Biolinguistics 5.4: 284–285, 2011 ISSN 1450–3417 http://www.biolinguistics.eu
Upcoming Changes in
Biolinguistics
Kleanthes K. Grohmann & Cedric Boeckx
This issue marks the completion of the 5th volume of Biolinguistics. In these five years, we have managed to produce a traditional journal-like publication — free of charge for readers and contributors alike, and open to all. The “traditional journal-like publication” is a journal that comes in volumes, one per year, with each volume further divided into regularly appearing issues. In the case of Bio-linguistics, our four issues per volume correspond to the four seasons and have been scheduled to appear at around the same time each year.1 (As an exception, since Biolinguistics first appeared in late 2007, the first publication was a single-issue volume.) Throughout volumes 2 to 5, we have been fortunate enough to receive contributions on interesting topics which we could thematically group into special issues, and for space and time reasons these counted as double issues, a practice we expect to continue in the future. Aiming for state-of-the-art research and the dissemination of cutting-edge ideas, within a variety of topics and even (sub-)fields in biolinguistics, we also try to ensure quality publication through a proper peer-review process for submitted articles and briefs.
However, this is about where the similarities to a “traditional journal-like publication” stop. We don’t have a publisher behind us, we don’t even have a financial support system that would allow us to contract professional editorial staff, so in sum, we don’t have much in terms of professional infrastructure that would allow us to emulate a “traditional journal-like publication” further. We’ve tried our best over the first few years but have reached our limit, most pressingly in terms of time. Leaving aside the review process itself, each issue costs us around 100 hours of working time formatting, proof-reading, and editing the material. Our resources do not allow this to continue, so we will be implementing some changes from volume 6 onwards, starting in 2012:
(1) We urge all authors to follow the Biolinguistics style sheet to a dot, as we will simply be unable to spend all these extra hours formatting their contributions for them; this concerns in particular references for which we have a style sheet but which most often are not formatted or even listed properly, and which subsequently take up a fair amount of our copy-editing time. In the past we have routinely added references that the authors mentioned in their texts but failed to list in the reference sections, or corrected them. We simply won’t be able to do this anymore.
(2) We urge all authors to ensure that their contributions to Biolinguistics are fully proof-read by native speakers and conform to general, high-quality academic writing in English concerning style, grammar, punctuation, etc. (especially as specified in the style sheet) — and in turn will not spend all these extra hours proof-reading and copy-editing contributions for authors.
Failure to comply with either (1) or (2) may constitute grounds to reject a fully reviewed, revised, and even otherwise accepted piece; that is, acceptance for publication in Biolinguistics is contingent on complying with the editorial policies of the journal. This will be stated clearly in due time on the journal’s website at
http://biolinguistics.eu/index.php/biolinguistics (or simply biolinguistics.eu). We are aware that these policies may lead to some visual changes in Biolinguistics, namely, that the homogenous style and layout of the journal will not be as consistent as it has been up to now. But we believe that this is a small price to pay for a free-for-all journal without major professional infrastructure and financial support. It also reflects the collective spirit of the community a little better, which means that in order for Biolinguistics to be successful as a free, open-access journal, everyone needs to contribute.
On this note, we would like to remind our readers that the entire project can only be successful if the growing community of biolinguists chips in, that is, authors vis-à-vis submitters and peer-reviewers. It is an obvious fact that we depend on the submissions of articles and briefs to the journal if we want to reach our goal of publishing relevant cutting-edge and trend-setting research on a regular basis. But we also need to be able to rely on more reviewers who accept our requests for review positively and promptly, and then deliver in a timely fashion. As an online journal, we have the advantage of being able to publish very soon after acceptance, but it requires some effort on all parties involved.
Continuing this ‘community spirit’ theme, we would appreciate if potential reviewers would not approach Biolinguistics the way they may deal with more traditional journals in linguistics, but make it a priority to deliver on time reviews, following the practice in other fields.
On the positive side, we are delighted to announce that, thanks to the hard work of Charles Reiss and his team, especially Melinda Heijl, we will very soon be able to offer a LaTeX style sheet for Biolinguistics as well! It will be a close enough mirror of the rather intricate Word-formatted style sheet, which will also look a little different from volume 6 onwards.
Lastly, we’re happy to announce our newly forged cooperation with EBSCO Publishing to disseminate the journal further. We are also participating in DOAJ, the Directory of Open Access Journals (http://www.doaj.org/doaj?func= openurl&issn=14503417&genre=journal), and we recently applied for inclusion in Thomson Reuters products (among others, for the ominous ‘impact factor’). Basically, we will continue to look for ways of improving the journal, and we welcome any suggestions and help in this domain.
Biolinguistics 5.4: 286–334, 2011 ISSN 1450–3417 http://www.biolinguistics.eu
A Biolinguistic Approach to the Vocalizations
of
H. Neanderthalensis
and
the Genus
Homo
Lluís Barceló-Coblijn
This paper revisits the old question about the possibilities and aptitudes H. neanderthalensis had for vocalization. New evidence will be discussed that moves the discussion beyond traditional interest in the presence and inter-pretation of the fossil record and its comparison with the closest species to H. sapiens, like chimpanzees and gorillas. An interdisciplinary perspective on the analysis coupled with information gathered from neuropsychology, genetics, and comparative psychology will prove useful for obtaining a new vision in biolinguistics, so that neurocognitive activity becomes important thanks, above all, to the comparison with other species. Finally, it will be argued that it is still plausible to accept the hypothesis on the formation of a vocal capacity prior to the cognitive faculty of language, and independent of it, so that Neandertals were probably able of vocalizing voluntarily, with communicative intentions and in a sophisticated way.
Keywords: air sacs; Homo (H. neanderthalensis); hyoid bone; vocal tract; von Economo neurons
1. Introduction
The issue about language and the communicative possibilities of Neandertals is not new at all. Indeed, it has been revisited from time to time. Intuition has traditionally told us that, at the moment when both hominins — H. sapiens and H. neanderthalensis — met, they had to be able to establish some kind of communi-cative interaction. This kind of thought has led people to take for granted that Neandertals possessed a cognitive faculty of language like modern humans do. Although this is possible, I argue that, contrary to that first impression and to
what could apparently be most likely (“what common sense tells us”), it is not so obvious that H. neanderthalensis could communicate as we do. Moreover, I will put forward the idea that vocalization is a capacity almost independent from the capacity we humans have to create linguistic constructions that can be conflated to semantic macro-concepts — though it could have co-evolved along with this latter capacity. Our approach takes into account pieces of evidence from several areas of science in an attempt to contribute with a much wider vision than the one we can get exclusively from the hyoid bone. I will apply a quite classic methodology in traditional philology: If one has not got any written proof of a lexical form that could have existed, while at the same time being certain of its existence on the basis of other indirect evidence (e.g., linguistic comparison or reconstruction), then a symbol (*) is applied to the alleged word until a document is found that can prove it. Instead, if one thinks that there is not enough evidence, or that it is not conclusive, a symbol (X) will be applied. In the case of a positive conclusion, the symbol will be (√). Let’s see some samples not directly related to vocalization:
Trait H. neanderthalensis H. sapiens
Bipedalism √ √
Chin X √
Hyperboreal adaptation1 * X
Table 1: Examples of traits and their evaluation, where (√) means ‘positive/present’, (X) means ‘negative/absent’ and (*) means ‘probable though not confirmed’.
Thus, we can see how ‘bipedalism’ is clearly inferable from the osseous morph-ology. Hence both species have a confirmation symbol for this trait. Instead, ‘chin’ is confirmed in H. sapiens, but discarded in the H. neanderthalensis case. Finally, ‘hyperboreal adaptation’ is clearly absent in modern humans, but not fully rejectable in the case of Neandertals. So I apply an asterisk because there are signs that this is a trait of this species, though more proofs and research are needed in order to finally confirm it.
Next, I will review some traits I consider particularly relevant regarding human vocalization, and I will try to extrapolate them to the Neandertal case. The idea is to draw some conclusions that help me offer a more complete and enriched picture of the issue. Traditionally (and understandably), attention has specially been focused on the analysis of the fossil record, that could reliably indicate a linguistic capacity in this species, so close to H. Sapiens from an evolutionary point of view. As we will see, the issue has become more complex than previously thought. Thanks to interdisciplinary work, even some apparently
insurmountable obstacles, like the fact that brain does not fossilize, can be overcome in a minimal (but substantial) way. Data from different scientific fields come together in such a way that different areas become linked, so that the issue can be studied from different, but confluent perspectives. In what follows I will also use the comparative method. Comparison between species provides us with clues about what is inferable from other species through homologies and convergences. However, I am aware of the limitations of this method and of the necessary caution in order to deal with the presence or absence of a particular trait in an extinct species.
The list of the traits that will be object of analysis in the present work is as follows:
Trait Brief Definition
Larynx It is an organ for phonation, an air conduct and a sphinteric mechanism which goes from the base of the tongue to the traquea.
Vocal Tract Animal cavity where the sound, produced in its source (the larynx), is filtered.
von Economo Neurons
Neurons characterized by its spindle shape and only present in the anterior cingular cortex, in the disgranular prefrontal cortex and in the frontal insula.
Lack of air sacs Absence of laryngeal air sacs which were connected to the hyoid bone.
Mirror Neurons Neurons that fire when the animal performs or observes an action by a conespecific.
FOXP2 Gene and protein related to specific language impairments. Region 2-4 kHz Region of the sound wave curve that contains relevant
acoustic information for humans.
Table 2: Traits that have been related to speech and vocalizations
As we can see, traits in Table 2 go well beyond the traditional way to set out the problem of vocalizations in extinct hominids. So, I have bet for inter-disciplinary work, importing information from genetics, neuropsychology, and acoustic models. This implies that the physiology of the vocal apparatus loses part of its leading role, favoring an approach that takes into account the role of some brain regions and, more concretely, of some neurons related to particular aspects of human cognition, language and communication. Thus, what matters for vocalization is not just anatomy, but also the neural areas involved in its control.
the discussions about the role of the hyoid bone in the larynx, as well as the possible autapomorphic character of the morphology of this bone in modern humans, as opposed to the rest of great apes. I will also discuss the hyoid bone’s orifice where the so-called air sacs are connected, which is lacking in humans but present in all other great apes.
Finally, I contend that vocalization is to a large extent independent of the emergence (in H. sapiens) of a capacity until now undetected in other species — I mean a capacity to create meaningful linguistic structures, and to endlessly code thoughts and reflections. Thus, I think there is a clear difference between speech and language, as shown in modern linguistics. Moreover, as a result of my analysis, I will also defend that the critic point in the Neandertal vocalization issue — once theoretically separated from the above mentioned linguistic capacity — is whether or not this hominin had the capabilities for motor control of orofacial and lingual muscular structures and, in the last analysis, the same cognitive capabilities related to linguistic sound processing (i.e. the phonetic and phonological aspects).
1.1. Some Basic Notions from Evolutionary Studies
In order to avoid ambiguities, I will use some traditional concepts from cladistics which are useful for describing the characteristics of particular species, particularly specific changes that attract the attention of scientists. Thus, in cladistics (seeKitching et al. 1998), a derived change is called apomorphy. Whereas adding to it the prefix sin- indicates that the character is shared with other species and their common ancestor, the prefix auta- indicates that the character is an exclusive novelty of a particular species. Instead, a homoplasy is a new character shared by two or more non-related species. Thus, (i) an example of an autapomorphy is the loss of hair in humans, but not in other primates, (ii) an example of a sinapo-morphy is the loss of hair in cetaceans, but not in other mammals, and (iii) an example of a homoplasy is the fact that both bats and birds have wings.
Figure 1: Cladistic concepts
explore the cognitive abilities related to both production and perception. In this way we will see how they — according to experimental work in this field on the one hand, and to the comparative method on the other hand — could have been very similar in the hominid ancestors of H. sapiens, like H. heidelbergensis, and, by extension, H. neanderthalensis. In doing so we can judge better whether it is a case of autapomorphy, sinapomorphy, or homoplasy.
This methodology is also applied in the third section, where I review some aspects of the neuronal apparatus sustaining part of the human cognition related to speech. I take an integrative view according to which the brain has neither evolved de novo, nor layer after layer, as if the only new and mutable part were just the cortex. The approach I put forward agrees with a vision of the brain according to which this organ has undergone different selective pressures and hazardous episodes (e.g., mutations) that made possible not only the expansion of old areas (e.g., prefrontal cortex), but also the slow and gradual modification of already extant structures (e.g., current basal ganglia). Therefore, I am cautious when inferring the presence of certain neurons: on the one hand mirror neurons, and on the other hand von Economo neurons. The former have been detected in several representatives of the primate world and it is argued that they could also be part of human brains; the latter constitute a kind of neurons which, from an evolutionary point of view, is very new in primates. In fact, they are present in only one family of hominoids: the Hominidae (see Figure 2), which covers the extant species of great apes. These neurons are particularly numerous in the tribe Hominini (both genera Homo and Pan).
Figure 2: Classification of Hominoidea
approach about the genus Homo because it reorganizes the vision one could have about it and, in consequence, it has implications for the conception one could have about a particular fact related to speech.
Finally, the comparative method is also applied at a molecular level. Thus, in the fourth section I will review succinctly the most important aspects of the gene FOXP2 (and of its protein) as regards the role it plays in vocalization and in the control of orofacial muscles, and other genes related to sound perception as well. This gene is interesting because it is related to some language impairments, but also because the dating of its evolution is quite recent. As a matter of fact, it could be circumscribed to the genus Homo or even only to a subset of it.
Figure 3: A possible classification of the subtribe Hominina
2. On Vocal Apparatus Reconstructions: The Archaeological Method
for debate for a long time. Nevertheless, the lack of data sometimes forced resear-chers to get into a high level of speculation, so much so that in 1866 the Société de Linguistique from Paris banned this topic as unsuitable for scientific research. The discovering of Neandertal remains and its acknowledgement as a species turned out the status of the modern human. This ‘cousin’ was seen somehow as a kind of ‘rival’. The modern human had been considered by many people as the final milestone of evolution. Since then, there has appeared a need to look for the factors that make us humans. One proposed factor was language. This apparently unique human capacity became center-stage in a great debate that still endures nowadays. Language attracted attention in the evolutionary debate, due to its potential explanations of facts still on discussion, like the extinction of Neandertals.2 Leaving aside the enormous responsibility suddenly acquired by speech concerning the probability of a species survival, Philip Lieberman said something that posterior studies have confirmed — although it is by and large not so radical in their conclusions: In some cases, a language or any other social factor like religion could be an important, if not essential, factor in the genomic evolution of a population.3
2.1. The Role of Fossil Record in Models and Reconstructions
Lieberman & Crelin (1971) carried out a study that heated the Neandertal issue, and so began a still enduring debate. The authors reconstructed the larynx of the specimen found in La Chapelle aux Saints in order to verify whether or not this Neandertal had, from a morphological point of view, the basics required to produce the vowels that are typical in modern human phonetics. This model was then compared with the larynx of both modern human newborns and chimpan-zees. Their conclusion was that the Neandertal laryngeal structure was more similar to that of a human newborn than to that of an adult human. Possible and impossible sounds for Neandertals are reproduced in Table 3. It is important to keep in mind that, among the vowels they could not produce, there are three vowels which are considered universal in modern human phonetics (Maddieson 1984). Furthermore, these are the most stable vowels, so that they are considered the minimal repertoire a language can have: [u], [i], [a].4 The authors argue that modern humans can produce them due to their high larynx position. How is it possible? According to current phonetics, the human vocal system is based on two tubes, two cavities: The first one is horizontal, from the lips to the velum, and a second one is vertical, from the velum to the glottis (see Figure 4).
2 For example, Lieberman (1992) has argued that the kind of ‘speech’ held by Neandertals was the genetic factor that provoked their extinction.
3 Examples of this are the results of several works on Basque population (Alonso et al. 2005, Alzualde et al. 2006), Catalan population from Mallorca descending from Jewish groups (Picornell et al. 2005), or the Catalan population from Ibiza, whose mitocondrial DNA could directly descend from Phoenician people (Tomàs et al. 2006, Zalloua et al. 2008). These social factors (same language, religion, or even genre) do not work on the genome directly, but they have an influence on mating selection and, hence, indirectly on the future genetic re-combinations.
Table 3: The Neandertal vocal sounds from Lieberman & Crelin’s (1971) model
While in adult humans these two cavities have similar length, newborns’ larynx is higher, so that they are not able to emit the same range of sounds. The same happens in chimpanzees. On the one hand, they have a high larynx when compared with humans’. On the other hand, the horizontal cavity is longer than the vertical one. The most provocative conclusion, according to Lieberman & Crelin, is that the drop of the larynx has not other adaptive advantage than benefiting oral communication, while the price is the danger of choking, too high if no compensation makes it bearable.
Later on, Lieberman (1973) speculated on an evolutionary process divided in three stages: (1) increase of confidence in vocal communication for activities like hunting; (2) emergence of a vocal tract which is able to produce acoustic signals which are both distinctive and resistant to articulatory errors; (3) evolution of neural mechanisms that made use of the pre-adapted5 properties of the supra-laryngeal vocal tract for a faster codification of communication. This idea has been redesigned and enhanced through time: It would have been a gradual process that flowed into the current human capacity for voluntarily articulating sounds, which would have been acquired, since H. erectus6 (Lieberman 1998).
Thus, according to Lieberman & Crelin, the ‘key factor’ of speech evolution would be the specific place where the hyoid bone was placed — a contention criticized by Falk (1975): It was based on a too high reconstruction of the hyoid bone position. Falk pointed out that Lieberman & Crelin did not take into account the requirements of ‘deglutition’, nor the age of the subject in relation to the distance of the hyoid bone below the mandible, nor the sexual dimorphism in the development of the thyroid cartilage. Moreover, Falk remarks that, in chimpanzees, the corpus of the hyoid bone “appeared as continuation of the laryngeal air sacs” (Falk 1974: 125).7
5 The author makes use of the concept ‘pre-adaptation’, also used by Darwin (1871): “[N]atural selection channeled development in particular directions of previous modifications selected for some other role”. This use of the term ‘pre-adaptation’ is not entirely adequate and differs from Darwin’s notion, since it should be an exaptation. The most important is the idea that follows: In organic evolution, there is a continuous exploitation of already existent structures.
6 Partially in support of this idea, MacLarnon & Hewitt´s (1999) conclusions point out that an enhancement of breath control is produced at some point between 1.6 millions of years and 100.000 years ago. Thus, probably at the time of H. erectus.
7 This is an important observation, since Falk is one of the first in noting the relevance of the laryngeal air sacs in the debate of the evolution of the vocal apparatus. Falk argues that the vertical movement of the hyoid bone “compresses the orifice of the laryngeal air sacs” (p.
H. neanderthalensis Consonants Vocals
Impossible [m], [n], [ɱ], [ɲ], [ɳ], [ŋ], [g], [k]
[u], [i], [a], [ɔ]
Figure 4: Vowels [i], [u], [a], in the human vocal tract (left side) and the repertoire of vowels in human languages (right side)
Thus, the discussion focused on the hyoid bone, in fact quite problematic: It is the only bone of our body which is not in direct touch with another bone, that is to say, it ‘floats’ within tissues. To the uncertain position of this bone, a further problem was added up: They had no sample of a hyoid bone of H. neander-thalensis. The debate did not change its course when a Middle Paleolithic hyoid bone — belonging in all probability to a Neandertal individual — was finally found at Kebara (Israel). Its morphology was similar or even identical to that of H. sapiens (Arensburg et al. 1989, Bar-Yosef et al. 1992). Paleoanthropologists were inclined to conclude that this finding supported a lower larynx and hence, that speech was already present in H. neanderthalensis. It is worth noting that the conclusions ventured from the observation of morphological remains were to some extent hazardous. This great debate about the linguistic capabilities of Neandertals has often been focused in such a way that sometimes it is not easy to discern whether the object of analysis is language or speech. It has been specially Lieberman who has been centered in doing research into Neandertal speech, and not language8 (as a cognitive faculty). Lieberman (1993) has rejected these conclu-sions about the hyoid bone from Kebara, arguing that the shape of the hyoid bone is irrelevant: A pig’s hyoid bone could pass for a human’s; hence, it does not follow that the shape is linked to a supralaryngeal air duct, since this must not to be linked to the hyoid bone. I partially disagree with Lieberman in this respect: The shape is important because, as we will see next, it helps us decide whether the air sacs were connected to the bone or not, and this fact differentiates us from the rest of great apes (gibbons, gorillas, chimpanzees and bonobos; see below section 4.3.).
125) and hence, this fact also participates in the mechanics when the air sacs are emptied and filled up. Chimps lack the mechanisms that prevents humans during deglutition, author adds (above all, because they do not need it at all). Section 2.3.1. of this work is devoted to laryngeal air sacs, whose absence in human beings has been claimed as one of the apomorphies of the genus Homo (see Figure 3).
Figure 5: Models of H. sapiens (male on the left; female in the middle) and H. neanderthalensis (male on right)
Gradually, new morphological elements were introduced which could shed some light on the debate and undo the knot that had tied it down: the basi-cranium angle and the hypoglossal canal. Lieberman (1993) has pointed out the relevance of basicranial flexion, a trait that was related to the supralaryngeal vocal tract, in which larynx is placed down in the neck. An often unnoticed detail is that this does not mean that Neandertals had no capacity for language or speech, but only that these would simply be different from ours (Lieberman & Crelin 1971: 221). This idea was strengthened by arguing that while one half of the human tongue remains in the oral cavity, the second half remains into the pharynx. Daniel Lieberman & McCarthy (1999) ousted the use of the basicranium angle as the ‘key factor’, when they compared the ontogeny of this feature in chimps and humans, and proved that there is no direct relation between the ver-tical growth and the vocal tract.9
In a letter to Science, Lieberman (1999) — against Wolpoff (1998) — dis-agreed with the physical anthropologists’s statements which base the presence of language in Neandertals on their having the hypoglossal canal10 (through which the tongue nerves pass). Lieberman (1999) argued that, the fact that chimps can-not talk tells us can-nothing about the hypoglossal canal11 (smaller in the genus Pan than in the genus Homo).
Thus it seems that — for the moment — the methodology pretending to report an articulated vocal capability based only on fossil morphological traits, has been exhausted. As we have seen, the traditional reconstructions do not let see either (1) whether or not Neandertals had the required superior vocal space, or (2) whether or not the different components (muscles, bones and cartilages) are located properly in the place they had in living individuals, or (3) whether or not Neandertals were able to manage all this bio-mechanic apparatus in order to produce voluntary vocalizations. As far as cognition is concerned, Krantz (1988) had already speculated that Neandertals had to have some neural circuit which made possible the vocalization exhibited by H. sapiens, but he did not add any
9 This has been admitted by Philip Lieberman (2002: 555).
10 In fact, Wolpoff echoed the work made by Kay et al. (1998) on the hypoglossal canal.
detail.12 The great problem is how to measure it in Neandertals, when the best we can get is an endocast of the brain, where one can vaguely detect the brain’s cir-cumvolutions, verify the lateralization and not much more as regards language.13
2.2. New Approaches for Vocalization: The Role of Larynx
In recent years a great technical development has taken place in the field of computational models. This has allowed facing the issue of vocalizations in non-humans from perspectives and techniques which differ from the aforementioned.14 Moreover, the tendency to consider the evolution of modern language lightly practically disappears, so that statements about linguistic capacities based only on the presence of the hyoid bone are not frequent any more (though they are often hinted, especially in paleoanthropology).
This notwithstanding, proposals focused on finding the ‘key factor’ of modern speech are still persistent. Such tendency to avoid the creation of a multi-factorial model which could integrate different proposals brings, time and time again, to a theoretical cul-de-sac. Now the discussion has moved to cavities rather than to bones. Making use of new technologies, Honda & Tiede (1998) proposed a new analysis centered in orofacial morphological factors which correlate with the individual variation of the larynx position, measured in a resting condition. Using orofacial geometry, the authors made a statistical analysis that predicts the height of the larynx. Orofacial geometrical data based on magnetic resonance imaging came from previous works (Honda & Tiede 1998, Tiede, 1998). The anal-yses of correlation showed that the height of the larynx can be predicted either from the aspect ratio of the oral cavity or from the antero-posterior dimension of the facial cranium. A concept taken into account in Honda & Tiede (1998) is that of maximal vowel space.15 Boë et al. (2002) used this parameter to estimate the vowel space in Neandertals. They concluded H. neanderthalensis had a vowel space similar to ours and, hence, could have been able to emit the same range of vowels, though they warned that this does not mean that Neandertals were able to talk. What they affirm is that Neandertals had no morphological limitations in this respect. These results have been criticized by Lieberman (2007), who
12 Lieberman (1992), of course, would disagree with that.
13 This is true regarding the particular cito-architectonic structure that could be involved in Neandertal vocalization. However, there is another kind of information we can get from studies of endocasts and skull shape. For instance, Gunz et al. (2010) have proved that both Neandertal and modern human infants had the same brain development in a first stage, but that the latter differ in a second stage, entering a ‘globuralization phase’ (argued as a derived H. sapiens trait). Thus, it seems that both species had the same brain volume at birth and that “most endocranial shape differences develop postnatally”. The authors speculate “that a shift away from the ancestral pattern of brain development occurring in early Homo sapiens [that is, previous to the Out-of-Africa] underlies brain reorganization and that the associated cognitive differences made this growth pattern a target for positive selection in modern humans.” (Gunz et al. 2010: R-922)
14 See de Boer & Fitch (2010) for a historical and critic summary of the computational models of vocal tracts.
ders that larynx cannot be considered the ‘key factor’ in any case;16 on the contra-ry, the author proposes that the descent and shape of the tongue is what makes possible to H. sapiens the emission of so many different vocalic and consonantal sounds (Lieberman 2007: 556).
Indeed, many authors have paid attention to the descent of larynx. However, sometimes two different debates have been mixed up, when they are essentially different: Having a vowel space like ours does not imply to have a low larynx like ours too. The most important difference between Boë and colleagues, on one side, and Liberman’s arguments, on the other, is that the former pays special attention to the relevance of articulatory control, whereas the latter focuses on anatomy. However, the methodology used by Boë et al. (2002) has been recently criticized by de Boer & Fitch (2010), due to the fact that the model of Boë and his collaborators is based on a previous model — Maeda’s (1990). De Boer & Fitch argue that this model is suitable only for modern human speech. The goal of Maeda’s model was to determine four points: the jaw’s position and the position, shape, and tip of the tongue. De Boer and Fitch also note that most animals do not have an L-shaped vocal tract — something essential, as we have seen — exactly as described in the classical source-filter theory. De Boer & Fitch add that, since Maeda’s model cannot report the irregularities of the tongue, its application to non-humans can lead to wrong inferences regarding the motor abilities of infants or of non-humans, as in the case of Boë and colleagues. Note in passing that the same criticism, hence, would be applicable to Lieberman & Crelin’s (1971) model. Research based in computational models should be en-riched with data from theoretical and field studies, so that the object of study can be outlined with more certainty. Introducing the data from the comparative method is a way to do that, as we will see next.
2.2.1. Data from the Comparative Method: The Larynx Exaptation
Nowadays there is a ‘new’ trend which recovers the concept of exaptation and applies it to the different traits involved in speech evolution. We have seen that the descent of larynx cannot be left aside when one analyzes the human vocal capacity. The fact that a mammal has a low or a high larynx has immediate acoustic consequences. Nonetheless, it is not the ‘key factor’ that made vocalizing possible. On the contrary, it is one change among others — possibly an apomor-phy of the genus Homo — which has been added to the rest.
The descent of larynx has been explained by Lieberman by means of a functionalist theory: The descent of the larynx was beneficial to speech. I disagree with this view, since there are other, more convincing, arguments, concerning a posterior exaptation of this fact. This kind of argumentation has been put forward by Fitch (2002) using the comparative methodology. Fitch argues that other extant non-human species can either lower the larynx momentarily or even maintain it in such a position permanently. Within those that have been proven to exhibit a permanent low larynx, we find two kinds of deers, the Red deer (Cervus elaphus) and the Fallow deer (Dama dama); the males present a permanent
low larynx. This seems to be also true for koalas (Phascolarctos cinereus). Canidae — for example, dogs — can lower the larynx for a moment. In other words, the exhibition of a low larynx is a trait more common in mammals than previously thought. Hence, it should not be so difficult to include Neandertals among them, as Fitch himself has noted.17 The author coincides with Lieberman about the gradual view of such a process, but differs from him regarding the function they could have given to it and thinks that the first hominids could have lowered the larynx momentarily (Fitch 2002: 37). Fitch’s arguments are based on two facts: (1) This trait is evident in other species, whose males can experiment a second descent of the larynx; (2) given that formants can be used to estimate the size of the emitter, the laryngeal descent could have been favored by the ability to recognize whether the opponent is big or small, or even to make believe the adversary that one is bigger than really is.18
Thus, while Lieberman & Crelin concede the descended larynx an adaptive function enhancing an articulatory ability — a progressive enhancement of vocalization in genus Homo —, Fitch, even admitting gradual change of the laryn-geal morphology, rejects that this has been due to linguistic factors (concretely, speech factors. Instead, Fitch believes that the quotidian use of the linguistic vocalization is an exaptation of this trait.
This debate has recently been tackled by means of new models: De Boer (2007) created different computational models and then compared the results with real data.19 The results pointed out that an optimal larynx is that whose values are approximately equivalent to the horizontal dimension of the vocal tract (that is to say, as Lieberman has repeatedly said, that values of the hori-zontal and vertical dimensions are 1:1). The feminine larynx is the one that is closer to the ‘optimal larynx’ — a fact that seems to agree with Lieberman et al.’s (1969) thesis that the vocal tract would have evolved in order to enable as many distinctive articulations as possible. Regarding the second descent of larynx in H. sapiens males, de Boer accepts the theory about the emitter’s size exaggeration (Fitch, 2000) — as the lowest formants point out-, and so the concomitant deeper voice. But he points out that, although this moves males away from the ‘optimal larynx’, in no way supposes a communicative impediment. According to de Boer, the results suggest that, if communication is important for surviving, then the larynx which is placed in the best communicative position will be selected.
I would like to stress that two different issues have been confused during the debate: (1) to have an independent articulatory space — as that proposed by Boë et al. (2002) — is independent of the function a low larynx could have (de Boer 2007); and (2) to have a particular physical trait does not always imply a particular function. Functions can vary in time. The original function of an organ could have nothing to do with the current function. Fitch (2002), as well as Hauser & Fitch (2003), have noted that the physical part gives us only one aspect
17 “If dogs can achieve a substantially lowered larynx, without any changes in basicranial angle or hyoid morphology, it seems likely that neanderthals, other fossil hominids or chim-panzees could as well” (Fitch 2002: 34).
18 This theory is not completely new: Ohala (1984) pointed out that in a lot of human cultures and other non-human species, the F0 is used to seem aggressive and for threatening. 19 The first, a very simple one, gives the author the values of the ‘optimal larynx’; the second
of the trait under analysis — voluntary vocalization with communicative intentions — and hence, the authors affirm that such a change, along with other important changes in the mechanisms of neuronal control, would have smoothed the path for the vocal capacity in H. sapiens. In this regard I agree with them and, as I will argue next, this clearly points in two directions: the neuronal aspect (see sections 4.1 and 4.2) and the genes that favor the neuronal control mechanisms in vocal tasks, like FOXP2 (see section 4.3).
2.2.2. Some Conclusions about Computational Models
The debate about the capabilities for vocalization in Neandertal has been focused until now on morphology, since this was the only secure source of data at first available. Attempts to infer a capacity for oral production counted on little evidences whose argumentation was not difficult to refute. The basicranium angle or the hypoglossal canal alone will never give us more than a trace of how this hominin could have been (traces and signs that must be added up, and not refused, into a global, integrative evolutionary theory of speech). The vocal space in Neandertals seems to be consistent enough in order to produce vocalic sounds, although Boë et al.’s (2002) model is more suitable for H. sapiens. Obviously, some species-specific differences are expected, which could move H. neanderthalensis away — at least qualitatively — from H. sapiens. Not for nothing are we talking about two species. Lieberman’s defense against the idea of conceding Neadertals a modern human-like speech, however fierce, does not discard the possibilities for a positive conclusion about the Neandertal vowel space, as the newly available resources and technology seem to point out (Boë et al. 2002, 2004, Fitch 2002). Hence, it is conceivable a Neandertals superior horizontal vocal tract very similar to that of modern humans, in which the potential vowels would have been very similar — if not exactly the same- to that we produce nowadays. De Boer & Fitch (2010), on the one hand, are pessimistic and think we will never know for sure how the Neandertal vocal tract was. Nonetheless, they admit that, even though mammals cannot produce vowels like humans do, it is also true that they could produce enough phonetic distinctions in order to build a basic oral language.20 In effect, this has been proven in other cetacean species (Zoidis et al. 2008) or even primates — be these monkeys (Ouattara et al. 2009a, b) or great apes like orangutans (Spillmann et al. 2010). Thus, voluntary vocalization in general is not an exclusive matter of humans and the H. Neanderthalensis should be included, according to these criteria, in this set of species. These conclusions are reflected in the following table:
Trait H. neanderthalensis H. sapiens
Low larynx * √
Vocal tract * √
Table 4: Probably Neandertals had a similar laryxn and vocal tract
Obviously, this does not rule out strictly Lieberman’s hypothesis: Neander-tals would not have ‘exactly’ the same vocal tract as humans and hence their vocalizations could perfectly differ from ours. To which extent, it is still not possible to verify. At the same time, Lieberman’s hypothesis does not rule out communication. It is possible that, even in a situation so constrained as the one this author assigns to Neandertals, ‘smooth’ communication could be achieved.
In the next sections I will try to enrich with new pieces of data those aspects that computational models cannot deal with, due to their inherent limitations.21 Regarding this question, let me quote de Boer & Fitch’s (2010) words about the explanatory scope of computational models of vocal tracts:
A real vocal tract cannot make all signals that can be produced by a system consisting of two cylindrical tubes, nor can a system consisting of two cylindrical tubes produce all signals that can be produced by a real vocal
tract. (de Boer & Fitch 2010)
Much the same could be said about other linguistic phenomena like syntax or semantics, in the sense that the models that describe the modern human cannot be good for describing the Neandertal. As a consequence, a strict application of such a view would automatically invalidate any kind of inference, even those from the comparative method, making almost impossible to deepen the hypothesis on what has been said about the extinct hominids or to offer alternative ones. However, I will show in the next sections that it is indeed possible to obtain information from other scientific areas, and that such information can be related to H. neanderthalensis and its potential vocal capability. These sources, in the line pointed by Fitch (2003), are neuropsychology and genetics. Thanks to the comparative method we will see data from other animals that can help us to draw a picture of the Neandertal phonating capability, richer than that offered by works only based on reconstruction or on simulation. I would like to stress that these works do not have to be excluded. On the contrary, they have rather to be complemented by these new data.
Next and finally ending this part on the vocal tract, I will briefly review the historical evolution of an apomorphy of the genus Homo, which distinguishes it from the rest of the Hominidae (see Figure 3): the hyoid bone in relation to the lack of laryngeal air sacs. The fact that these two traits are evolutionary related to each other has had important consequences on the phonating ability of the whole genus Homo.
2.3. The Hyoid Bone in the Genus Homo and the Missing of Air Sacs
As I have advanced (in section 2.1.), we cannot figure out the evolution of voluntary vocalization only from the hyoid bone. However, it is a good pointer of the phylogenetical path followed by the vocal tract: The hyoid bone has implications in the final result of phonation and, this being, the difference in
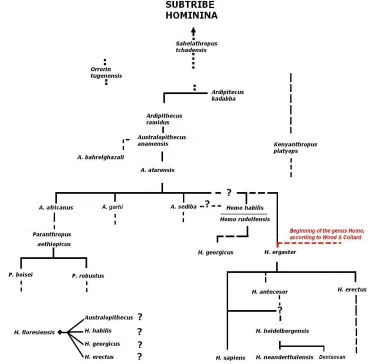
morphology has to produce (minimal) changes. Falk (1975) said that the critical flaw in Lieberman & Crelin’s (1971) results was that the authors had wrongly placed this bone. Thus, the concomitant result was an unproductive vocal tract in comparison with that of H. sapiens. Interestingly, Falk also pointed out the direct relation of this floating bone with other primates’ air sacs of the tribe Hominini, like chimpanzees (Pan troglodytes) and bonobos (Pan paniscus): the hyoid bone compresses the orifice of the air sacs. Thus, one may ask some reasonable questions: in which moment has changed the hyoid bone? Is this a change of the whole genus Homo, or just of the species H. sapiens? Can it be extendable even to the whole subtribe Hominina described in Figure 3?
We find the answer in the fossil record. As said, I assume Wood & Collard’s (1999) proposal according to which the genus Homo has its birth at the African H. erectus clade, also called H. ergaster.22 I’ll show that the hyoid bone is
another trait in favor of this classification.
Additional to the Neandertal hyoid bone from Kebara, two more hyoid bones were found in the Iberian peninsula (Martínez et al. 2008). They date from about 530.000 years and belong to the H. Heidelbergensis, species, considered a precursor of both Neandertals and modern humans (Mounier et al. 2011), though this last issue is more controversial.23 Anyway, if a distribution as the one showed in Figure 2 is accepted, the common ancestor of Neandertals and modern humans already had a modern or derived hyoid bone.
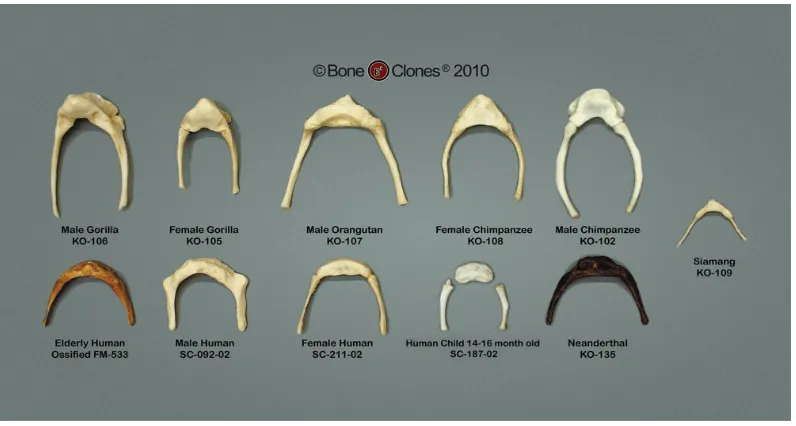
Figure 6: Different hyoid bone models from different primates. © Bone Clones www.boneclones.com, with permission
22 These authors thus transfer H. habilis and H. rudolfensis to the genus Australopitecus (see Figure 3).
However, the most important fossil remain that seems to mark the end of an archaic period of the hyoid bone in the subtribe Hominina is the one found at Dikika, Ethiopia (Alemseged et al. 2006), which belongs to an Australopithecus afarensis. The most remarkable characteristic is that the orifice in the corpus clearly reflects the presence of laryngeal air sacs, a trait absent in H. heidelbergen-sis, H. neanderthalensis and H. sapiens. A morphological analysis of the Australopi-thecus hyoid bone places it in the range of Gorilla gorilla and very close to Pan.24
This floating bone has evolved in many different ways in different species (Figure 6). But its effect on modern acoustics begins with the disappearance of air sacs.
Laryngeal air sacs have been almost unnoticed by scholars. It is Fitch, above all, who has demanded more attention for this apomorphy of the genus Homo (Fitch 2000, Hauser & Fitch 2003): Among great apes, only H. sapiens lacks them. Air sacs have an important effect on vocalizations, producing a sound with its own formant. Campbell’s monkeys use this sound as a particular call apparently linked to a particular meaning (Ouattara et al. 2009a, b).
One could question whether or not these sacs appeared previously to the emergence of our species. Again, I apply the principle of parsimony: There are plenty of mammals exhibiting laryngeal air sacs, like primates, but also cetaceans (Reidenberg & Laitman 2008, Hewitt et al. 2002) which suggests that air sacs are a quite common ancient trait. Within great apes such novelty can be considered an autapomorphy of H. sapiens.25
Apart from that, in relation to the rest of the extinct hominids of the genus Homo that eventually cohabited with H. sapiens, the lack of air sacs is a trait shared with H. neanderthalensis.26
24 If in the future it is shown that the species currently classified as H. habilis had a primitive hyoid bone like that from Dikika, it could be more forcefully argued (1) thar such species is a member of the genus Australopithecus (i.e. Australopithecus habilis); (2) that the derived hyoid bone is an apomorphy of the genus Homo and (3) that all remaining hominids of this lineage (from H. erectus till H. sapiens) shared such morphology, much modern and closer to that shown by H. sapiens. Much the same could be said, of H. floresiensis: a hominid which presents several morphological features that had been lost at the node of H. ergaster. Its inclusion into the genus Homo is due to the fact — as in the H. habilis’ case — that some tools were present in the near of the remains (some of them belonging to the Mode I and some other to the Mode IV), relating them to that species. Its morphology, however, moves it away from the rest of the members of the genus Homo.
25 Note that there exists a kind of laryngeal air sac in H. sapiens, but it has a pathological origin: The laryngeal ventricle of Morgani — a little and elliptical cavity located within the false and true vocal folds — can suffer an abnormal saccular dilatation which could expand outside or inside the larynx. This has been called laryngocele. The increment of air pressure could be its trigger. The most known functional factors are cough, singing or blowing into an instrument (Giovanniello et al. 1970). It is a disease known among trumpet players and glass makers.
26 And probably with another new species: Recently Krause et al. (2010) have published the results of an analysis of DNA of an unknown specimen dubbed the ‘Denisovian hominid’. It is for sure a hominid, and lived 40k ago in the middle of Siberia, at the Denisova cave. Precisely 100 km far away from that place, the Teshik-Tash cave (Uzbekistan), at more or less the same time, was inhabited by Neandertals — as Krause et al.’s (2007) DNA analysis has confirmed.
Experimental studies using computational models have also dealt with this characteristic physical trait and have found that air sacs affect vocalic formants elevating them and thus loosing vocalic space (de Boer 2008, Riede et al. 2008). Although it is necessary to make more research in this field, especially on the functions of air sacs, some conclusions can be drawn. First, the variety of species having this organ suggests us its ancient character. Its absence is a derived trait in extant modern humans, shared with Neandertals. Though it should be confirmed, the evidence points to the possibility that H. erectus already lacked air sacs and also had a derived hyoid bone.
Second, we cannot discard that in some cases it could be a case of new and convergent novelty or homoplasy (see Figure 1).
Third, experiments have stressed the instability effect produced by air sacs and this supports the benefit of maintaining a stable sound source in oral communication, above all in short distances. While air sacs are useful in long distances, as shown by the behavior of Campbell’s monkeys, who create a security perimeter around their living area (Ouattara et al. 2009a, b), or by the long vocalizations of orangutans27 — semi-solitary great apes — whose calls seem to be a distortion source in short distances. This could be the context that would be favored by the sudden disappearance, probably neutral in a first moment, of the air sacs in the genus Homo: close communication within con-specifics.
Lastly, the effect caused by laryngeal air sacs does not indicate that the path covered by the genus Homo was always the ‘improvement path’ of sound production. Air sacs seem to increment the range of vocalizations: Some cercopitecs are known to make use of this organ to emit a particular call (‘boom’). Nevertheless, I agree with Riede et al. (2008) that modern humans are capable of a greater and more refined increment of vocalizations thanks to the multiple combination of articulation points. Note in passing, that this mastery of vocal articulation would put for the moment into the same group the whole genus Homo, from H. erectus till H. sapiens (see Figure 3).
In the next table I show two pieces of data that we unquestionably know regarding the Neandertal man:
Trait H. neanderthalensis H. sapiens
Derived hyoid bone √ √
Lack of air sacs √ √
Table 5: Both species show a derived hyoid bone
that (1) Denisovians were closer to Neandertals than to Sapiens, and (2) that Denisovians interbreed with the Sapiens that reached southeast Asia and Oceania (Reich et al. 2011, Vorobieva et al. 2011). What seems evident is the fact that the diversity and co-existence of hominids in some areas was something usual before the exit of H. sapiens from Africa. 27 Such vocalizations provide information to orangutans helping them to confirm the
3. On the Perceptive Capabilities of H. Neanderthalensis
The discussion carried on until now about the vocal space or over the possibility of emitting articulate sounds by Neandertals makes sense only if this hominid was capable of perceiving and processing them as we do: categorizing them linguistically, in order to reproduce them later on as audible signals. Could the H. neanderthalensis recreate the same sounds, and if so, in the same way as modern humans?
Sound is a phenomenic experience of our cognition: The information from the auditory organs arrives to particular cerebral regions that transform the neural activity into something perceived as sound. The organ of Corti, in the inner ear, contains the structures that allow the conversion of acoustic stimuli into neural activity.
Next, I will deal with aspects of sound processing that are shared by mammals — for example, formants — in such a way that I’ll be able to offer relevant information about the speech processing of Neandertals.
3.1. Formant Perception by H. Neanderthalensis
An aspect of acoustic processing shared with the rest of mammals is formant perception. This observation was formulated in the 70’s by Morton (1977) and recovered by Fitch (2002). Formants let us differentiate between vocal sounds, and in normal conditions only two formants (F1 and F2) are necessary. In 1969 Lieberman and collaborators already suggested that other non-human mammals could be able of perceiving formants. In effect, some years later it has been proven true in rhesus macaque (Macaca mulatta; Hauser et al. 1993) and vervet monkeys (Chlorocebus aethiops; Owren 1990).28 Fitch (2002) holds that formants provide information about the emitter’s body size: Indeed, several studies seem to corroborate a positive correlation of body size and decreased frequencies in humans, domestic dogs and monkeys (Fitch 1997, Fitch & Giedd 1999, Riede & Fitch 1999). These studies suggest that mammals with a larger body produce lower frequencies. According to Fitch (2002), our mammalian ancestors could have used the frequencies of formants in order to estimate the body size, on the basis of the emitter’s vocalizations. Moreover, the author reminds us the fundamental role of formant recognition and the fact that most mammals and current birds can discriminate formants. According to Fitch, this suggests that the reptilian ancestor of both lineages was also capable of doing it. With all this in mind, I see no reason to think that Neandertals were not able to do the same as H. Sapiens and this in a similar way: The Neandertal man’s body was more robust than the anatomically modern human’s, but not so robust as to create a signify-cant difference regarding formant perception.
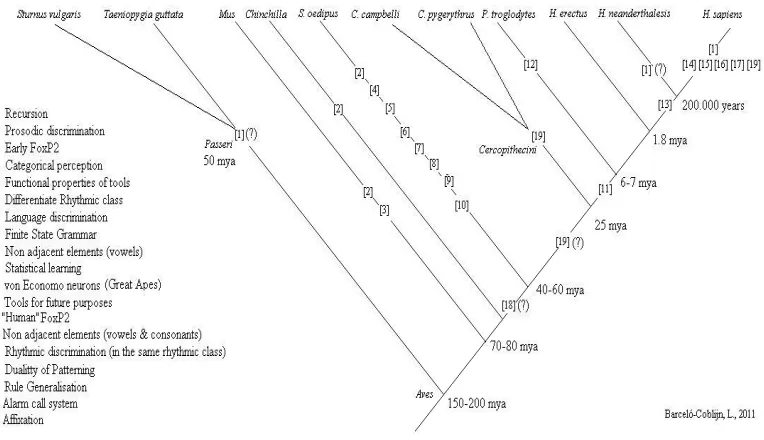
The most basic micro-abilities related to speech perception are theoretically also available to H. neanderthalensis. There are plenty of data from the compara-tive method showing that a lot of percepcompara-tive micro-abilities we link today to language have their origins millions of years ago (see the cognitive cladogram in
Figure 6): The ability to discriminate between different linguistic rhythmic patterns has been positively proven in five-month-old babies (Nazzi et al. 2000), but also in cotton-top tamarins (Saguinus oedipus; Tincoff et al. 2005), and in com-mon mice (Mus; Toro et al. 2003); categorical perception has been put to the test with positive results in chinchillas (Chinchilla; Kuhl & Miller 1975).
Figure 7: Cognitive cladogram showing traits and abilities proved in other species29
Therefore, if one pretends to relate these cognitive micro-abilities to those of modern humans and to postulate a unique origin as well, much the same should be applicable to H. neanderthalensis.
An aspect I cannot assure for the moment (pending improvements in new computational models, or even of new genetic data) is which range of frequencies and which combinations this hominid could produce. In this regard, I coincide with the consideration about the fact that the modern human is the extant mammal which can produce the broadest range of formant frequencies (Lieber-man et al. 1972, 1969).
Hauser & Fitch (2003) have suggested that another kind of information could be extracted from formants: the speaker’s identity. Given the individual differences in the oral and nasal cavities, some subtle differences are produced within speakers, which allow their individual identification. In this sense, it has been suggested that such information could be within the scope of other primates (for example, macaques; Rendall et al. 1998), but Hauser & Fitch (2003) consider that there is still not enough information about it, or verifications that allow us to conclude anything on this point. We know that the recognition of group hierarchy is not an exclusive trait of humans, but also of bonobos and other social
mammals (Seyfarth et al. 2005). Identity recognition by means of formants could be important, depending on the species. Although it is still debated what kind of familiar groups were predominant in the Neandertal society, it is generally agreed that this species could count on a strong social — and possibly also cultural — component (though it would be necessary a less broad definition of the concept of culture). If it is confirmed that this kind of information extraction was already available for hominids before the split between modern humans and Neandertals (as in the case of macaques, mentioned above), it seems that there is no reason to think that a source like that, so useful for social relationships, had fallen from grace in the evolutionary path followed by Neandertals. If the contrary is confirmed, it should be considered the possibility that this is an ability only developed by H. sapiens — but not by the rest of primates.
Ohala (1984) proposed a theory, from an ethological point of view, about the use of the fundamental frequency (F0) in aggressive contexts. According to his observations, it seems that when F0 is lower, the voice acquires a ‘dominant’ edge. Curiously enough, the author pointed out that, in humans, a high F0 is also used to calm an aggressor, trying not to look like a potential threat.30 Thus, the
author implicitly supports the positive interpretation by the hearer. Unfortunately, we are still far from knowing how the formants were produced by Neandertals.
3.2. Looking for the Auditory Bases of H. Neanderthalensis
Genetics has contributed enormously to the understanding of species evolution. In particular, maybe the Neandertal case is one of the most prominent in recent t times, given the narrow bond between both hominids and the endless debates surrounding them (this notwithstanding, Neandertals are not always mentioned in works on genetics, from which straightforward inferences can be drawn). In a recent work, Clark et al. (2003) have detected several genes which have been the target of positive selection during the evolution of mammals — by means of comparison of the genomes of mice, chimps and humans. Regarding the ear and acoustic perception, the authors point to the genes DIAPH1, FOXI1, EYA4 and
OTOR. Some genes of the EYA family31 seem to play a relevant role in the
development of eyes and ears; much the same can be said about FOXI1
(Vidarsson 2007). Most interesting, however, is the fact that the malfunction in the majority of these genes could produce different kinds of deafness, as we can see in Table 6.32
For the moment, it has not been stated explicitly whether Neandertals had exactly the same variants of these genes. Taking into account the great genetic affinity of both species, it would not be surprising that Neandertals had also the same alleles. This notwithstanding, it should also be found out whether their expression was exactly the same for Neandertals. A work compiling the last years
30 Interestingly, Ohala pointed out that this could be a kind of infantile mimicry, and that for
obvious reasons natural selection has left most species with a strong inhibition against infanticide.
31 Whose name comes from ‘Eyes Absent’.
of genetic research on Neandertals has been published recently (Green et al. 2010). In the included list of genes (pp. 714f.), there are all genes that have had a change of amino-acids which are present in current humans, but are ancestral to Neandertals. All of the above mentioned genes are in this provisional list. There also appears the gene EYA2, a gene known to interact with the gene GNAZ. The latter gene has been related to the maintenance of the ionic balance of perilym-phatic and endolymperilym-phatic cochlear fluids. Finally, we find a protein linked to the tectorial membrane,33 ALPHA-TECTORIN (codified by the gene TECTA34) whose
disruption provokes another form of deafness (Verhoeven et al. 1998).
Gene Gene ID Related expression areas Negative repercussions for the ear and other
DIAPH1 1729 Regulation of actin polymerization in hair cells of the inner ear
Autosomal dominant, fully penetrant, nonsyndromic sensorineural progressive low-frequency hearing loss
FOXI1 2299 Kidney, organ of Corti, epydermis, cochlea and vestibulum
Human neurosensorial deafness
EYA4 2070 Organ of Corti Postlingual, progressive,
autosomal dominant hearing loss at the deafness
EYA1 2138 Kidney, eye, ear, branquial arches Branchiootic syndrome35
OTOR 56914 Cartilage development and maintenance
Different forms of deafness
Table 6: Genes related to the inner ear formation and deafness
Altogether, there is firm evidence of the positive selection of some genes related to the ear, to the capacity of auditory perception, and it seems that they have played a fundamental role in the evolution of mammals, in this case in the genus Homo. Note that theirs is not a minor role, in the sense that it affects very basic structures and most of the time they get expressed in different parts of the body (ear, eye, kidney, etc.). Their selection and fixation is not produced suddenly and it would be surprising that Neandertals differed greatly from modern humans regarding these genes. The fact is that, given the high percentage of genetic similarity and the fact that introgressive hybridization36 was
possible (Green et al. 2010), one must contend that the individuals descendant of
33 An extra cellular matrix of the inner ear that contacts the streocilia bundles of specialized sensory hair cells.
34 The Gene IDs of GNAZ and TECTA are 2781 and 7007 respectively. 35 The hearing loss is variable with respect to severity and age of onset.



![Figure 4: Vowels [i], [u], [a], in the human vocal tract (left side) and the repertoire of vowels in](https://thumb-us.123doks.com/thumbv2/123dok_us/8811308.1777612/12.595.115.496.93.255/figure-vowels-human-vocal-tract-left-repertoire-vowels.webp)