an arbitrary
num-the highest fitness under all conditions. By implicitly ber of loci. However, the essential interchangeability
allowing such genotypes, Via and Lande (1987) con-of the alleles implies that there will be essentially no
cluded thatG⫻E interactions could not maintain sta-correlation between the phenotypes produced by a
ble polygenic variation. given genotype across unrelated environments (i.e., two
Another critical assumption is that the temporal and environments chosen at random from the distribution
spatial scales of fluctuating allelic effects are sufficiently of environments responsible for maintaining variation;
small, relative to the timescale of selection, that we can GillespieandTurelli1989, 1990;Gimelfarb1990).
average over these fluctuations to approximate the al-Genetic variation that shows so little consistency of
ef-lele-frequency dynamics with deterministic differential fects would severely limit the resemblance between
par-equations. Both the linkage equilibrium and averaging ents and offspring across different environments.
approximations were made byGillespieandTurelli Below we explore the consequences of allowing
ap-(1989), and we explore their validity numerically with preciable differences in the mean effects of different
temporal fluctuations in allelic effects (and sex-depen-alleles. We show that under simple forms of spatial and
dent allelic effects). We conjecture that more highly temporal variation in allelic effects, the conditions for
autocorrelated temporal fluctuations would maintain the maintenance of variation become much more re- less variation (Gillespie andGuess 1978), whereas a strictive than those indicated byGillespieandTurelli coarser spatial variation can maintain more variation (1989). Nevertheless, a surprisingly simple necessary (BartonandTurelli1989;Barton1999).
ble for maintaining variation. This may reconcile the poly- frequency dynamics can be described by morphism conditions with experimental observations.
⌬pi ⫽
piqi 2
ln w
pi
MODELS AND APPROXIMATE ANALYSES
⫽Spiqi 2
共
␣2
i(pi⫺qi)⫺2␣i(z⫺ )
兲
⫹o(S) (3) We analyze in turn pleiotropic balancing selection,G ⫻ E with spatial variation and complete mixing,
(Wright1937). (The first equation above is exact for sex-dependent allelic effects, andG⫻E with temporal
one locus; the approximation is the calculation of mean variation. The connection uniting these alternative sce- fitness for the one-locus genotypes.) Assuming weak narios is that in the weak-selection limit, they lead to selection, we can approximate (3) by
essentially identical allele-frequency dynamics and
hence similar stability properties for equilibria. This is dpi
dt ⫽ Spiqi
2
共
␣2
i(pi⫺ qi)⫺2␣i(z ⫺ )
兲
. (4) somewhat surprising, since temporal G ⫻ E leads tostochastic fluctuations in allele frequencies whereas
Note that loci other thanienter these dynamics only pleiotropic balancing selection, spatial variation, and
through their contribution toz, and this is true for all sex-dependent allelic effects are wholly deterministic
of the models we consider. Note also that the allele-(but seeGillespieandTurelli1989 for motivation of
frequency dynamics depend on the allelic effects only this deterministic approximation and our results below
through ␣i ⫽ i ⫺ ␥i and z⫺ ⫽ 2兺ni⫽1pi␣i ⫹ for numerical support). We start with the simplest
deter-2兺n
i⫽1␥i⫺ . Because the scale of measurement of our ministic model to illustrate our stability analyses and
trait is arbitrary,can absorb any constants that enter then apply essentially the same analyses to “averaged”
the determination of the mean phenotype (such as the versions of more complex models involving
environ-␥iand the contributions of monomorphic loci not con-ment- or sex-dependent allelic effects. We support our
sidered in our analyses). Thus, we are free to choose average-based analytical approximations with exact
any values for the i and ␥i that satisfy ␣i ⫽ i ⫺ ␥i. multilocus numerical analyses and also use numerical
Without loss of generality, we assume that analyses to explore the properties of simultaneously stable
alternative equilibria. i⫽ ␣i/2 and␥i ⫽ ⫺␣i/2 for alli, (5)
Pleiotropic balancing selection: LetBi andbidenote
so that the alleles at locusi. We letpi,tdenote the frequency of
Bi in generation t and set qi,t ⫽ 1 ⫺ pi,t. We assume
z⫽
兺
n
i⫽1
␣i(pi ⫺qi). (6)
that selection is sufficiently weak and linkage sufficiently loose that we can ignore linkage disequilibrium. We
As shown initially byWright(1935) from an approxi-assume diploidy and random mating. Leti(␥i) denote
mation like (4) (cf.Bulmer1971), stabilizing selection the additive contribution ofBi(bi) to the trait of interest.
will generally eliminate additive polygenic variation (see We set␣i⫽ i⫺ ␥i, so that␣idenotes the average effect
Bu¨ rgerandGimelfarb1999 for a recent review). To of a substitution at locusi(FalconerandMackay1996,
maintain variation, we assume that the loci experience Chap. 7). (Table 1 provides a glossary of notation.)
As-balancing selection of some sort. The simplest such suming no dominance or epistasisfor the trait, the
popu-mechanism is overdominance, but our analysis also cov-lation mean and additive genetic variance in
genera-ers cases in which additive effects on fitness are linear tiontare
functions of allele frequencies. This may be a good approximation for a wide range of models of negative
zt⫽2
兺
ni⫽1
(pi,ti⫹qi,t␥i) and VA,t⫽2
兺
nt⫽1 ␣2
ipi,tqi,t. (1)
frequency dependence, especially if allele frequencies are not perturbed too far from equilibrium.
We assume constant Gaussian stabilizing selection on We assume that the relative contributions to fitness this trait with optimumand strengthS, so that the fit- from pleiotropic effects are 1⫺
siqˆi, 1, and 1⫺ sipˆi for ness assigned to genotypes producing mean phenotype B
iBi,Bibi, andbibi, respectively, with 0⬍siⰆ1,qˆi⫽1⫺
G (averaged over nongenetic sources of variation) is pˆ
i, and 0 ⬍ pˆi ⬍ 1 for all i. These fitnesses lead to a
w(G)⫽exp(⫺(S/2)(G⫺ )2); this produces both
domi-stable equilibrium atpˆi, a fixed parameter in the model. nance and epistasis for fitness. For weak selection, we For definiteness, we assume that these pleiotropic fitness can approximatew(G) by a linear function ofS. In the effects are multiplicative across loci and that this pleio-weak-selection limit, the population’s mean fitness is tropic selection acts before stabilizing selection in the
life cycle, with both affecting viability. However, these
as-w⫽1⫺S
2(VA ⫹(z⫺ )
2)⫹o(S), (2)
sumptions are irrelevant in our weak-selection approxi-mation. With weak selection, we can, likeBulmer(1973) whereo(S) denotes a quantity that vanishes faster than and Gillespie (1984), superimpose the pleiotropic overdominant selection on the trait-induced selection
allele-TABLE 1
Glossary of repeatedly used notation
Symbol Usage (relevant equation in the text)
Bi One of the two alleles at locusi(biis the other), (1)
K Fraction of the total genetic variance attributable to mean effects of genotypes, (44)
S Strength of stabilizing selection, (2)
VA Additive genetic variance, (1)
pi,t Frequency ofBiin generationt, (1)
pˆi Equilibrium frequency ofBiunder balancing selection alone, (7)
qi Frequency ofbi, (1)
si Intensity of balancing selection at locusi, (7)
vi si/(␣2iS) in the model of pleiotropic balancing selection, (7), or squared coefficient
of variation of the effect of substitution at locusiin the model ofG⫻E, (25)
z Average trait value in the population, (1)
z*i Contribution to the mean phenotype from all loci buti,兺j⬆i␣j(pj⫺qj), (8)
␣i (Mean) effect of a substitution at locusi, (1) and (25)
i(␥i) Additive contribution ofBi(bi) to the trait, (1) and (25)
⌬ Departure of the population mean from the optimum,z⫺ , (7b)
⌬f Under pleiotropic balancing selection, the amount by which the population would
depart from the optimum in the absence of stabilizing selection, (23c)
␦i ⌬normalized by the (mean) effect of a substitution at locusi,i.e.,⌬/␣i, (7)
Optimum trait value under stabilizing selection, (2)
ij The correlation between the substitution effects at lociiandj, (28a)
to approximate the allele-frequency dynamics by selection is eliminated and the population mean is at the optimum (⌬ ⫽ ␦i ⫽ 0). Condition (9) creates a dpi
dt ⫽ ⫺ S
2piqi
共
⫺␣2
i(pi⫺qi)⫹2␣i(z⫺ )⫹2 si
S(pi⫺pˆi)
兲
system of linear equations for thepithat will generally have a unique solution for a fixed set of polymorphic⫽S␣2i
2 piqi
共
(pi⫺qi)⫺2␦i⫺2vi(pi⫺pˆi)兲
, (7a) loci. We first focus on the stability of fully polymorphicequilibria at which allnloci satisfy (9), but we show be-where
low that precisely the same stability conditions emerge whenever two or more loci are polymorphic. Conditions vi⫽
si ␣2
iS
, ␦i⫽ ⌬
␣i
, ⌬ ⫽z⫺ . (7b)
for the feasibility of fully polymorphic equilibria are dis-cussed below, along with boundary equilibria at which Given that a deviation from the optimum of␣ireduces some or all of the loci are fixed.
fitness byS␣2
i/2,viquantifies the intensity of balancing As shown inappendix a, stability of the fully polymor-selection at locus i relative to stabilizing selection. To
phic equilibrium is determined solely by thevi. To main-make the stability analysis more transparent, we can
tain a stable polymorphism, balancing selection must rewrite (7a) as
be sufficiently strong relative to stabilizing selection (Bulmer1973). The stability of the fully polymorphic
dpi
dt ⫽ ⫺S␣
2
ipiqi
共
pi⫺ 1 2 ⫹1 ␣i
(z*i ⫺ )⫹vi(pi⫺pˆi)
兲
, (8) equilibrium depends on the eigenvalues of the Jacobian matrixA⫽ (aij), withwhere z*i ⫽兺j⬆i␣j(pj⫺qj) denotes the contribution to the mean phenotype from all loci but i.
aii⫽
pi
冢
dpi
dt
冣
⫽ ⫺Spiqi␣2
i(1⫹vi) (10a)
Stability of fully polymorphic equilibria:From (7a) we see that each locus can fall into one of three possible
equilib-and ria:pi ⫽0,pi⫽1, orpi, satisfying
aij ⫽
pj
冢
dpi
dt
冣
⫽ ⫺2Spiqi␣i␣j fori⬆j, (10b) pi⫽2vipˆi⫺ 1⫺ 2␦i 2(vi ⫺1)
(9)
evaluated at allele frequencies that satisfy (9). The fully (note that␦idepends on all of the allele frequencies),
polymorphic equilibrium is locally stable if all of the with vi and␦idefined in (7b). [Because of the simple
eigenvalues of this matrix have negative real parts. In form (3) for our approximate dynamics, only point
equi-general, it is difficult to calculate the eigenvalues. Never-libria can occur; Bu¨ rger2000, Appendix A3.] As
ex-theless, the stability conditions can be determined be-pected, the polymorphic equilibrium (9) becomespˆi if
first a completely symmetric model, as discussed by selection of intensity S acts toward an optimum ,
independently across a set of characters, labeled,i.e., Bulmer(1973), for which all of the eigenvalues and
the equilibrium allele frequencies can be explicitly de- w(G)⫽Exp[⫺兺(S/2)(G⫺ )2] (corresponding to
multiplying the Gaussian selection across characters). termined.
Suppose that the loci are interchangeable with ␣i⫽ Following the arguments leading to (7a), we obtain
␣,si⫽s, andpˆi⫽pˆ; thenvi⫽vfor alliand the equations
(9) for the equilibrium allele frequencies have a unique dpi
dt ⫽ S˜T
2 piqi
共
(pi ⫺qi)⫺ 2␦˜i⫺ 2v˜i(pi ⫺pˆi)兲
, (17a) solution:where
pi⫽ p⫽
2 ⫹(2n⫺ 1)␣ ⫹2v␣pˆ
2␣(2n⫺ 1⫹ v) . (11)
S˜T⫽兺S␣i2,,␦˜i⫽兺
S␣2i,␦i,
ST
,␦i,⫽z⫺ ␣i,
,v˜i⫽ si
ST . In this symmetric case, the stability matrix Ahas all
(17b) diagonal elements equal and all off-diagonal elements
equal.Ahas only two distinct eigenvalues, The polymorphic equilibria can still be represented by (9) withvireplaced byv˜iand␦ireplaced by␦˜i.
1⫽ ⫺Spq␣2(v⫺1) and 2⫽ ⫺Spq␣2(2n⫹v⫺1),
However, the stability conditions are more complex (12)
than those for the one-dimensional model, because the where1has multiplicityn ⫺1. Obviously,2is always stability-determining matrixAin (10) is replaced by
negative, but1 is negative if and only if
aii⫽ ⫺STpiqi(1 ⫹v˜i) (18a)
v⬎1. (13)
and Thus, as Bulmer (1973) found by assuming that
aij⫽ ⫺2pipj
兺
S␣i,␣j, fori⬆ j. (18b)z⫽ , s ⫽ ␣2S is the lower bound on the intensity of
balancing selection relative to stabilizing selection that Because of the summation in (18b), the signs of the must be exceeded to produce a stable polymorphism. eigenvalues of (18) do not depend solely on the v˜i. Essentially the same constraint onviarises for the gen- Unlike the one-character model in which at most one
eral model (7a). locus is expected to be polymorphic with vi ⬍ 1, for
The necessary and sufficient conditions derived in multiple characters withv˜i⫽0 for alliand equal allelic appendix afor stability are that either effects, the number of stably polymorphic loci can be as large as the number of traits (Hastings andHom
vi ⬎1 for alli, (14)
1989) or larger with strong selection (Gimelfarb1992). We return to this result when we consider sex-depen-or one locus (locus 1, say) hasv1 ⬍ 1, but this locus
dent allelic effects. obeys
Stability, feasibility, and positions of alternative equilibria:
Next, we consider equilibria for the one-character model
v1⬎1⫺
1 (1/2)⫹
兺
ni⫽2(1/(vi ⫺1))
. (15)
in which some loci are monomorphic. Several complexi-ties arise due to the possible simultaneous stability of For large numbers of polymorphic loci, the sum in
multiple equilibria with different numbers of polymor-the denominator is large, and so this condition is barely
phic loci and fixation of eitherBiorbiat the monomor-different from the simpler sufficient condition (14).
phic loci. First, consider the conditions for polymorphic Indeed, as shown inappendix a, a necessary condition
equilibria to be feasible. The conditions will depend on for stability is
whethervi⬎1 (recall that at most one stably polymor-(vi ⫹1)(vj⫹1) ⬎4 for alli⬆ j, (16) phic locus can violate this). Ifvi⬎ 1, (9) implies that
0⬍ pi⬍1 only if so that condition (14) is not far from being both
neces-sary and sufficient. Conditions (14) and (15) can be
vi⬎ max
冢
1⫹ 2␦i2pˆi
, 1⫺2␦i 2qˆi
冣
. (19a)
understood from Wright’s (1935) result that in the absence of balancing selection,i.e.,vi ⫽ 0 for alli, at
Ifvi⬍1, feasibility requires most one locus is expected to be polymorphic in the
weak-selection limit. At loci withvi⬎1, balancing
selec-vi⬍ min
冢
1⫹ 2␦i2pˆi
, 1⫺2␦i 2qˆi
冣
. (19b)
tion is strong enough to maintain polymorphism. Our weak-selection analysis indicates that at most one such
locus can be polymorphic with 1⬎vi ⬎ ⫺1. Unless⌬ ⫽0, (19a) constrains at least all but one of
vito exceed 1 by an amount that depends on ⌬.
Con-Multiple characters: The model readily generalizes to
multiple characters, but the resulting stability condi- versely, if stability is achieved with one locus satisfying
vi⬍1, (19b) puts an upper bound on thisvithat must tions involve an important difference that illuminates
Overall, these feasibility conditions for polymorphisms ⌬ ⫽
兺
i僆⍀1␣i⫺
兺
i僆⍀0␣i⫹
兺
i僆⍀p␣i(pi⫺qi)⫺ . (22) and the conditions described next for stability of
fixa-tion equilibria require allele frequencies that make ⌬ Substituting expression (9) for the equilibrium allele
very small. frequencies and rearranging, we find that
Consider an equilibrium at which pi⫽ 0 for all iin
the set⍀0,pi⫽1 foriin⍀1, and 0⬍pi⬍1 foriin⍀p. ⌬ ⫽ ⌬f
1⫹2C, (23a)
In this case, the stability matrix A can be partitioned
into blocks corresponding to the fixed and polymorphic where loci, because
⌬f⫽
兺
i僆⍀1
␣i ⫺
兺
i僆⍀0␣i⫹
兺
i僆⍀p␣ivi(pˆi⫺qˆi)
vi⫺1
⫺ , (23b)
aij ⫽0 fori⬆jifiis in either⍀0or⍀1, (20a)
whereas
and
aii⫽
S␣2
i
2 (2pˆivi ⫺1⫺2␦i) fori僆 ⍀0, (20b) C ⫽
兺
i僆⍀p 1
vi⫺1
. (23c)
and The population mean would depart from the optimum
by ⌬f in the absence of stabilizing selection returning
aii⫽
S␣2
i
2 (2qˆivi⫺ 1⫹ 2␦i) fori僆⍀1. (20c) the trait toward the optimum. [Without stabilizing selec-tion, the terms in the final summation in (23b) reduce to ␣i(pˆi ⫺ qˆi).] In this sense, ⌬f represents a natural
Thus, the eigenvalues governing the stability of the fixed
resting point of the system under balancing selection loci (i.e., i 僆 ⍀0 傼 ⍀1) are simply i ⫽ aii, and the
alone. Stabilizing selection generally reduces this devia-stability conditions for the subsystem of polymorphic
tion by a factorB⫽1/(1⫹2C) (as noted inappendix loci (i.e.,i僆⍀p) are just (14) and (15). Equations 20b
b, the factor is negative ifvi⬍1 for one of the iin⍀p,
and 20c show that the stability conditions for the fixed
but we ignore this special case). Thus,Bis a cumulative loci are
measure of the strength of stabilizing selection relative to balancing selection. As noted above, we generally
vi⬍
1⫹2␦i 2pˆi
ifpi⫽0, (21a)
expectvi⬎ 1 at stably polymorphic loci. For any fixed lower bound on thevi, (23c) shows that as the number and
of polymorphic loci increases, the population mean will converge to the optimum by slightly perturbing the
poly-vi⬍
1⫺2␦i 2qˆi
ifpi⫽1. (21b) morphic allele frequencies away from pˆ
i as described by (9).
Hence, the conditions for the stability of the fixed equi- The stability conditions for the full system, including libria are complementary to the feasibility conditions fixed and polymorphic loci, are detailed inappendix b. (19a) for the polymorphic equilibria withvi ⬎ 1. The The qualitative conclusion is that, for a wide range of implications of (21) can be seen by assuming, without parameter values, loci withvi⬎1 can be stably polymor-loss of generality, that⌬ ⫽z⫺ is negative and all of phic and loci withvi⬍ 1 are generally monomorphic. the ␣i are positive. In this case, increasing pi at each Moreover, although alternative equilibria may be simul-locus moves the population mean closer to the opti- taneously stable, they generally produce mean pheno-mum. Then, inequalities (21) with vi ⫽ 0 imply that, types very near the optimum. These generalizations are whenever possible, the multilocus system will equilibrate illustrated by our numerical examples below, which also so that |⌬| is less than mini僆⍀0{␣i/2}. Inequalities (21) suggest that the alternative equilibria produce similar imply that |⌬| is even smaller withvi⬎ 0. equilibrium levels of genetic variation.
Because alternative multilocus equilibria will gener- Consequences of G ⫻ E with spatial variation and
ally produce different values of⌬, and hence different complete mixing: Next we consider a deterministic ␦ifor each locus, conditions (19) and (21) do not pre- model that involves only stabilizing selection on the clude a locus from having stable alternative fixation and trait, but allows for environment-specific allelic effects, polymorphic equilibria (cf.HastingsandHom1990). which can produce balancing selection at individual In particular, ifvi is only slightly above one, the locus loci (GillespieandTurelli1989). FollowingLevene can be stably polymorphic at an equilibrium with⌬very (1953), we assume that each environment contributes near 0, but stably monomorphic at equilibria with larger a constant proportion to the random-mating pool that |⌬|. This is illustrated numerically below. forms the next generation of zygotes. In our weak-selec-Now consider⌬at equilibria. Assuming as above that tion limit, this means that we can simply average the
pi⫽0 fori僆⍀0,pi⫽ 1 fori僆⍀1, and 0⬍pi⬍1 for equations that emerge in each environment, weighting each environment by its fractional contribution to the
next generation (cf.GillespieandLangley1976). Let whereE(z*i)⫽兺j⬆i(pj⫺qj)␣jdenotes the average
contri-i,k(␥i,k) denote the effect ofBi (bi) in environment k. bution to the mean phenotype from the loci other than With weak selection, as in (4), we can approximate the i. It is easy to see that Cov(i⫺ ␥i,z*i )⫽0 ifc˜ij⫽0 for allele frequency dynamics in this environment by alli⬆ j.Gillespieand Turelli(1989) assumed this and found that interlocus correlations did not affect their polymorphism condition. We show below that this
dpi
dt ⫽ ⫺ Spiqi
2 (⫺(pi⫺ qi)(i,k⫺ ␥i,k)
2
conclusion depends critically on their symmetry assump-tion concerning interlocus correlaassump-tions.
⫹ 2(i,k⫺ ␥i,k)(z⫺ )). (24)
Analysis of fully polymorphic equilibria: At equilibrium, We assume that S and remain fixed across environ- each locus must satisfypi⫽0,pi⫽ 1, or
ments. Averaging over environments, we define
pi ⫽ 1 2⫺ ␣
i[E(z*i )⫺ ]⫹ ␣2id˜ii⫹ Cov(i⫺ ␥i,z*i )
␣2
i(1 ⫹vi)
.
E(i⫺ ␥i)⫽ ␣i and Var(i⫺ ␥i)⫽vi␣2i. (25)
(30) Thus, the effect of a substitution at locusihas mean␣i
Note that increasing d˜ii, corresponding to raising the and variance isvi␣2i, so thatviis the square of the
coeffi-cient of variation of the substitution effect. We demon- variance of effect for allele Bi relative to the variance strate that these vi play the same role in the stability forbi, decreases the equilibriumpi, consistent with the analysis of this model as do thevi defined by (7b) for general principle that selection in variable environ-the pleiotropy model. Averaging over environments, as ments tends to favor more homeostatic genotypes ( Gil-done inGillespieandTurelli(1989), we obtain lespie1974). The stability of the fully polymorphic equi-librium depends on the eigenvalues of the Jacobian matrixA⫽(aij) with elements
dpi
dt ⫽ ⫺ Spiqi
2
共
⫺␣2
i(1 ⫹vi)(pi⫺qi)⫹2␣i[E(z)⫺ ]
⫹2 Cov(i⫺ ␥i,z)
兲
. (26) aii⫽ pi
冢
dpi
dt
冣
⫽ ⫺Spiqi␣2
1(1⫹vi) (31a) As discussed after Equation 4, we can absorb constants
and that enterE(z) into, so we assume E(i)⫽ ␣i/2 and
E(␥i)⫽ ⫺␣i/2, without loss of generality. Thus,E(z)⫽
兺i(pi⫺qi)␣i. Rearranging (1), we have aij⫽
pj
冢
dpi
dt
冣
⫽ ⫺2Spiqi␣i␣j(1 ⫹c˜ij) fori⬆ j, (31b) z ⫽兺
n
i⫽1
(pi⫺qi)(i ⫺ ␥i)⫹
兺
ni⫽1
(i⫹ ␥i), (27)
evaluated at allele frequencies that satisfy (30), withc˜ij as defined in (28a). If Cov(i⫺ ␥i,z*i )⫽0, the terms where thei and ␥i have environment-specific values.
c˜ij in (31b) vanish and the stability conditions are pre-Hence, the term Cov(i⫺ ␥i,z) that enters (26) and
cisely those for the pleiotropy model, (14) and (15). In the analyses below depends on the scaled covariances,
this case, theG⫻ E model reduces to the pleiotropic
c˜ijandd˜ij, defined by
balancing selection model (apart fromd˜ii, which does Cov(i⫺ ␥i,j⫺ ␥j)⫽c˜ij␣i␣j⬅ij␣i␣j
√
vivj (28a) not affect stability of polymorphic equilibria) withpˆi⫽ 0.5 at all loci. Following the argument inappendix a, andit is easy to see that in general the stability properties of (31) depend only on theviand theijdefined in (28a). Cov(i⫺ ␥i,j⫹ ␥j)⫽d˜ij␣i␣j, (28b)
In general, positive correlations across loci are desta-wherec˜ii⫽viandijdenotes the correlation of substitu- bilizing in the sense that largerv
iare needed to achieve tion effects at lociiandj. Note thatc˜ij ⫽d˜ij ⬅0 for all stability with
ij⬎0 than withij⫽0. The destabilizing
i⬆jif either the allelic effects at different loci fluctuate effect is dramatic. The necessary condition for stability, independently or we impose the symmetry constraints, analogous to (16), is
Cov(i,j)⫽Cov(␥i,␥j)⫽Cov(␥i,j)⫽Cov(i,␥j) for alli ⬆ j. Gillespie andTurelli (1989) assumed the (v
i⫹ 1)(vj⫹1) ⬎4(1⫹ ij
√
vivj)2 for alli⬆j. latter. Under the less restrictive assumptions that Cov(i,(32)
j)⫽Cov(␥i,␥j) and Cov(␥i,j)⫽Cov(i,␥j), we have Withij⬎0 and2ij⬎3/4, (32) cannot be satisfied for
d˜ij ⬅ 0 for all i and j. In particular, d˜ii ⫽ Var(i) ⫺ three or more loci. The general stability conditions can Var(␥i), so that d˜ii⫽ 0 if Var(i)⫽Var(␥i). Separating be explicitly obtained following the procedure given the terms in (26) that depend on locusi, we have in appendix a, but they seem too complicated to be informative. However, the qualitative effects of
correla-dpi
dt ⫽ ⫺ Spiqi
2
共
␣2
i(1⫹vi)(pi⫺ qi)⫹ 2␣i[E(z*i)⫺ ] tions across loci can be seen under the symmetry as-sumptionsvi⬅vandij⬅. In this case, a feasible fully
⫹2␣2
vi⬍ 1⫹ 2ei, ifpi ⫽0, and vi ⬍1⫺2ei, ifpi ⫽1.
⬍1/2 and v⬎ 1
1⫺2. (33) (36)
As expected, positive values of Cov(i⫺ ␥i,z*i ) andd˜ii Hence, polygenic variation can be stably maintained
promote the stability ofpi⫽0. under this model ofG⫻E interactions only if the loci
Properties of polymorphic equilibria: One of our central experience at most moderate positive correlations
motivations for allowing significant differences in the among their fluctuating allelic effects, and the variance
mean effects of alleles was to determine whether appre-in effects is sufficiently large. As illustrated by our
analy-ciable heritable variation could be maintained by G⫻
sis of sex-dependent allelic effects, with only two
envi-Ethat would provide persistent selection response. We ronments, |ij| ⫽ 1 and polymorphism condition (32)
have shown that maintaining variation requires a suffi-cannot be satisfied.
ciently large coefficient of variation of the allelic effects
Stability, feasibility, and position of alternative equilibria:
and sufficient independence of the fluctuations across Consider an equilibrium withpi⫽0 fori僆⍀0,pi⫽ 1
loci. At least two biologically interesting questions fol-fori僆⍀1, and 0⬍pi⬍1 fori僆⍀p. First note that if
low. First, how similar are the phenotypes of various Cov(i⫺ ␥i,z*i )⫽0 andd˜ii⫽0, we can use the results
relatives, for instance, parents and offspring, and sec-for the pleiotropic balancing selection model with the
ond, how variable are the phenotypes produced by spe-additional constraint pˆi ⫽ 0.5 for all i. This generally
cific genotypes across the range of environments re-simplifies the analysis. For instance, the stability
condi-sponsible for maintaining the variation (cf. Yamada tions for the fixed loci reduce to
1962;GillespieandTurelli1990;Gimelfarb1990).
vi ⬍1⫹2␦i, ifi僆⍀0, and vi⬍ 1⫺ 2␦i, ifi僆 ⍀1, The second question is answered more easily than the
(34) first, because the similarity of relatives will depend on
the similarity of their environments. Even if this is where␦i⫽ [E(z)⫺ ]/␣i. Numerical examples of
pleio-known, the correlations between relatives will depend tropic overdominance presented below, which assume
on additional parameters, which do not enter the
poly-pˆi ⫽ 0.5, illustrate the approximate equilibria and
dy-morphism conditions, that describe the covariance of namics of this model.
the fluctuating effects of alleles within and across loci. New phenomena appear with Cov(i⫺ ␥i,z*1) ⬆ 0
These parameters also enter the variance for the mean andd˜ii⬆0. As noted above, positive correlations across
phenotypes produced by specific genotypes across envi-loci make stable polymorphisms more difficult to obtain
ronments. To illustrate this, we calculate the expected and positive values of d˜ii tend to lower pi. Hence, we
variance of the mean phenotype of a randomly drawn expect that positive correlations between loci andd˜ii⬎
genotype and then partition the equilibrium variance 0 will broaden the conditions for stability ofpi⫽ 0. As
in mean phenotypes to quantify the consistency of geno-before, the stability matrix A can be partitioned into
typic differences across environments. blocks corresponding to the fixed and polymorphic loci,
LetGk(g) denote the average phenotype of a specific because
multilocus genotypegin a specific environmentk(the
aij ⫽0 fori⬆jifiis in either⍀0or⍀1, (35a) same analyses apply to both spatial and temporal
varia-tion). Under our additivity assumption, whereas
Gk(g)⫽
兺
ni⫽1
Gi,k(g), (37)
aii⫽
S␣2
i
2 (vi⫺1⫺2ei) fori僆 ⍀0, (35b)
where Gi,k(g) denotes the contribution of the diploid and
genotype at locusiin environmentk. Under the assump-tions that lead to (24), the allele-frequency dynamics
aii⫽
S␣2
i
2 (vi⫺1⫹2ei) fori僆 ⍀1, (35c) depend on the moments of allelic effects only through the means and variances of the substitution effects.
with Thus, we had to specify only E(
i ⫺ ␥i) ⫽ ␣i and Var(i ⫺ ␥i)⫽vi␣i2. However, we will see that the
vari-ei⫽ ␦i⫹d˜ii⫹
Cov(i⫺ ␥i,z*i )
␣2
i
. (35d)
ance of genotypic values depends separately on the vari-ances and covarivari-ances of the allelic effects within and Thus, the eigenvalues governing the stability of the between loci. We assume that
fixed loci (i.e.,i僆⍀0傼⍀1) are simplyi⫽aii, and the
Var(i)⫽Var(␥i)⫽ci␣2i/2 and Cov(i,␥i)⫽ ici␣2i/2, stability conditions for the subsystem of polymorphic
(38) loci (i.e.,i僆⍀p) are determined by the eigenvalues of
(31), which depend only on the parameters for the so that polymorphic loci. Equations 35b and 35c show that the
Hence, in the calculations above,e.g., Equation 8,vi⫽ tal “patches” or years), it bears no simple relationship to heritability estimates, which also account for
“micro-ci(1⫺ i). [Note thatidescribes correlations between
allelic effects within loci, whereasijin (28a) describes cor- environmental” variation (i.e., phenotypic differences among genetically identical individuals within the same relations between substitution effects at different loci.]
First, assume uncorrelated fluctuating allelic effects spatial or temporal macroenvironment).
Our linkage equilibrium assumption and the defini-across loci, so that
tion of␣iimply that irrespective of correlations in fluc-Var[Gk(g)|g]⫽
兺
n
i⫽1
Var[Gi,k(g)|g], tuations within or among loci,
Varg{E[Gk(g)|g]}⫽2
兺
ni⫽1 ␣2
ipiqi. (45)
Eg{Var[Gk(g)|g]}⫽
兺
ni⫽1
Eg{Var[Gi,k(g)|g]}, (40)
Hence, for independent fluctuations across loci, whereEgdenotes averaging over the distribution of
ge-notypes in the population. A central feature of all
ran-K⫽
兺
n i⫽1␣2ipiqi
兺
ni⫽1␣2ipiqi⫹
兺
ni⫽1vi␣2i共
(1/(1 ⫺ i))⫺piqi兲
. dom environment models is that Var[Gi,k(g)|g] dependson whether genotypegis homozygous or heterozygous
(46)
at locusi (Gillespie andTurelli1989). Using (38),
The qualitative implications of (46) are most easily seen Var[Gi,k(g)|g]⫽2ci␣2i ifgis homozygous for either allele with exchangeable loci (
i.e.,␣i ⫽ ␣,ci ⫽c,i⫽ , and at locusi, and Var[Gi,k(g)|g]⫽ci␣2i(1⫹ i) ifgis hetero- p
i⫽p), for which zygous at locusi. Thus,
K⫽ pq(1⫺ )
pq(1⫺ )⫹v[1⫺ pq(1 ⫺ )]
, (47)
Eg{Var[Gi,k(g)|g]}⫽2ci␣2i[1⫺piqi(1⫺ i)]
⫽2vi␣2i
冢
1 1⫺ i⫺piqi
冣
. (41) and the stability criterion is simplyv⬎1. Equation 47 implies that K decreases as and v increase and asp departs from 0.5. Hence, for stable equilibria, K is Note that this depends on bothviandi. With
indepen-maximized when ⫽ ⫺1,v ⫽ 1, andp ⫽ 0.5. At this dent fluctuations across loci, we have
point,K⫽0.5. However, when the within-locus effects are uncorrelated, as the between-locus effects are
as-Eg{Var[Gk(g)|g]}⫽ 2
兺
ni⫽1
vi␣i2
冢
1 1⫺ i⫺piqi
冣
. (42)sumed to be,Kⱕ0.25.
In general, positive between-locus covariances reduce To understand the implications of (42), we need a
Kbecause the numerator remains constant butEg{Var scale-independent quantification of this expected
within-[Gk(g)|g]} in the denominator increases. The effects of genotype variance. By analogy to broad-sense
heritabil-these covariances depend not only on the covariances ity, we can define an index for the stability of
environ-of substitution effects,i.e., Cov(i ⫺ ␥i,j ⫺ ␥j) as de-ment-dependent genetic effects as the fraction of the
scribed by (28), but also on the covariances between total genetic variance (across both genotypes and
envi-the individual alleles at each locus, i.e., Cov(i, j), ronments) attributable to the mean effects of different
Cov(␥i,␥j), and Cov(i,␥j). Nine different expressions genotypes. In general, averaging over both
environ-for Cov{[Gi,k(g)|g], [Gj,k(g)|g]} are generated by the ments and genotypes, we have
three genotypes at each locus. To illustrate the quantita-Var[Gk(g)]⫽Varg{E[Gk(g)|g]}⫹Eg{Var[Gk(g)|g]}, (43) tive effects, we focus on the completely symmetrical case explored by Gillespie and Turelli (1989) and where, as indicated, the inner expectations on the
right-Gimelfarb(1990) with hand side are taken over environments and the outer
expectations are taken over genotypes. The first term Cov(
i,j)⫽ Cov(␥i,␥j)⫽Cov(i,␥j) is the variance of the mean genotypic values (i.e., the
“main effect” of genotypes) and the second is the aver- ⫽ Cov(
j,␥i)⫽
B
√
ci␣2icj␣2j2 . (48)
age across-environment variance for individual geno-types. We define the “consistency” of these genotypes as
As discussed following Equations 28, this implies that Cov(i⫺ ␥i,j⫺ ␥j)⫽0 for alli⬆j, so that the allele
K⫽ Var{E[Gk(g)|g]}
Var[Gk(g)]
. (44)
frequency dynamics are still approximated by (29) with
d˜ii⫽Cov(i⫺ ␥i,z*i)⫽0. In particular, in the
symmetri-Knear 0 implies that the differences among mean ef- cal case withc
i(1 ⫺ i)⫽ v for all i, the polymorphic fects are small relative to the standard deviations of equilibrium is stable wheneverv⬎1. For exchangeable genotypes’ effects across environments, andK near 1 loci satisfying (48), (47) is replaced by
implies relatively large mean effects (e.g., K ⫽ 1 with
constant allelic effects). Because K focuses on parti- K⫽ pq(1⫺ )
pq(1⫺ )⫹v[1 ⫺pq(1⫺ )⫹ (n⫺ 1)B] . tioning the variance of genotypic means within and
Thus, for any positiveB,Kapproaches 0 for large num- the qualitative conclusion that sex-dependent, additive allelic effects cannot maintain stable polygenic varia-bers of loci (seeGimelfarb1990 for an analogous
re-sult). The implications of these upper bounds onKfor tion. From our sexes-averaged approximation, we ex-pect that at most one locus can remain stably polymor-the maintenance of variation byG⫻Einteractions are
considered in thediscussion. phic under weak selection. In fact, however, several
numerical examples described below indicate that up
Sex-dependent allelic effects: A special case of this
multiple-environment model approximates allele-fre- to two loci may be stably polymorphic for loose linkage. This shows that our averaging approximation misses quency dynamics with sex-dependent allelic effects. In
this case, the two sexes are the alternative “environ- some of the subtleties of sex-dependent selection, while accurately capturing its inability to maintain variation ments.” As first argued byHaldane(1926) and
demon-strated rigorously for one locus by Nagylaki (1979), at many loci. The stable two-locus polymorphisms we find are reminiscent of Hastings and Hom’s (1989, the dynamics of weak, sex-dependent viability selection
1990) results concerning pleiotropic effects on two char-can be approximated by simply averaging the fitnesses
acters. A more careful analysis of sex-dependent allelic of each genotype over the two sexes. This is equivalent
effects requires distinguishing the allele frequencies in to averaging the allele-frequency dynamics as in (26),
the two sexes, so that at linkage equilibrium the stability but now each random variable takes on only two values.
analysis for n loci involves 2n variables rather than n. This greatly simplifies and constrains the expressions
This is discussed elsewhere. for the coefficients of variation and the correlations of
Temporal variation andG⫻E:Finally, we apply our substitution effects across loci. For instance, if␣f,i(␣m,i)
analyses to generalize the treatment byGillespieand denotes the effect of a substitution at locusion females
Turelli(1989) of temporally fluctuating allelic effects. (males), we have
Our approximation is based on averaging over the distri-bution of allelic effects to approximate the stochastic
vi⫽
冢
␣f,i⫺ ␣m,i
␣f,i⫹ ␣m,i
冣
2
. (50)
dynamics by a set of deterministic equations identical to those obtained for G ⫻ E with spatial variation. As The condition vi ⬎ 1 requires that ␣f,i and ␣m,i have
noted below, this approximation relies on weak selec-different signs. Thus, if we use the convention that each
tion. Ultimately, the usefulness of our approximations
Bi denotes the allele that increases the trait value in
depends on their ability to predict the maintenance of females, the polymorphism conditionvi⬎1 implies that
variation with biologically plausible levels of selection eachBi mustdecreasethe trait in males. By considering
and environmental fluctuation. We present numerical the symmetrical model with ⫽ 0, it is easy to see,
results below, suggesting that our approximate polymor-however, thatvi⬎1 cannot suffice to maintain polymor- phism conditions are surprisingly accurate. Our deter-phism. The multilocus recursions for the allele
frequen-ministic analysis does not address the fluctuations of cies depend separately on the fitnesses assigned to each
allele frequencies inherent in stable polymorphisms genotype in males and females. When ⫽0, our
sym-maintained by temporal fluctuations. This is explored metrical selection model implies that altering the signs
numerically below.
of all of the allelic effects in one sex will not change As with spatial variation, under particular symmetry the fitnesses. Hence, for any assignment of allelic effects, assumptions concerning the interlocus correlations in identical dynamics must emerge if all of the signs of fluctuating allelic effects [see (28) and (29)], the deter-allelic effects in one sex are reversed. If the initial assign- ministic approximation is precisely equivalent to the ment of effects satisfiesvi⬎1 for alli, by reversing the pleiotropy model analyzed above. As with spatial varia-signs of effects in one sex, we get identical dynamics tion, the critical parameter governing the stability of but the new values ofviare the reciprocals of the old. polymorphism at each locus is justv
i, the squared coef-The additional constraint required for stable poly- ficient of variation of the substitution effect at that locus. morphism involves the correlations in the fluctuat- Because we obtain identical approximations for tempo-ing effects across loci. Our convention of labeltempo-ing the ral and spatial variation, we discuss the analytical ap-alleles so thatBiandBjincrease the trait value in females proximations only briefly.
implies that The assumptions of this model are the same as with
spatial variation, except that the allelic-effect
parame-ij⫽ 1 for alli⬆j (51)
ters,i,tand␥i,t, vary across generations. Note that (25) (indeed, a two-valued bivariate random variable can allows for arbitrary correlation between the fluctuating have only correlations ⫾1). Thus, constraint (32) im- effects of the two alleles at a locus. Within-locus correla-plies that a fully polymorphic equilibrium cannot be tions ofi,tand␥i,tdo not explicitly affect the dynamics, stable, although sex-dependent selection can readily because selection depends only oni,t ⫺ ␥i,t. However, maintain polymorphism at one locus (Kidwell et al. as discussed in the context of spatial variation, intralocus
1977). correlations can significantly affect the properties of the
scale of parameter variation. As before, the basic re- to be fixed within any one generation. With high levels of positive autocorrelation for allelic effects,K would cursion is
remain high even when averages are taken over several generations. Thus, temporalG⫻Emay maintain high ⌬pi,t⫽ ⫺
Spi,tqi,t
2
共
⫺(pi,t⫺ qi,t)(i,t⫺ ␥i,t)2
levels of genetic variation with consistent differences among genotypes over the timescale of reasonable ex-⫹ 2(i,t⫺ ␥i,t)(zt⫺ )
兲
⫹o(S). (52)perimental analyses. This point is elaborated below. A central assumption of our analysis is that selection
is weak;i.e., S Ⰶ 1. We also assume that the timescale
NUMERICAL ANALYSES
of allele-frequency change is slower than the timescale
of the environmental fluctuations, for instance, that the Pleiotropic balancing selection: The number of pa-successive environments are independent or at most rameters in this model makes it impractical to explore “weakly autocorrelated” (Gillespie and Guess1978). equilibria and dynamics systematically across the param-These assumptions allow us to (i) average the random eter space. However, the qualitative features of alterna-fluctuations over time and (ii) approximate the dynam- tive equilibria and dynamics are illustrated by the follow-ics of the discrete-generation stochastic model (52) by a ing two numerical examples. In both examples, we system of deterministic differential equations. [Because assume for simplicity thatpˆi⫽1/2 at all loci. Loci with the leading term in (52) is proportional toS, the infini- extreme values ofpˆi are less likely to be polymorphic tesimal variance in a diffusion approximation vanishes, because of the constraints associated with the feasibility leaving a deterministic limit (cf.GillespieandTurelli conditions (19) and the increased influence of genetic 1989).] Taking the expectation of the right-hand side drift. We concentrate on observable quantities such as of (52) over the fluctuations in allelic effects, ignoring mean fitness, deviation of the trait mean from the opti-the higher-order terms and going to opti-the continuous- mum, and genetic variance.
time limit, we obtain the time-averaged, weak-selection Example 1—alternative equilibria:To illustrate the impli-approximation (26) used above to discuss spatial varia- cations of the stability and feasibility conditions, we first tion with complete mixing. We have not made any ex- consider an example in which the␣i andvi at 20 loci plicit assumptions about the temporal correlations of were drawn independently from gamma distributions the fluctuating parameters. However, our analysis im- (Table 2), and the optimum is ⫽ 0. Because of this plicitly assumes that autocorrelations decay faster than symmetry, the equilibria come in pairs, the elements of the timescale of allele frequency change (1/S). which have⌬of opposite sign, produced by replacing The approximate polymorphism conditions are pre- eachpiby 1 ⫺pi.appendix c describes the procedure cisely those obtained for the spatial model considered for finding the alternative stable equilibria. Note that above. Again, the central parameters governing the sta- since all of the equilibria discussed have some mono-bility of polymorphisms are thevi, which describe the morphic loci, the polymorphic loci must produce a squared coefficient of variation of substitution effects mean near an optimum different from 0, which cannot at individual loci [see (25)], and theij, which describe be achieved by multilocus heterozygotes or by any com-the correlations between substitution effects at differ- bination of homozygotes. This “effective optimum,” de-ent loci [see (28a)]. In particular, when ij ⫽ 0, we notedeff, is
expect that loci satisfying vi ⬎ 1 will tend to remain
eff⫽ ⫺
兺
i僆⍀1
␣i⫹
兺
i僆⍀0␣i; (53)
polymorphic if the expected population mean is close enough to the optimum. Loci withvi⬍1 will generally
not be stably polymorphic, and they will fix for alleles its values are given in Table 2.
For the parameter values in Table 2, there are seven that produce an expected population mean very near.
As before, we predict from (32) that no stable multilocus pairs of distinct stable and feasible equilibria (or, more accurately, seven pairs of allele frequencies that satisfy polymorphisms can be maintained for loci with large
positiveij. the constraints for stable and feasible equilibria given
by our approximations); only the stable allele frequen-One important difference between the spatial and
temporal models concerns the interpretation of the con- cies that produce negative⌬are shown. All of the equi-libria share similar properties: mean fitnesses are within sistency index,K, defined by (44), and its relationship
to empirical observations. The key point, as made by 0.55S of each other, and the deviation from the opti-mum is |⌬|⬍0.373 (relative to a mean allelic effect of GillespieandTurelli(1989, 1990), is that the
tempo-ral variation responsible for maintaining variation need ␣ ⫽1 and a range of genotypic values |G| ⬍ 21.07). Barring the most extreme pair of equilibria (presented not be observable over a few generations. For instance,
even though the “true” value ofK, obtained by averaging in the first column of Table 2), mean fitnesses are within 0.0051Sof each other, and the deviation from the opti-overallof the environments responsible for maintaining
TABLE 2
Multiple stable equilibria maintained by stabilizing selection on a single trait and overdominance
Allele frequencies (pi) at alternative stable equilibria (ei)
Locus ␣i vi e1 e2 e3 e4 e5 e6 e7
1 2.27458 0.048 0 0 0 0 1 0 0
2 1.92538 0.181 0 1 1 1 0 1 1
3 0.320365 0.220 1 0 1 1 0 1 1
4 0.054810 0.099 1 1 0 0 0 1 1
5 0.03597 0.486 1 1 0 1 0 0 0
6 0.004355 0.906 1 1 1 1 1 1 0
7 0.000191 0.792 1 1 1 1 1 1 1
8 0.000042 0.102 1 1 1 1 1 1 1
9 0.000053 3.364 1 1 1 1 1 1 1
10 0.761252 1.034 1 1 1 0.6995 0.7657 0.5248 0.5652
11 0.011769 3.528 1 1 0.5534 0.5174 0.5232 0.5022 0.5057
12 0.02935 2.720 1 0.9072 0.5315 0.5103 0.5137 0.5013 0.5034
13 0.21609 1.387 1 0.7457 0.5190 0.5062 0.5082 0.5008 0.5020
14 0.325396 1.038 1 0.7048 0.5158 0.5052 0.5069 0.5006 0.5017
15 0.0502681 3.936 1 0.6393 0.5108 0.5035 0.5047 0.5004 0.5011
16 0.037032 5.905 1 0.6132 0.5086 0.5029 0.5038 0.5004 0.5009
17 0.224072 1.906 1 0.6012 0.5078 0.5025 0.5034 0.5003 0.5008
18 4.50501 1.357 0.7321 0.5128 0.5010 0.5003 0.5004 0.5000 0.5001
19 3.70138 4.877 0.5260 0.5014 0.5001 0.5000 0.5000 0.5000 0.5000
20 7.28178 5.723 0.5108 0.5006 0.5000 0.5000 0.5000 0.5000 0.5000
eff 2.8141 0.4862 0.0384 0.0430 0.0573 0.0053 0.0141
⌬ ⫺0.3728 ⫺0.0206 ⫺0.0016 ⫺0.0005 ⫺0.0007 ⫺0.0001 ⫺0.0002
VA 41.2927 43.5909 43.6135 43.6161 43.6157 43.6166 43.6165
L/S 122.3500 121.802 121.797 121.797 121.797 121.797 121.797
L⌬/S 0.0695 2.1⫻10⫺4 1.2⫻10⫺6 1.3⫻10⫺7 2.4⫻10⫺7 2.1⫻10⫺9 1.4⫻10⫺8
LV/S 20.6464 21.7954 21.8068 21.8080 21.8079 21.8083 21.8083
LB/S 101.3340 100.0070 99.9904 99.9891 99.9893 99.9889 99.9889
Allelic effects␣iand relative overdominancevifor 20 loci were drawn from independent gamma distributions
with means 1, 2, respectively, and variance 4. The range of the trait is |z|⬍21.07, and the maximum possible
variance isVA⫽48.11. With an optimum at ⫽0, there are 14 stable equilibria (according to our approximate
stability criteria); only 7 are shown here, since the other 7 differ by only a sign change (and replacing thepi
by 1⫺pi). The last four rows give total genetic load and its three components [as defined in (56)], scaled
relative toS.
in the combinations of fixed loci. As expected from
|⌬| ⬍ ␣i(vi⫺ 1)
2 for alli僆 ⍀p. (54)
the stability conditions for fixation equilibria (21), the polymorphic loci that differ among equilibria tend to
Hence, for these loci to remain polymorphic, |⌬| must be those of small effect, and the different combinations
be less than mini僆⍀p{␣i(vi⫺1)/2}. Conversely, if the loci of polymorphic and monomorphic loci all bring the
in⍀0are to remain stably fixed, (21a) requires
population mean very close to the optimum. This ex-plains the similarity in overall fitness properties of the
|⌬|⬍ ␣i(1⫺vi)
2 for alli僆⍀0. (55)
different equilibria. Given that selection is simply tend-ing to climb toward local fitness optima, it is not clear
whether equilibria with higher mean fitness will tend With multiple loci contributing to the character, (54) and (55) ensure a very close approach to the optimum. to be reached preferentially.
We can gain some insight by considering the feasibil- Because the deviation from the optimum is small, allele frequencies at polymorphic loci with significant effects ity conditions for the polymorphic equilibria, (19), and
the stability conditions for the fixation equilibria, (21). are close topˆi. For small⌬, the allele frequencies should be near1⁄
2(1/(1⫺vi))⫹pˆi(vi/(vi⫺1)), which is a com-For definiteness, we assume that⌬ ⬍0 and ␣i⬎0 for
alli, so that directional selection will tend to increase promise between the unstable equilibrium under stabi-lizing selection (first term) and the stable equilibrium thepi. As before, we let⍀pdenote the set of polymorphic
loci and⍀0 the set of loci withpi ⫽ 0. Withpˆi⫽ 1/2, under balancing selection (second term). Forpˆinear 0.5, the genetic variance will beⵑ1⁄
2兺␣2i, where the sum is the feasibility condition (19) requires that for the loci