KONVERSI ENZIMATIS KULIT PISANG MENJADI GLUKOSA MENGGUNAKAN ENZIM SELULASE DARI Aspergillus niger L-51 YANG
DIAMOBILISASI DENGAN ZEOLIT ALAM (Skripsi)
Oleh RICA AULIA
JURUSAN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
ABSTRACT
ENZYMATIC CONVERSION OF BANANA PEELS TO GLUCOSE USING CELLULASE ENZYME FROM Aspergillus niger L-51
IMMOBILIZED WITH NATURAL ZEOLITE
By
Rica Aulia
Cellulase is a hydrolase enzyme that hydrolyze cellulose in β-1,4-glycoside bonds. Cellulase is widely used in industry to convert cellulose to glucose. In this study, immobilization cellulase enzyme from Aspergillus niger L-51 was carried out using natural zeolite to improve the stability of the enzyme. The cellulase enzyme from the immobilization process is used to convert banana peel to glucose. The stages carried out in this study include: production, isolation, purification, immobilization using natural zeolite, characterization of cellulase enzymes purified before and after immobilization, and enzymatic conversion of banana peels. The results showed that the enzyme specific activity from the purification was 11.560 U/mg and its purity increased 5.2 times compared to the crude extract of cellulase enzyme with a specific activity 2.194 U/mg. The purified enzyme has an optimum temperature at 60oC; KM = 34.683 mg/mL
substrate; Vmax = 52.631 μmol/mL minute, while the immobilized enzyme has an
optimum temperature at 70oC; KM = 0.606 mg/mL substrate; Vmax = 0.545 μmol/mL
minutes. The residual activity of purified and immobilized enzymes in the thermal stability test was 18 and 34%, respectively. The results of purification of enzyme kinetics obtained by ki value = 0.016 1/minute; ∆Gi = 76.695 kJ/mol, and t1/2 = 43.310
minutes, while the immobilization enzyme obtained ki value = 0.009 1/minute; ∆Gi =
78.240 kJ/mol, and t1/2 = 77 minutes. Kivalues, t1/2, and∆Gi showed that immobilized
enzyme was more stable than purified enzyme so it can be used to stage hydrolysis.
Hydrolysis of banana peel with immobilized cellulase enzyme for 24 hours produced the highest glucose level, which was 11.524 mg/mL.
ABSTRAK
KONVERSI ENZIMATIS KULIT PISANG MENJADI GLUKOSA MENGGUNAKAN ENZIM SELULASE DARI Aspergillus niger L-51 YANG
DIAMOBILISASI DENGAN ZEOLIT ALAM
Oleh Rica Aulia
Selulase adalah enzim hidrolase yang menghidrolisis selulosa pada ikatan β-1,4-glikosida. Selulase banyak digunakan dalam industri untuk mengkonversi selulosa menjadi glukosa. Pada penelitian ini telah dilakukan amobilisasi enzim selulase dari
Aspergillus niger L-51 menggunakan zeolit alam untuk meningkatkan stabilitas enzim
tersebut. Enzim selulase hasil amobilisasi digunakan untuk mengkonversi kulit pisang menjadi glukosa. Adapun tahapan yang dilakukan dalam penelitian ini meliputi: produksi, isolasi, pemurnian, amobilisasi menggunakan zeolit alam, karakterisasi enzim selulase hasil pemurnian sebelum dan setelah amobilisasi, dan konversi kulit pisang secara enzimatis. Hasil penelitian menunjukkan aktivitas spesifik enzim hasil pemurnian sebesar 11,560 U/mg dan kemurniannya meningkat 5,2 kali dibandingkan ekstrak kasar enzim enzim selulase yang mempunyai aktivitas spesifik sebesar 2,194. Enzim hasil pemurnian memiliki suhu optimum 60oC; KM= 34,683 mg/mL substrat; Vmaks =
52,631μmol/ mL.menit, sedangkan enzim hasil amobilisasi memiliki suhu optimum 70oC; KM= 0,606 mg/mL substrat; Vmaks = 0,545 μmol/ mL.menit. Aktivitas sisa dari
enzim hasil pemurnian dan enzim hasil amobilisasi pada uji stabilitas termal berturut-turut sebesar 18 dan 34%. Data kinetika enzim hasil pemurnian diperoleh nilai ki =
0,016 menit-1;∆Gi= 76,695 kJ/mol, dan t1/2= 43,31 menit, sedangkan enzim amobilisasi
diperoleh nilai ki = 0,009 menit-1;∆Gi = 78,240 kJ/mol, dan t1/2 = 77 menit. Nilai ki, t1/2
dan ∆Gi yang diperoleh menunjukkan bahwa enzim hasil amobilisasi lebih stabil dibandingkan dengan enzim hasil pemurnian sehingga digunakan untuk tahap hidrolisis. Hidrolisis kulit pisang kepok dengan enzim selulase amobil selama 24 jam menghasilkan kadar glukosa tertinggi, yaitu 11,524 mg/mL.
Kata kunci : selulase, Aspergillus niger L-51, amobilisasi, zeolit alam, kulit pisang,
KONVERSI ENZIMATIS KULIT PISANG MENJADI GLUKOSA MENGGUNAKAN ENZIM SELULASE DARI Aspergillus niger L-51
YANG DIAMOBILISASI DENGAN ZEOLIT ALAM
Oleh RICA AULIA
Skripsi
Sebagai Salah Satu Syarat Untuk Mencapai Gelar SARJANA SAINS
Pada
Jurusan Kimia
Fakultas Matematika dan Ilmu Pengetahuan Alam
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
RIWAYAT HIDUP
Penulis dilahirkan di Bandar Lampung, pada tanggal
28 Januari 1997, sebagai anak kedua dari tiga
bersaudara yang Penulis merupakan putri dari
pasangan Bapak Yulianto dan Ibu Junariah.
Penulis mengawali jenjang pendidikan dari Taman
Kanak- Kanak Aisyah Bandar Lampung pada tahun 2001–2002.
Kemudian melanjutkan pendidikan Sekolah Dasar (SD) di SD Al-Azhar 2
Bandar Lampung hingga tahun 2008. Pendidikan Sekolah Menengah
Pertama (SMP) diselesaikan pada tahun 2011 di SMP Negeri 4 Bandar
Lampung. Pendidikan Sekolah Menengah Atas (SMA) diselesaikan di
SMA YP UNILA Bandar Lampung pada tahun 2014 dan pada tahun yang
sama penulis diterima sebagai Mahasiswa Jurusan Kimia Fakultas
Matematika dan Ilmu Pengetahuan AlamUniversitas Lampung melalui
jalur UMPTN (Ujian Mandiri Perguruan Tinggi Negeri).
Selama menjadi mahasiswa, penulis aktif di Lembaga Kemahasiswaan
Himpunan Mahasiswa Kimia (HIMAKI) FMIPA Universitas Lampung
sebagai anggota Sosial Masyarakat (SOSMAS) pada tahun 2015-2016.
Penulis juga pernah menjadi asisten praktikum Biokimia untuk Jurusan
Kimia FMIPAUniversitas Lampung pada tahun2017. Pada bulan
Juli-Agustus 2017 penulis melaksanakan Kuliah Kerja Nyata (KKN) di Desa
MOTTO
“ La Tahla”
(Hei, Jangan Mengeluh! )
“ Allah tidak membebani seseorang itu melainkan
sesuai dengan kesanggupannya.”
( Al Baqarah (2) : 286 )
“Apabila Kita Ridho Pada Sesuatu Yang Mengecewakan Hati
Kita, Maka Percayalah Allah Akan Menggantikan
Kekecewaan Itu Dengan Sesuatu Yang Tidak Kita Sangka”
Jangan pernah menunggu waktu yang tepat untuk
melakukan sesuatu, karena sesuatu datang di waktu yang
tidak kita duga.
Sesungguhnya Keberhasilan Yang Kita Gapai Merupakan
Doa Orang Tua Yang Diridhoi Allah SWT
“Jangan terdiam teruslah bergerak, karena dunia tidak akan
pernah menunggu mu”
Alhamdulillah Puji Syukur Atas Kehadirat Allah SWT yang Maha Pengasih dan Maha Penyayang
Kupersembahkan karya sederhanaku ini sebagai wujud sayang, bakti, dan tanggung jawab kepada:
Kedua orang tuaku yang telah merawatku dengan penuh kasih sayang, yang selalu memberikan do’a–do’a terbaik, nasihat,
dukungan, serta senantiasa berkorban untukku.
Kakak & adikku yang selalu memberikan dukungan dan semangat.
Para ibu dan bapak dosen yang selama ini telah memberikan banyak ilmu pengetahuan, pelajaran, arahan, serta
bimbingannya kepadaku.
Seluruh sahabat dan teman-teman terdekatku yang selama ini telah memberikan banyak dukungan, bantuan dan motivasi
kepadaku.
Serta Almamaterku, tercinta
SANWACANA
Puji syukur kehadirat Allah SWT atas limpahan rahmat dan karunia-Nya
sehingga penulis dapa tmenyelesaikan penulisan skripsi yang berjudul
“Konversi Enzimatis Kulit Pisang Menjadi Glukosa Menggunakan Enzim Selulase dari Aspergillus niger L-51 yang Diamobilisasi dengan Zeolit Alam”. Skripsi ini adalah salah satu syarat untuk
memperoleh gelar Sarjana Sains pada Jurusan Kimia, Fakultas Matematika
dan Ilmu Pengetahuan Alam,Universitas Lampung.
Penulis menyadari bahwa skripsi ini tidak mungkin terselesaikan tanpa
adanya bimbingan, dorongan, nasihat serta bantuan dari berbagai pihak.
Oleh karena itu penulis menyampaikan terimakasih kepada:
1. Ibu Junariah dan Bapak Yulianto selaku kedua orangtua atas kasih
sayang yang telah diberikan selama ini serta segala doa, dukungan,
nasihat dan motivasinya sehingga penulis dapat menyelesaikan
skripsi ini dengan baik.
2. Bapak Prof. Dr. Ir. Yandri A.S, M.S. selaku Pembimbing I
penelitian dan Pembimbing Akademik atas segala bimbingan,
dukungan dan motivasi serta saran yang diberikan kepada
menyelesaikan skripsi ini.
3. Ibu Dr. Nurhasanah, S.Si., M.Si. selaku Pembimbing II penelitian
atas bimbingan, kritik, saran dan arahannya kepada penulis
sehingga penulis dapat menyelesaikan penelitian dengan baik.
4. IbuProf. Dr. Buhani, M.Si. selaku pembahas penelitian yang
telah memberikan kritik serta saran kepada penulis sehingga
skripsi ini terselesaikan dengan baik.
5. Kakak ku Rico Fambela dan Adikku Muhammad Abdul Wahid yang
selalu memberikan kasih sayang yang tulus, kecerian serta dukungan
untuk keberhasilanku.
6. Seseorang yang selalu menemaniku dalam jatuh dan bangun untuk
tugas akhir ini“Renaldi Arlento”. Terimakasih atas kebahagiannya,
dukunganmu, semangat , nasihatmu, canda dan tawa, keluh kesah,
saran dan kritik yang telah diberikan. Terimakasih sudah menjadi
pendengar yang baik selama ini untukku.
7. Bapak Dr. Eng. Suripto Dwi Yuwono, M.T. selaku Ketua Jurusan
Kimia Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA)
Universitas Lampung.
8. Prof. Warsito, S.Si., D.E.A., Ph.D., selaku Dekan Fakultas Matematika
dan Ilmu Pengetahuan Alam (FMIPA) Universitas Lampung.
9. Bapak Jon Isman selaku laboran Laboratorium Biokimia yang telah
banyak membantu dalam menyediakan alat serta bahan untuk
10. Bapak A.Gani selaku staff loket FMIPA KIMIA yang selalu
membantu memudahkan dan mendukung penulis menyelesaikan
tugas akhir.
11. Seluruh dosen dan staf administrasi di Jurusan Kimia FMIPA
Universitas Lampung yang telah memberikan banyak ilmu
pengetahuan dan motivasi sehingga penulis dapat menyelesaikan
skripsi ini.
12. Erika Liandhini seseorang yang bersamaku selama ini untuk sampai
titik akhir perjuangan perkuliahan. Terimakasih atas pengertianmu,
dukungan, bantuan dan tawamu.
13. Teman seperjuangan penelitan dilab, Riza Mufarida Akhsin, Bunga
Lantri D, Ni Putu , Asrul , Bidari , Leony yang selama ini
menemani dan mendukung penulis dalam segala hal tentang
penelitian.
14. Sahabatku AIB yang memang AIB; Aniza, Ayuning, Hesty, Ana,
Devi, Erien, Pew, Ferita, Ismi, Dira dan Cindy yang selama ini
mendukung dan menemaniku selama masa perkuliahan, makan,
ngelab, ngapa-ngapain. ILoveYouAll~
15. Kakak-kakak, teman-teman dan adik-adik seperjuangan diLab
Biokimia yang selama ini membantu penulis jika ada kesulitan
dilab.
16. Teman-teman seperjuangan Kimia 2014 yang tidak bisa
disebutkan namanya satu persatu. Terimakasih sudah
17. Sahabat masa SMP dan SMA-ku : AyuMon, Aniza, IndriGun, Ipeh,
Mbulqu(Ayuning), Aldha, Tami, Kiky, Imut dan Ungil(Irma) yang
telah memberikan semangat dan motivasi. Thx u All.
18. Semua pihak yang tidak dapatku sebutkan satu persatu yang telah
mendukung dan memotivasi penulis untuk menyelesaikan skripsi
ini dan menyelesaikan studi sebagai mahasiswa S1Kimia.
Akhir kata penulis menyadari bahwa skripsi ini masih kurang dari
kesempurnaan. Penulis berharap semoga skripsi yang sederhana ini dapat
i
DAFTAR ISI
DAFTARTABEL ... iv
DAFTAR GAMBAR ... vi
DAFTAR LAMPIRAN ... viii
I. PENDAHULUAN ... 1
A. Latar Belakang ... 1
B. Tujuan Penelitian ... 4
C. Manfaat Penelitian ... 5
II. TINJAUAN PUSTAKA... 6
A. Enzim ... 6
1. Klasifikasi enzim ... 7
2. Sifat katalitik enzim ... 9
3. Faktor yang mempengaruhi aktivitas enzim ... 9
4. Teori pembentukan enzim substrat ... 13
B. Aspergillus niger ... 14
C. Enzim Selulase... 15
D. Produksi dan Isolasi Enzim Selulase ... 16
E. Pemurnian Enzim Selulase... 17
F. Pengujian Aktivitas Selulase dengan Metode Mandels ... 19
G. Penentuan Kadar Protein dengan Metode Lowry ... 19
H. Zeolit ... 20
I. Amobilisasi ... 23
J. Kinetika Reaksi Enzim ... 25
ii
1. Stabilitas termal enzim... 27
2. Stabilitas pH enzim ... 28
L. Kulit Pisang... 29
M. Hidrolisis Enzimatik ... 30
N. Glukosa ... 32
III. METODE PENELITIAN ... 33
A. Tempat dan Waktu Penelitian ... 33
B. Alat dan Bahan... 33
C. Prosedur Penelitian ... 34
1. Pembuatan media inokulum dan fermentasi ... 34
2. Isolasi enzim selulase... 35
3. Pemurnian enzim selulase... 35
4. Uji aktivitas enzim selulase metode mandels ... 37
5. Penentuan kadar protein metode lowry... 38
6. Amobilisasi enzim selulase hasil pemurnian dengan zeolit... 39
7. Karakterisasi enzim selulase murni dan amobil... 40
8. Konversi enzimatis menjadi glukosa ... 42
IV. HASIL DAN PEMBAHASAN ... 45
A. Enzim Selulase dari Aspergillus niger L-51 ... 45
B. Enzim Selulase Amobil... 48
C. Enzim Selulase Hasil Pemurnian dan Enzim Selulase Amobil ... 50
1. Suhu optimum enzim selulase hasil pemurnian dan enzim selulase amobil ... 50
2. Stabilitas termal enzim selulase hasil pemurnian dan amobil... 51
3. Nilai Km dan Vmaks enzin selulase hasil permurnian dan enzim selulase hasil amobilisasi ... 52
4. Pemakaian berulang enzim amobil ... 54
D. Konstanta Laju Inaktivasi Termal (ki), Waktu Paruh (t1/2), danPerubahan Energi Akibat Denaturasi (∆Gi) Enzim Selulase Hasil Pemurnian dan Hasil Amobilisasi... 55
1. Konstanta laju inaktivasi ... 56
2. Perubahan energi akibat denaturasi (∆Gi)... 56
3. Waktu Paruh... 57
E. Produksi Konversi Enzimatis Kulit Pisang... 57
V. KESIMPULAN DAN SARAN ... 60
iii
B. Saran ... 61
iv
DAFTAR TABEL
Tabel Halaman
1. Pemurnian enzim selulase dari Aspergillus niger L-51 ... 48
2. Nilai ki, t1/2, dan∆Gi enzim selulase hasil pemurnian dan hasil amobilisasi 55 3. Hubungan antara berbagai tingkat kejenuhan ammonium sulfat dengan aktivitas unit enzim selulase ... 69
4. Hubungan antara kejenuhan ammonium sulfat fraksi (0-20%) dan (20-80%) dengan aktivitas spesifik enzim selulase... 69
5. Pengikatan enzim selulase pada matriks (zeolit alam)... 70
6. Hubungan antara suhu (oC) aktivitas enzim selulase hasil pemurnian ... 71
7. Hubungan antara suhu (oC) aktivitas enzim selulase hasil amobilisasi ... 71
8. Hubungan antara aktivitas unit (U/mL) enzim selulase hasil pemurnian selama inaktivasi termal 100 menit... 72
9. Hubungan antara aktivitas unit (U/mL) enzim selulase hasil amobilisasi selama inaktivasi termal 100 menit... 72
10. Data untuk penentuan Km dan Vmaks enzim selulase hasil pemurnian berdasarkan persamaan Lineweaver-Burk ... 73
11. Data untuk penentuan Km dan Vmaks enzim selulase hasil amobilisasi berdasarkan persamaan Lineweaver-Burk ... 73
12. Hubungan antara pengulangan enzim selulase hasil amobilisasi dengan aktivitas unit (U/mL)... 74
v
14. Penentuan nilai ki (konstanta laju inaktivasi termal) enzim hasil amobilisasi pada suhu 50oC... 75
15. Absorbansi glukosa pada berbagai konsentrasi untuk menentukan kurva standar glukosa... 78
16. Hasil konversi enzimatis kulit pisang ... 79
17. Absorbansi serum albumin (BSA) pada berbagai konsentrasi untuk
vi
DAFTAR GAMBAR
Gambar Halaman
1. Hubungan suhu dengan aktivitas enzim ... 10
2. Hubungan pH dengan aktivitas enzim ... 10
3. Hubungan kecepatan reaksi dengan konsentrasi enzim... 11
4. Teori kunci gembok dan teori induksi ... 14
5. Struktur zeolit... 22
6. Struktur Kerangka Zeolit ... 22
7. Kurva Lineweaver-Burk ... 27
8. Mekanisme hidrolisis selulosa dengan enzim ... 32
9. Skema proses fraksinasi enzim dengan ammonium sulfat ... 36
10. Skema singkat mengenai prosedur penelitian ... 44
11. Hubungan antara kejenuhan ammonium sulfat (0-95%) dengan aktivitas unit enzim selulase dari Aspergillus niger L-51... 46
12. Hubungan antara fraksi enzim pada 2 tingkat kejenuhan ammonium sulfat dengan aktivitas unit enzim selulase dari Aspergillus niger L-51 ... 47
13. Hubungan antara aktivitas unit (U/mL) enzim selulase pada berbagai pH pengikatan matriks zeolit alam ... 50
14. Suhu optimum enzim selulase hasil pemurnian dan amobil ... 51
vii
16. Grafik Lineweaver-Burk enzim selulase hasil pemurnian dan amobil ... 53
17. Pemakaian berulang enzim hasil amobilisasi... 54
18. Grafik ln (Ei/E0) enzim selulase hasil pemurnian dan hasil amobil... 56
19. Hasil konversi enzimatis dari kulit pisang ... 58
20. Kurva standar glukosa... 78
viii
DAFTAR LAMPIRAN
Lampiran 1 : Data aktivitas enzim selulase hasil fraksinasi dengan ammonium
sulfat... 69
Lampiran 2 : Data pengikatan enzim selulase pada matriks ... 70
Lampiran 3 : Data aktivitas enzim selulase hasil pemurnian dan hasil amobil pada variasi suhu ... 71
Lampiran 4 : Data aktivitas enzim selulase hasil pemurnian dan hasil amobil selama inaktivasi terml 100 menit ... 72
Lampiran 5 : Data untuk penentuan nilai KMdan Vmaksenzim selulase hasil pemurnian dan hasil amobil ... 73
Lampiran 6: Data aktivitas enzim selulase amobil hasil pengulangan enzim selulase hasil amobil ... 74
Lampiran 7 : Data untuk penentuan nilai ki enzim selulase hasil pemurnian dan hasil amobil pada suhu 50oC... 75
Lampiran 8 : Perhitungan nilai∆Gi dan t1/2enzim selulase hasil pemurnian... 76
Lampiran 9 : Perhitungan nilai∆Gi dan t1/2enzim selulase hasil amobil... 77
Lampiran 10 : Data absorbansi glukosa dan kurva standar glukosa ... 78
ix
Lampiran 12 : Data absorbansi serum albumin (BSA) dan kurva standar albumin (BSA)... 81
I. PENDAHULUAN
A. Latar Belakang
Kulit pisang kepok merupakan limbah pertanian yang belum banyak
dimanfaatkan oleh masyarakat. Kulit dari pisang kepok biasanya oleh masyarakat
hanya dibuang dan hal itu menjadi permasalahan limbah di alam karena akan
meningkatkan keasaman tanah dan mencemarkan lingkungan (Setiawati dkk.,
2013). Minimnya pengetahuan dan informasi mengenai potensi-potensi yang
dimiliki suatu bahan dapat membuat bahan tersebut menjadi tidak berguna dan
tidak memiliki nilai ekonomis. Pembuatan etanol dari limbah selulosa merupakan
rangkaian dari proses pembuatan glukosa, tahapan diawali dengan menghidrolisis
limbah selulosa tersebut sehingga menghasilkan glukosa yang nantinya dapat
digunakan sebagai bahan baku pembuatan etanol. Terdapat beberapa metode pada
proses konversi karbohidrat menjadi gula atau glukosa, di antaranya dengan
hidrolisis asam, secara enzimatis dan menggunakan katalis padat.
Kulit pisang memiliki kandungan selulosa yang tinggi (60-65 %), hemiselulosa
6-8 %, dan lignin 5-10 % (Tjahyono, 1998). Untuk lebih mengoptimalkan
fungsinya, kulit pisang dibuat menjadi bahan yang dapat bermanfaat, salah
2
dapat dipisahkan dan diolah menjadi glukosa dengan proses hidrolisis. Proses
hidrolisis limbah pisang dilakukan dengan cara mereaksikan pati pisang dengan
air berlebih menggunakan katalisator. Menurut Atmaji dkk. (2013) metode
hidrolisis secara enzimatis lebih ramah lingkungan dibandingkan dengan katalis
asam, akan tetapi harganya lebih mahal. Pada penelitian ini hidrolisis secara
enzimatis dengan menggunakan enzim selulase.
Penggunaan enzim selulase tersebut digunakan untuk memecah kulit pisang
kepok menjadi glukosa berdasarkan pada sifat enzim selulase yang dapat
menghidrolisis ikatanβ-1,4-glikosidik pada selulosa, yaitu enzim tersebut bekerja
secara sinergis mendegradasi selulosa dan melepaskan gula reduksi (glukosa)
(Sixta, 2006). Enzim dapat diisolasi dari hewan, tumbuhan dan mikroorganisme.
Namun, secara umum enzim diisolasi dari mikroorganisme karena pertumbuhan
mikroorganisme relatif lebih cepat sehingga enzim yang dihasilkan lebih banyak.
Aspergillus niger adalah spesies yang biomasanya sering digunakan sebagai
adsorben dan dapat dibiakkan dengan baik dalam media agar, ramah lingkungan,
dan bernilai ekonomis. Aspergillus niger adalah salah satu mikroorganisme yang
berkemampuan baik dalam menghasilkan enzim (Frazier and Westhoff, 1981).
Pada umumnya enzim hanya mampu bekerja pada kondisi fisiologis dan tidak
tahan terhadap kondisi ekstrim (Goddette et al., 1993). Untuk mendapatkan
enzim yang stabil dapat melalui proses amobilisasi. Amobilisasi enzim adalah
suatu enzim yang secara fisik maupun kimia tidak bebas bergerak yang masih
mempunyai aktivitas katalitik dan dapat digunakan secara berulang maupun
proses kontinyu (Winarno, 1986). Pada saat digunakan, enzim amobil dapat
3
enzim amobil dapat dipisahkan dari produk dan diperoleh kembali, sehingga
enzim amobil dapat digunakan berulang kali (Darwis dan Sukara, 1990).
Penggunaan enzim amobil hingga saat ini masih banyak digunakan dalam proses
industri karena mempunyai keunggulan tertentu. Menurut Wang et al. (1979)
selain dapat digunakan berulangkali, penggunaan enzim amobil dalam industri
mempunyai beberapa keuntungan lain, di antaranya adalah pemakaian produknya
tidak terkontaminasi oleh enzim, memudahkan proses pengendalian reaksi, dapat
digunakan untuk analisis, dan pada proses amobilisasi tertentu dapat
meningkatkan stabilitas enzim. Amobilisasi enzim dapat dilakukan dengan
metode pengikatan (carrier-binding), metode ikat silang (cross linking), dan
metode penjebakan (entrapment). Salah satu metode yang paling sederhana
adalah dengan metode pengikatan (carrier-binding) secara adsorpsi fisik pada
suatu padatan pendukung. Metode adsorpsi fisik memiliki kelebihan, yaitu
aktivitas dari enzim tetap tinggi (tidak terjadi perubahan konformasi enzim) dan
media dapat diregenerasi (Susanto dkk., 2003).
Padatan pendukung yang digunakan pada penelitian ini adalah zeolit alam. Zeolit
alam digunakan sebagai senyawa pengamobilisasi enzim karena sifat yang
dimiliki oleh zeolit memungkinkan untuk dimodifikasi menjadi katalis, adsorben,
dan sebagai matrik pengamobil, sehingga dapat digunakan untuk meningkatkan
stabilitas enzim dan mendapatkan kondisi optimum enzim selulase bebas dan
amobil, yang meliputi pH optimum, suhu optimum, dan stabilitas termal (Sutarti
dan Rachmawati, 1994). Pemanfaatan zeolit sebagai media pendukung
4
dari Aspergillus niger dan terbukti dapat meningkatkan stabilitas enzim selulase.
Enzim hasil pemurnian bekerja optimum pada suhu 50ºC, sedangkan enzim
amobil pada suhu 75ºC. Aktivitas spesifik enzim hasil pemurnian diperoleh
sebesar 38, 029 U/mg, meningkat 7 kali dibandingkan ekstrak kasar enzim yaitu
5,394 U/mg (Oktariani, 2018).
Berdasarkan latar belakang dan permasalahan yang diuraikan, maka pada
penelitian ini akan dilakukan amobilisasi enzim selulase dari Aspergillus niger
L-51 dan menggunakan enzim amobil tersebut untuk mengkonversi kulit pisang
menjadi glukosa.
B. Tujuan Penelitian
Tujuan dari penelitian ini adalah sebagai berikut :
1. Memperoleh enzim selulase dari Aspergillus niger L-51 dengan aktivitas dan
tingkat kemurnian yang tinggi.
2. Meningkatkan kestabilan enzim selulase dari Aspergillus niger L-51 melalui amobilisasi menggunakan zeolit alam.
5
C. Manfaat Penelitian
Hasil penelitian ini diharapkan dapat digunakan sebagai dasar untuk
mengoptimalkan potensi kulit pisang untuk produksi gula yang dihidrolisis oleh
II. TINJAUAN PUSTAKA
A. Enzim
Enzim adalah biokatalisator yang sangat efektif yang dapat meningkatkan
kecepatan reaksi kimia spesifik secara nyata, suatu reaksi tanpa enzim akan
berlangsung lambat (Lehninger, 2005). Suatu enzim dapat mempercepat laju
reaksi kira-kira 108sampai 1011kali lebih cepat dibandingkan dengan reaksi yang
tidak menggunakan katalisis (Poedjiadi, 1994). Molekul enzim biasanya
berbentuk bulat (globular), sebagian besar terdiri lebih dari satu polipeptida
(Wirahadikusumah, 1997) dan umumnya mempunyai berat molekul yang
beraneka ragam berkisar 104-107kDa (Dryer dan Montgomery, 1993).
Mikroorganisme, terutama ragi, telah banyak digunakan selama beberapa ribu
tahun untuk membuat bir, minuman anggur dan beberapa produk lainnya.
Namun, baru pada tahun 1878 oleh Kuhne, komponen sel ragi yang bertanggung
jawab terhadap fermentasi disebut sebagai enzim. Kurang dari dua dasawarsa
berikutnya, sifat enzim yang tidak hidup dibuktikan secara jelas dengan
menggunakan ekstrak ragi yang bebas sel dan ternyata ekstrak tersebut mampu
7
Enzim bekerja dengan cara menempel pada permukaan molekul zat-zat yang
bereaksi sehingga mempercepat proses reaksi. Percepatan reaksi terjadi karena
enzim menurunkan energi pengaktifan yang dengan sendirinya akan
mempermudah terjadinya reaksi. Enzim mengikat molekul substrat membentuk
kompleks enzim substrat yang bersifat sementara kemudian terurai membentuk
enzim bebas dan produknya (Lehninger, 2005).
Enzim banyak digunakan di berbagai sektor industri, terutama industri makanan.
Selain itu, enzim juga dapat digunakan dalam industri deterjen, farmasi, dan
tekstil. Lebih dari 2000 enzim yang telah diisolasi, hanya 14 enzim yang
diproduksi secara komersial. Kebanyakan dari enzim ini adalah hidrolase,
misalnya amilase, protease, pektinase dan selulase. Enzim penting lainnya adalah
glukosa isomerase dan glukosa oksidase. Alasan digunakannya enzim dalam
industri karena enzim mempunyai kelebihan (Flower, 1998) antara lain :
1. Kemampuan katalitik yang tinggi mencapai 109-1012kali laju reaksi
nonaktivitas enzim.
2. Spesifikasi substrat yang tinggi.
3. Reaksi dapat dilakukan pada kondisi yang lunak, yaitu pada tekanan dan
temperatur rendah.
1. Klasifikasi enzim
Klasifikasi enzim dapat dibedakan sebagai berikut :
a. Berdasarkan tempat bekerjanya enzim dibedakan menjadi dua, yaitu:
1. Endoenzim, disebut juga enzim intraseluler, yaitu enzim yang bekerja
8
2. Eksoenzim, disebut juga enzim ekstraseluler, yaitu enzim yang
bekerja di luar sel.
b. Berdasarkan fungsinya enzim dapat dibedakan menjadi enam kelas dan
tiap kelas mempunyai beberapa subkelas. Pada tiap subkelas, nama resmi
dan nomor klasifikasi dari tiap enzim mengindikasikan reaksi yang
dikatalisis berdasarkan IUPAC yaitu:
1. Oksidoreduktase, yaitu enzim yang mengkatalisis reaksi
oksidasi-reduksi, yang merupakan pemindahan elektron, hidrogen atau
oksigen.
2. Transferase, yaitu enzim yang mengkatalisis perpindahan gugus
molekul dari suatu molekul ke molekul yang lain, seperti gugus
amino, karbonil, metal, asil, glikolis atau fosforil.
3. Hidrolase, yaitu enzim yang mengkatalisis pemutusan ikatan antara
karbon dengan berbagai atom lain dengan adanya penambahan air.
4. Liase, yaitu enzim yang mengkatalisis penambahan gugus fungsi dari
suatu molekul tanpa melalui proses hidrolisis.
5. Isomerase, yaitu enzim yang mengkatalisis reaksi isomerasi.
6. Ligase, yaitu enzim yang mengkatalisis reaksi penggabungan dua
molekul dengan dibebaskannya molekul pirofosfat dari nukleosida
trifosfat.
c. Berdasarkan cara terbentuknya enzim dibedakan menjadi dua, yaitu:
1. Enzim konstitutif, yaitu enzim yang jumlahnya dipengaruhi kadar
9
2. Enzim adaptif, yaitu enzim yang pembentukannya dirangsang oleh
adanya substrat, contohnya enzim β-galaktosidase yang dihasilkan
oleh bakteri E. coli yang ditumbuhkan di dalam medium yang
mengandung laktosa (Lehninger, 1982).
2. Sifat katalitik enzim
Sifat-sifat katalitik dari enzim ialah sebagai berikut :
a. Enzim mampu meningkatkan laju reaksi pada kondisi biasa (fisiologis)
dari tekanan, suhu, dan pH.
b. Enzim berfungsi sebagai selektifitas tinggi terhadap substrat (substansi
yang mengalami perubahan kimia setelah bercampur dengan enzim) dan
jenis reaksi yang dikatalisis.
c. Enzim memberikan peningkatan laju reaksi yang tinggi dibanding
dengan katalis biasa (Page, 1989).
3. Faktor yang mempengaruhi aktivitas enzim
Beberapa faktor yang mempengaruhi aktivitas enzim sebagai berikut :
a. Suhu
Enzim dapat mempercepat terjadinya reaksi kimia pada suatu sel hidup.
Dalam batas-batas suhu tertentu, kecepatan reaksi yang dikatalisis enzim
akan naik bila suhunya naik. Reaksi yang paling cepat terjadi yaitu pada
suhu optimum (Rodwell, 1987). Suhu yang terlalu tinggi akan
menyebabkan enzim terdenaturasi (Poedjiadi, 1994). Pada suhu 0oC
10
(Lay dan Sugyo, 1992). Hubungan antara aktivitas enzim dengan suhu
ditunjukkan dalam Gambar 1.
Aktivitas Enzim
[image:32.595.182.400.158.263.2]Suhu
Gambar 1. Hubungan suhu dengan aktivitas enzim (Poedjiadi, 1994).
b. pH
Enzim bersifat amfolitik, yang berarti enzim mempunyai konstanta
disosiasi pada gugus asam maupun gugus basanya, terutama pada gugus
residu terminal karboksil dan gugus terminal aminonya, diperkirakan
perubahan kereaktifan enzim akibat perubahan pH lingkungan (Winarno,
[image:32.595.186.381.533.654.2]1986). Hubungan kecepatan reaksi dengan pH ditunjukkan pada
Gambar 2.
11
c. Konsentrasi enzim
Konsentrasi enzim secara langsung mempengaruhi kecepatan laju reaksi
enzimatik, laju reaksi meningkat seiring dengan bertambahnya
konsentrasi enzim (Poedjiadi, 1994). Laju reaksi tersebut meningkat
secara linier selama konsentrasi enzim jauh lebih sedikit daripada
konsentrasi substrat, hal ini biasanya terjadi pada kondisi fisiologis (Page,
1997). Hubungan antara kecepatan reaksi dengan konsentrasi enzim
ditunjukkan dalam Gambar 3.
Kecepatan reaksi
[image:33.595.188.410.334.451.2]Konsentrasi enzim
Gambar 3. Hubungan kecepatan reaksi dengan konsentrasi enzim (Page,
1997).
d. Konsentrasi substrat
Kecepatan reaksi enzimatis pada umumnya tergantung pada konsentrasi
substrat. Kecepatan reaksi akan meningkat apabila konsentrasi substrat
meningkat. Peningkatan kecepatan reaksi ini akan semakin kecil hingga
mencapai suatu titik batas yang pada akhirnya penambahan konsentrasi
substrat hanya akan sedikit meningkatkan kecepatan reaksi (Lehninger,
12
e. Pengaruh kadar air
Kadar air dari bahan sangat mempengaruhi laju reaksi enzimatik. Kadar
air bebas yang rendah mengakibatkan difusi enzim atau substrat,
akibatnya hidrolisis hanya akan terjadi pada bagian substrat yang
langsung berhubungan dengan enzim. Misalnya pada kadar air 20 % atau
kira-kira bahan mengandung 4 % air bebas, amilase hanya menghasilkan
produk hidrolisis glukosa dan maltosa. Pada kadar air yang lebih tinggi,
selain glukosa dan maltosa terbentuk juga dekstrin (Winarno, 1986).
f. Pengaruh kadar garam
Kadar elektrolit yang tinggi umumnya mempengaruhi kelarutan protein,
karena itu sering digunakan untuk melarutkan beberapa jenis protein.
Peristiwa ini disebut dengan salting in. Sebaliknya beberapa jenis larutan
garam lain dapat digunakan untuk membuat protein enzim menjadi tidak
larut. Proses ini disebut dengan istilah salting out, yang dapat
dimanfaatkan untuk mengisolasi enzim. Garam ammonium sulfat sering
digunakan untuk fraksinasi dan isolasi enzim karena sifat kelarutannya
dalam air yang tinggi dan tidak mengganggu bentuk dan fungsi enzim
(Scopes, 2002).
g. Aktivator dan inhibitor
Beberapa enzim memerlukan aktivator dalam reaksi katalisnya. Aktivator
adalah senyawa atau ion yang dapat meningkatkan kecepatan reaksi
13
kofaktor. Kofaktor tersebut dapat berupa ion-ion anorganik seperti Zn,
Fe, Ca, Mn, Cu atau Mg atau dapat pula sebagai molekul organik
kompleks yang disebut koenzim (Martoharsono, 1984).
Inhibitor merupakan suatu zat kimia tertentu yang dapat menghambat
aktivitas enzim (Wirahadikusumah, 1997). Pada umumnya cara kerja
inhibitor adalah dengan menyerang sisi aktif enzim sehingga enzim tidak
dapat berikatan dengan substrat dan fungsi katalitik enzim tersebut akan
terganggu (Winarno, 1986).
h. Kadar protein enzim
Menurut Scopes (2002) kandungan protein di dalam enzim sangat
berpengaruh terhadap daya katalitik enzim tersebut. Pada umumnya
dengan meningkatnya kadar protein dalam suatu enzim, maka daya
katalitiknya akan meningkat. Salah satu metode dapat digunakan dalam
menentukan kadar protein adalah metode Lowry.
4. Teori pembentukan enzim-substrat
Cara kerja enzim (Page, 1997) dapat dijelaskan dengan dua teori, yaitu teori
kunci gembok (lock and key theory) dan teori kecocokan yang terinduksi
(induced fit theory), ditunjukkan dalam Gambar 4. Menurut teori
kunci-gembok, enzim dan substrat bergabung bersama membentuk kompleks,
seperti kunci yang masuk dalam gembok, hal ini dikarenakan adanya
kesesuaian bentuk ruang antara substrat dengan sisi aktif enzim, sehingga sisi
14
dengan energi aktivasi yang rendah. Setelah bereaksi, kompleks lepas dan
melepaskan produk serta membebaskan enzim. Sedangkan menurut teori
kecocokan yang terinduksi, sisi aktif enzim merupakan bentuk yang fleksibel.
Ketika substrat memasuki sisi aktif enzim, bentuk sisi aktif termodifikasi
melingkupi substrat membentuk kompleks. Ketika produk dihasilkan, maka
enzim akan dilepaskan dalam bentuk bebas dan dapat bereaksi kembali
[image:36.595.178.472.291.399.2]dengan substrat yang baru.
Gambar 4. Teori kunci-gembok dan teori induksi (Shahib, 2005).
B. Aspergillus niger
Aspergillus niger merupakan fungi dari ascomycota yang berfilamen, mempunyai
hifa, bercabang-cabang dan bersekat, berwarna terang atau tidak berwarna, dan
ditemukan melimpah di alam (Esser, 2002). Aspergillus niger dapat tumbuh
optimum pada suhu 30-37oC, pH 4-6 dengan suhu minimum 6-8oC dan suhu
maksimal 45-47oC, dan bersifat aerob atau memerlukan oksigen pada proses
pertumbuhannya. Jamur ini memiliki warna dasar putih atau kuning dengan
lapisan konidiospora tebal berwarna coklat gelap sampai hitam. Jamur ini
15
Koloninya berwarna putih pada agar dekstrosa kentang (PDA) 25oC dan berubah
menjadi hitam ketika konidia dibentuk. Kepala konidia dari Aspergillus niger
berwarna hitam, bulat, cenderung memisah menjadi bagian-bagian yang lebih
longgar seiring dengan bertambahnya umur (Rao, 1998).
Aspergillus niger dapat tumbuh dengan cepat sehingga banyak digunakan secara
komersial dalam produksi asam sitrat, asam glukonat, dan pembuatan beberapa
enzim seperti amilase, peptinase amiloglukosida, dan selulase (Broekhuijsen et
al., 1993; Okada, 1985). Aspergillus niger mampu mensintesis asam sitrat dalam
medium fermentasi ekstraseluler dengan konsentrasi yang cukup tinggi, jika
dibiakkan dalam media yang kadar garamnya rendah dan mengandung gula
sebagai sumber karbon (Hang et al., 1977; Ji et al.,1992).
C. Enzim Selulase
Enzim yang dapat menghidrolisis ikatan β-1,4-glikosidik pada selulosa adalah
selulase. Selulase adalah enzim terinduksi yang disintesis oleh mikroorganisme
selama ditumbuhkan dalam medium selulosa (Lee, 2001). Selulase termasuk
sistem multienzim yang terdiri dari tiga komponen. Untuk menghidrolisis
selulosa yang tidak larut atau selulosa kristal diperlukan kerja sinergik dari ketiga
komponen enzim tersebut.
Menurut Reese (1976) adapun ketiga komponen enzim tersebut, yaitu:
1. Ekso-β-(1,4)-glukanase dikenal sebagai faktor C1. Faktor ini diperlukan
16
2. Endo-β-(1,4)-glukanase dikenal sebagai faktor Cx. Faktor ini diperlukan
untuk menghidrolisis ikatan β-(1,4)-glukosida (selulosa amorf)
3. Β-(1,4)-glukosidase menghidrolisis selobiosa menjadi glukosa.
Aktivitas selulase disebabkan oleh enzim non hidrolitik C1, hidrolisis selulosa
yang telah diaktifkan dilakukan oleh enzim Cx. Menurut hipotesa ini, mikroba
yang tumbuh pada selulosa yang telah dilonggarkan oleh asam fosfat atau selulosa
tersubtitusi akan kekurangan enzim C1, tetapi banyak menghasilkan enzim Cx
(Muchtadi et al., 1992).
Selulase dapat diproduksi oleh fungi, bakteri, dan ruminansia. Produksi enzim
secara komersial biasanya menggunakan fungi atau bakteri. Fungi yang bisa
menghasilkan selulase antara lain dari genus Tricoderma, Aspergillus, dan
Penicillium, sementara bakteri penghasil selulase antara lain Acidothermus,
Bacillus, Clostridium, Pseudomonas dan Rhodothermus (Sukumaran et al., 2005).
D. Produksi dan Isolasi Enzim Selulase
Enzim dapat diisolasi secara ekstraseluler dan intraseluler. Enzim ekstraseluler
merupakan enzim yang bekerja diluar sel, sedangkan enzim intraseluler
merupakan enzim yang bekerja di dalam sel. Ekstraksi enzim ektraseluler lebih
mudah dibandingkan ekstraksi enzim intraseluler, karena tidak memerlukan
pemecahan sel dan enzim yang dikeluarkan dari sel mudah dipisahkan dari
pengotor lain serta tidak banyak bercampur dengan bahan-bahan sel lain (Pelczar,
17
Isolasi enzim selulase dapat dilakukan dengan metode sentrifugasi. Metode ini
digunakan untuk memisahkan enzim ekstraseluler dari sisa-sisa sel. Sentrifugasi
akan menghasilkan supernatan yang jernih dan endapan yang terikat kuat pada
dasar tabung, yang kemudian dipisahkan secara normal. Sel-sel mikroba biasanya
mengalami sedimentasi pada kecepatan 5000 selama 15 menit (Scopes, 1982).
Prinsip sentrifugasi berdasarkan pada kenyataan bahwa setiap partikel yang
berputar pada laju sudut yang konstan akan memperoleh gaya keluar (F). Besar
gaya ini bergantung pada laju sudut ω (radian/detik) dana radius pertukarannya
(sentimeter) (Cooper, 1997).
E. Pemurnian Enzim Selulase
Setelah enzim selulase diisolasi, selanjutnya enzim tersebut dimurnikan
menggunakan metode fraksinasi dengan menggunakan ammonium sulfat
(NH4)2SO4dan dialisis.
1. Fraksinasi
Fraksinasi merupakan proses pengendapan protein atau enzim dengan
penambahan senyawa elektrolit seperti garam ammonium sulfat, natrium
klorida atau natrium sulfat. Pada umumnya garam yang sering digunakan
adalah ammonium sulfat karena (1) kebanyakan enzim tahan terhadap garam
ini, (2) memiliki kelarutan yang besar dalam air, (3) mempunyai daya
pengendapan yang besar, dan mempunyai efek penstabil terhadap kebanyakan
18
Suhartono (1989), penambahan senyawa elektrolit menurunkan kelarutan
protein, karena kelarutannya dipengaruhi oleh kekuatan ion. Dengan
meningkatnya kekuatan ion, kelarutan enzim akan semakin besar atau disebut
dengan peristiwa salting in, setelah mencapai suatu titik tertentu kelarutannya
akan semakin menurun atau disebut peristiwa salting out. Pada kekuatan ion
rendah, protein akan terionisasi sehingga interaksi antar protein akan
menurun dan kelarutan akan meningkat. Peningkatan kekuatan ion ini
meningkatkan kadar air yang terikat pada ion, dan jika interaksi antar ion
kuat, kelarutannya menurun akibatnya interaksi antar protein lebih kuat dan
kelarutannya menurun (Agustien dan Munir, 1997).
2. Dialisis
Dialisis adalah suatu metode yang digunakan untuk memisahkan garam dan
larutan protein atau enzim berdasarkan prinsip membran semipermeabel
akibat adanya proses fraksinasi menggunakan garam pada tahap pemurnian
larutan protein atau enzim. Secara umum proses dialisis adalah dengan
memasukkan enzim atau larutan protein hasil fraksinasi kedalam kantong
dialisis yang bersifat membran semipermeabel (kantong selofan). Jika
kantong yang berisi larutan enzim dimasukkan ke dalam buffer sambil
diputar-putar, maka molekul kecil yang ada di dalam larutan enzim akan
keluar melewati pori-pori membran, sedangkan molekul besar akan tertahan
dalam kantong dialisis. Kendala ini dapat dilakukan dengan cara mengganti
19
konsentrasi rendah sampai ion-ion dalam kantong dialisis dapat diabaikan
(Lehninger, 1982).
F. Pengujian Aktivitas Selulase dengan Metode Mandels
Pengujian aktivitas selulase dilakukan dengan metode Mandels (Mandels et al.,
1976), yaitu berdasarkan pembentukan glukosa dari substrat Carboxy methyl
Cellulase (CMC) oleh enzim selulase yang dideteksi dengan penambahan
pereaksi Dinitrosalisilic acid (DNS) ke dalam larutan uji serta proses pemanasan,
sehingga akan dihasilkan larutan berwarna kuning hingga merah pekat. Semakin
pekat warna larutan sampel dibandingkan larutan kontrol, maka semakin tinggi
aktivitasnya.
G. Penentuan Kadar Protein dengan Metode Lowry
Penentuan kadar protein bertujuan untuk mengetahui bahwa protein enzim masih
terdapat pada setiap fraksi pemurnian (tidak hilang dalam proses pemurnian)
dengan aktivitas yang baik. Salah satu metode yang digunakan untuk menentukan
kadar protein adalah metode Lowry. Metode ini bekerja pada kondisi alkali dan
ion tembaga (II) yang akan membentuk kompleks dengan protein. Ketika reagen
folin-ciocelteau ditambahkan, maka reagen akan mengikat protein. Ikatan ini
secara perlahan akan mereduksi reagen folin menjadi heteromolybdenum dan
20
Pada metode ini, pengujian kadar protein didasarkan pada pembentukan komplek
Cu2+dengan ikatan peptida yang akan tereduksi menjadi Cu+pada kondisi basa.
Cu+dan rantai samping tirosin, triptofan, dan sistein akan bereaksi dengan reagen
folin-ciocelteau. Reagen ini bereaksi menghasilkan produk yang tidak stabil yang
tereduksi secara lambat menjadi molybdenum atau tungesteen blue. Protein akan
menghasilkan intensitas warna yang berbeda tergantung pada kandungan triptofan
dan tirosinnya.
Metode ini relatif sederhana dan dapat diandalkan serta biayanya relatif murah.
Namun, kekurangan dari metode ini adalah sensitif terhadap perubahan pH dan
konsentrasi protein yang rendah. Untuk mengatasi hal tersebut dapat dilakukan
dengan menggunakan volume sampel dalam jumlah kecil sehingga tidak
mempengaruhi reaksi (Lowry et al., 1951).
H. Zeolit
Zeolit merupakan adsorben yang unik, karena memiliki ukuran pori yang sangat
kecil dan seragam jika dibandingkan dengan adsorben yang lain seperti karbon
aktif dan silika gel, sehingga zeolit hanya mampu menyerap molekul-molekul
yang berdiameter sama atau lebih kecil dari diameter celah rongga, sedangkan
molekul yang diameternya lebih besar dari pori zeolit akan tertahan dan hanya
melintasi antar partikel. Dalam keadaan normal ruang hampa dalam kristal zeolit
terisi oleh molekul air yang berada disekitar kation. Bila zeolit dipanaskan maka
air tersebut akan keluar. Zeolit yang telah dipanaskan dapat berfungsi sebagai
21
Kemampuan zeolit mengadsorpsi sangat bergantung pada rasio Si/Al. Rasio Si/Al
rendah, zeolit bersifat hydrophilic mempunyai afinitas tinggi terhadap air dan
senyawa polar lainnya. Sebaliknya jika rasio Si/Al tinggi, maka zeolit bersifat
hydrophobic dan mengadsorpsi senyawa non-polar (Yuliusman dkk., 2010).
Sifat-sifat zeolit secara langsung berasal dari karakteristik tertentu, dari struktur
kristalnya, sehingga memiliki luas permukaan yang cukup besar. Zeolit dikenal
sebagai adsorben yang selektif dan mempunyai efektifitas adsorpsi yang tinggi
(Newsam, 1991).
Perbedaan jenis zeolit mempunyai daya serap (adsorption) molekul yang
berbeda-beda secara selektif. Keselektifan ini tergantung dari struktur masing-masing
jenis zeolit, sehingga zeolit dapat digunakan sebagai:
1. Penyaring ion, molekul atau sebagai katalis
Zeolit dapat menyaring ion, molekul, maupun atom karena mempunyai
saluran (channel) dan rongga (cavity) dalam struktur zeolit bila oxygen
window dari saluran atau rongga lebih kecil dari ion, molekul atau atom.
Zeolit mempunyai pori sehingga juga dapat digunakan sebagai katalis untuk
mempercepat reaksi dalam proses kimia.
2. Bahan penyerap
Bila zeolit dipanaskan pada suhu tinggi maka akan terjadi dehidrasi,
penguapan yang dikandungnya sehingga menyebabkan zeolit akan selektif
22
Proses penyerapan molekul oleh zeolit terjadi karena strukturnya juga
mempunyai polaritas yang tinggi.
3. Penukar ion
Pertukaran ion pada dasarnya terjadi dalam suatu cairan yang mengandung
anion, kation, dan molekul air. Penukaran ion juga dapat terjadi apabila salah
satu atau sebagian ion terikat pada matriks mikropori bersifat padat. Molekul
air dapat berada dalam mikropori bersama ion (kation, anion) dengan muatan
yang berlawanan dengan ion matriks sehingga terjadi kesetimbangan muatan
untuk mencapai keadaan netral, sehingga ion yang berada dalam cairan dapat
bergerak bebas di dalam matriks mikropori (Ginting dkk., 2007).
Struktur zeolit mengandung saluran atau hubungan rongga yang berisi kation dan
[image:44.595.218.407.467.548.2]molekul air seperti yang tercantum pada Gambar 5.
Gambar 5. Struktur Zeolit (Kahar, 2007)
Struktur kerangka zeolit seperti yang tercantum pada Gambar 6.
[image:44.595.190.434.653.713.2]23
Karakteristik struktur zeolit antara lain :
1. Sangat berpori, karena kristal zeolit merupakan kerangka yang terbentuk dari
jaring tetrahedral SiO4dan AlO4.
2. Pori-porinya berukuran molekul karena pori zeolit terbentuk dari tumpukan
cincin beranggotakan 6, 8, 10 atau 12 tetrahedral.
3. Dapat menukarkan kation, karena perbedaan muatan Al3+dan Si4+
menjadikan atom Al dapat kerangka kristal bermuatan negatif dan
membutuhkan kation penetral. Kation penetral yang bukan menjadi bagian
kerangka ini mudah diganti dengan kation lainnya.
4. Dapat dijadikan padatan yang bersifat asam. Karena penggantian kation
penetral dengan proton-proton menjadikan zeolit padatan asam Bronsted.
5. Mudah dimodifikasi karena setiap tetrahedral dapat dikontakkan dengan
bahan-bahan pemodifikasi (Kahar, 2007).
I. Amobilisasi
Amobilisasi enzim adalah proses pengikatan enzim secara fisik pada suatu
matriks tertentu yang tidak larut dalam air. Keuntungan teknik amobilisasi
diantaranya: dapat meningkatkan stabilitas enzim, memudahkan pengendalian
kondisi reaksi, enzim dapat digunakan berulang, dan kemurnian enzim maupun
produk lebih tinggi. Namun, kekurangan teknik amobilisasi, yaitu terjadinya
penurunan aktivitas katalitik enzim dan terjadinya pergeseran pH atau suhu
24
Amobilisasi dapat diklasifikasikan dalam tiga kategori, yaitu:
1. Metode pengikatan(carrier-binding) yang didasarkan pada pengikatan
enzim dengan carrier atau matriks yang tidak larut dalam air. Aktivitas
enzim amobil dipengaruhi oleh ukuran partikel dan luas permukaan
matriks. Pengikatan dapat dilakukan dengan cara:
a. Adsorpsi fisik yaitu enzim diadsorpsi pada permukaan matriks melalui
ikatan hidrofobik, ikatan hidrogen, dan gaya Van der Waals. Metode ini
mudah dilakukan,ekonomis, tidak merusak konformasi enzim, dan
penurunan aktivitas enzim cenderung rendah. Namun, kekuatan ikatan
antara enzim dan matriks cukup lemah dan rentan terhadap perubahan
pH. Jika pH atau kekuatan ion berubah, maka akan terjadi kebocoran
matriks. Matriks yang dapat digunakan, contohnya: bentonit, silikagel,
zeolit, kitosan, dan alumina. Enzim dan matriks dapat dipisahkan
kembali melalui filtrasi maupun sentrifugasi (Suhartono, 1989).
b. Ikatan kovalen antara gugus fungsi enzim yaitu αatau β-amino; α, β,atau
γ-karboksil; sulfohidril; hidroksil; imidazol; dan fenolik dengan
matriks yang mengandung gugus reaktif seperti diazonium; asam azida;
isosianat; dan halida. Ikatan yang terbentuk cukup kuat dalam mencegah
kebocoran matriks. Namun, jika konformasi berubah maka aktivitas
enzim akan hilang. Matriks yang digunakan pun sulit diregenerasi
c. Ikatan ionik antara gugus karboksil enzim bermuatan negatif dengan
gugus amina suatu matriks bermuatan positif pada matriks yang tidak
25
adsorpsi fisik.
2. Metode ikatan silang (cross-linking) antara molekul enzim dengan pereaksi
bergugus fungsi ganda. Kedua gugus fungsi tersebut akan mengikat
molekul enzim. Pereaksi yang biasa digunakan yaitu glutaraldehid. Untuk
meningkatkan stabilitas enzim, metode ini umumnya dipadukan dengan
metode adsorpsi.
3. Metode penjebakan (entrapment) yaitu penggabungan enzim ke dalam
kisi- kisi gel maupun polimer semipermeabel (mikrokapsul). Matriks gel
yang dapat digunakan, antara lain: poliakrilamida,κ-karagenan, dan
alginat. Polimer yang umum digunakan, yaitu selulosa asetat dan
amilum. Keunggulan metode ini yaitu tidak terjadinya perubahan
konformasi dan inaktivasi enzim karena enzim tidak berikatan dengan
matriks gel. Namun, kemampuan pembentukan kompleks enzim-substrat
cukup rendah apalagi jika berat molekul terlalu besar karena terhalang
kisi gel (Sirisha et al., 2016).
J. Kinetika Reaksi Enzim
Menurut Suhartono (1989) kinetika enzim merupakan salah satu cabang ilmu
enzimologi yang membahas faktor-faktor yang mempengaruhi laju reaksi.
Faktor-faktor yang mempengaruhi laju reaksi seperti; suhu, pH, konsentrasi
enzim, konsentrasi substrat, aktivator, inhibitor, dan waktu inkubasi. Berdasarkan
26
fase yaitu pembentukan kompleks enzim substrat (ES), E adalah enzim dan S
adalah substrat, modifikasi dari substrat membentuk produk (P) yang masih
terikat dengan enzim (EP), dan pelepasan produk dari molekul enzim (Shahib,
2005).
E+S ES EP E+P
Konsentrasi substrat pada laju reaksi divariasikan dan konsentrasi enzim tetap.
Kecepatan awal (V0) diukur sebagai kemiringan targen dalam kurva kemajuan
pada waktu t = 0. Ketika [S] >> [E], maka V0berbanding lurus dengan
konsentrasi enzim dalam campuran reaksi. V0adalah fungsi hiperbola
(persamaan Michaelis-Menten) dari [S]0yang bervariasi.
Ketika [S]0sangat besar maka V0= Vmaks (kecepatan maksimum) dan ketika V0
=Vmaks maka nilai [S]0= Km (tetapan Michaelis).
Persamaan Michaelis-Menten dapat menghasilkan garis lurus ketika satu variabel
baru diplotkan terhadap yang lain. Transformasi persamaan Michaelis-Menten
yang digunakan adalah persamaan“double reciprocal”Lineweaver-Burk.
Dengan mengambil resiprok dari dua sisi persamaan Michaelis-Menten, maka
27
Plot dari pasangan data (1/[S]0,i, 1/V0,i), untuk i = 1, ..., ...,n, dengan n adalah
jumlah pasangan data, akan memberikan suatu garis lurus dengan ordinat dan
[image:49.595.223.406.180.299.2]absis intercept 1/Vmaks dan -1/Km, seperti pada Gambar 7
Gambar 7. Kurva Lineweaver-Burk (Suhartono, 1989).
K. Stabilitas Enzim
Menurut Wiseman (1985) dan Kazan et al. (1997) stabilitas enzim dapat
diartikan sebagai kestabilan aktivitas enzim selama penyimpanan dan penggunaan
enzim tersebut, serta kestabilan terhadap berbagai senyawa yang bersifat merusak
enzim seperti pelarut tertentu (asam atau basa) dan pengaruh suhu dan pH yang
ekstrim atau kondisi-kondisi non fisiologis lainnya. Ada dua cara untuk
meningkatkan stabilitas enzim agar tetap tinggi, yaitu menggunakan enzim yang
memiliki stabilitas enzim alami dan mengusahakan peningkatan stabilitas enzim
yang secara alami tidak atau kurang stabil.
1. Stabilitas termal enzim
Pada suhu yang terlalu rendah kemantapan enzim tinggi, tetapi aktivitasnya
28
kemantapannya rendah. Kenaikkan suhu enzim akan mempengaruhi
kecepatan laju reaksi, namun hanya sampai batas tertentu dan dapat
menyebabkan terjadinya denaturasi protein. Daerah suhu saat kemantapan
dan aktivitas enzim cukup besar disebut suhu optimum untuk enzim tersebut
(Wirahadikusumah, 1997).
Dalam industri, pada proses reaksinya biasanya menggunakan suhu yang
tinggi. Penggunaan suhu yang tinggi bertujuan untuk mengurangi tingkat
kontaminasi dan masalah-masalah viskositas serta meningkatkan laju reaksi.
Penggunaan enzim pada suhu yang lebih tinggi hingga 85-100 °C hanya
dijumpai pada proses hidrolisis pati dengan menggunakan α-amilase
bakterial. Oleh sebab itu,diperlukan enzim dengan stabilitas termal pada
rentang suhu yang tinggi.
2. Stabilitas pH enzim
Semua reaksi enzim dipengaruhi oleh pH medium tempat reaksi terjadi
(Suhartono, 1989). Stabilitas enzim dipengaruhi oleh banyak faktor seperti
suhu, pH, pelarut, kofaktor, dan kehadiran surfaktan (Eijsink et al., 2005).
Dari faktor faktor tersebut, pH memegang peranan penting. Diperkirakan
perubahan keaktifan pH lingkungan disebabkan terjadinya perubahan ionisasi
enzim, substrat atau kompleks enzim substrat. Enzim menunjukkan aktivitas
maksimum pada kisaran pH optimum enzim dengan stabilitas yang tinggi
(Winarno, 1989).
Pada reaksi enzimatik, sebagian besar enzim akan kehilangan aktivitas
29
optimum untuk reaksi enzimatik. Inaktivasi ini terjadi karena unfolding
molekul protein sebagai hasil dari perubahan kesetimbangan elektrostatik dan
ikatan hidrogen (Kazan et al., 1997).
L. Kulit Pisang
Pisang kepok merupakan salah satu jenis buah-buahan tropis yang tumbuh subur
dan mempunyai wilayah penyebaran merata di seluruh wilayah Indonesia. Pisang
termasuk komoditas unggulan yang mudah diusahakan, berumur singkat, dan
dapat dipanen sepanjang tahun.
Menurut Tjitrosoepomo (1991) pisang kepok memiliki klasifikasi dan ciri
morfologi sebagai berikut :
Regnum : Plantae
Divisio : Spermatophyta
Sub Divisi : Angiospermae
Classis : Monocotyledonae
Ordo : Musales
Familia : Musaceae
Genus : Musa
Spesies : Musa paradisiaca L.
Bagian yang dapat dimakan dari buah pisang adalah dua per tiga bagian dan
sepertiga bagian sisanya merupakan limbah pisang. Angka tersebut merupakan
jumlah limbah yang cukup banyak yang dapat mencemari lingkungan apabila
30
pisang juga mengandung karbohidrat yang relatif tinggi yaitu 18,50 % (Setiawati
dkk., 2013). Limbah kulit pisang kepok memiliki kandungan selulosa yang tinggi
(60-65 %), hemiselulosa 6-8 %, dan lignin 5-10 % (Tjahyono, 1998). Limbah
kulit pisang kepok mengandung monosakarida terutama glukosa sebesar 8,16 %.
Limbah kulit pisang berpotensi untuk dimanfaatkan sebagai bahan baku dalam
pembuatan bioetanol melalui proses fermentasi (Setiawati dkk., 2013).
M. Hidrolisis Enzimatik
Hidrolisis merupakan proses pemecahan polisakarida di dalam biomassa
lignoselulosa, yaitu selulosa dan hemiselulosa menjadi monomer gula
penyusunnya. Pada hidrolisis sempurna selulosa akan menghasilkan glukosa,
sedangkan hemiselulosa menghasilkan beberapa monomer gula pentosa (C5) dan
heksosa (C6). Hidrolisis dapat dilakukan secara kimia (asam) atau enzimatik.
Aplikasi hidrolisis menggunakan enzim secara sederhana dilakukan dengan
mengganti tahap hidrolisis asam dengan tahap hidrolisis enzim selulase.
Keuntungan hidrolisis enzimatis dibandingkan dengan hidrolisis asam adalah
kondisi reaksi ringan dan tidak terjadi reaksi samping yang berarti (Seftian dkk.,
2012).
Faktor-faktor yang berpengaruh pada proses hidrolisis enzim diantaranya, yaitu
kualitas dan konsentrasi substrat, metode perlakuan awal yang diaplikasikan,
aktivitas enzim selulase dan kondisi proses hidrolisis seperti suhu dan pH. Suhu
31
hidrolisis. Suhu dan pH optimum pada enzim selulase umumnya pada 40 –50oC
dan pH 4–5, sehingga waktu yang digunakan tergantung pada kondisi tersebut.
Salah satu faktor utama yang berpengaruh terhadap hasil yang diperoleh dan
kecepatan hidrolisis enzimatis adalah substrat. Konsentrasi substrat yang tinggi
dapat menyebabkan penghambat yang memperlambat proses hidrolisis.
Terjadinya penghambat oleh substrat tergantung pada perbandingan antara
banyaknya enzim terhadap banyaknya substrat. Masalah pengadukan dan
perpindahan panas juga akan timbul pada substrat yang berkonsentrasi tinggi.
Banyaknya enzim yang ditambahkan pada substrat sangat berpengaruh terhadap
kecepatan proses hidrolisis. Semakin banyak enzim yang ditambahkan akan
semakin cepat proses hidrolisis yang terjadi dan hasil yang diperoleh juga
semakin banyak, tetapi semakin tinggi biaya yang harus dikeluarkan. Banyaknya
enzim yang ditambahkan pada substrat biasanya 5 –35 FPU/gram substrat.
Pengurangan biaya untuk penyediaan enzim pada proses hidrolisis enzim dapat
dilakukan dengan daur ulang enzim selulase. Bercampurnya enzim dalam
hidrolisat dan terbentuknya sisa proses yang berupa padatan (kemungkinan lignin)
mempersulit proses pemisahan enzim. Alternatif yang dapat dilakukan untuk
32
Gambar 8. Reaksi hidrolisis selulosa dengan enzim (Tomas et al., 2010).
N. Glukosa
Glukosa merupakan suatu aldoheksosa, disebut juga dekstrosa karena
memutar bidang polarisasi ke kanan. Glukosa merupakan komponen utama
gula darah, menyusun 0,065-0,11% darah kita. Glukosa dapat terbentuk dari
hidrolisis pati, glikogen, dan maltosa. Glukosa sangat penting bagi kita karena
sel tubuh kita menggunakannya langsung untuk menghasilkan energi.
Glukosa dapat dioksidasi oleh zat pengoksidasi lembut seperti pereaksi
Tollens sehingga sering disebut sebagai gula pereduksi (Budiman, 2009). hidrolisis enzimatik
selulase
hidrolisis enzimatik glukosa
hidrolisis enzimatik
Produksi degradasi glukosa
III. METODE PENELITIAN
A. Waktu dan Tempat Penelitian
Waktu penelitian dilakukan pada bulan Maret–Juli 2018 di Laboratorium
Biokimia Jurusan Kimia Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Lampung.
B. Alat dan Bahan
Alat-alat yang digunakan pada penelitian ini adalah alat-alat gelas, jarum ose,
mikropipet Eppendroff, neraca analitik, lemari pendingin, pembakar spirtus,
sentrifuga, magnetik stirer, autoclave model S-90N, oven, laminar air flow
CRUMA model 9005-FL, waterbatch shaker incubator HAAKE, pH meter,
penangas air, waterbath incubator, ayakan 100 mesh, dan spektrofotometer
UV-VIS Cary Win UV 32.
Adapun bahan-bahan yang digunakan adalah Potato Dextrose Agar (PDA),
(NH4)2SO4, KH2PO4, CaCl2, MgSO4, urea, FeSO4•7H2O, ZnSO4•7H2O, CoCl2,
pepton, NaOH, glukosa, Carboxymethyl Cellulase (CMC), akuades,
34
Na2SO3, HCl, Na2CO3, CuSO4•5H2O, Bovine Serum Albumin (BSA), akuades,
kantong selofan, dan zeolit. Adapun mikroorganisme yang digunakan adalah
jamur Aspergillus niger L-51 penghasil enzim selulase.
C. Prosedur Penelitian
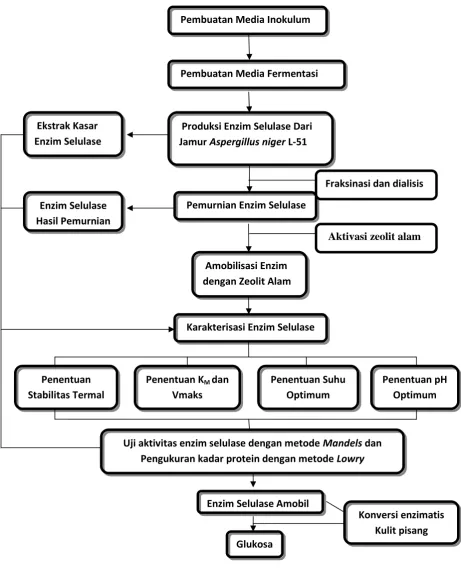
1. Pembuatan Media Inokulum dan Media Fermentasi
a. Pembuatan Media Inokulum
Media inokulum digunakan sebagai media adaptasi awal pertumbuhan dan
media perkembangbiakan spora jamur pada media cair. Media inokulum
dibuat dengan cara menimbang bahan-bahan yang terdiri dari (NH4)2SO40,14
g; K H2PO40,2 g; urea 0,03 g; CaCl20,03 g; MgSO40,03 g; FeSO4•7H2O
0,0005 g; ZnSO4•7H2O 0,00014 g; CoCl20,0002 g; pepton 0,075 yang
dilarutkan dalam buffer fosfat 0,2 M pH 5 sebanyak 100 mL dalam labu
Erlenmeyer 250 mL dan disterilisasi menggunakan autoclave pada suhu
121oC, tekanan 1 atm selama 15 menit. Selanjutnya media inokulum dikocok
dalam waterbath shaker incubator dengan kecepatan 130 rpm pada suhu
35oC selama 24 jam (Meriyanti, 2014).
b. Pembuatan Media Fermentasi
Media fermentasi yang digunakan (gL-1) terdiri dari (NH4)2SO41,4 g;
KH2PO42,0 g; urea 0,3 g; CaCl20,3 g; MgSO40,3 g; FeSO4•7H2O 0,005
g; ZnSO4•7H2O 0,0014 g; CoCl20,002 g; pepton 0,75 g; dan media
35
tekanan 1 atm selama 15 menit. Selanjutnya dimasukkan larutan media
inokulum sebanyak 2% total volume media fermentasi ke dalam
masing-masing media fermentasi secara aseptis lalu dikocok dalam waterbath shaker
incubator dengan kecepatan 130 rpm pada suhu 35oC selama 72 jam. Lalu
diuji aktivitas enzim selulase dengan metode Mandels (Meriyanti, 2014).
2. Isolasi Enzim Selulase
Isolasi enzim selulase dilakukan menggunakan metode sentrifugasi. Prinsip
sentrifugasi, yaitu memisahkan endapan dari suspensi dengan teknik sedimentasi.
Sentrifugasi digunakkan untuk memisahkan enzim ekstraseluler dari sisa-sisa gel.
Sentrifugasi dilakukan pada suhu rendah (di bawah suhu kamar) untuk menjaga
kehilangan aktivitas enzim (Suhartono, 1989). Untuk memisahkan enzim dari
komponen sel lainnya digunakan metode sentrifugasi pada kecepatan 5000 rpm
selama 20 menit. Filtrat yang diperoleh merupakan ekstrak kasar enzim yang
selanjutnya dilakukan uji aktivitas enzim selulase dengan metode Mandels.
3. Pemurnian Enzim Selulase
Setelah enzim selulase diisolasi, kemudian enzim tersebut dimurnikan
menggunakan metode fraksinasi dengan menggunakan ammonium sulfat
(NH4)2SO4dan dialisis.
a. Fraksinasi
Ekstrak kasar enzim yang telah diperoleh selanjutnya diendapkan dengan
36
yaitu 0-20%; 20-40%; 40-60%; 60-80%; dan 80-95%. Skema fraksinasi
dapat dilihat pada Gambar 9.
Ekstrak Kasar Enzim
+ (NH4)2SO4(0-20%)
Endapan (F1) Filtrat
+ (NH4)2SO4(20-40%)
Endapan (F2) Filtrat
+ (NH4)2SO4(40-60%)
Endapan (F3) Filtrat
+ (NH4)2SO4(60-80%)
Endapan (F4 Filtrat
+(NH4)2SO4(80-95%)
[image:58.595.117.458.158.490.2]Endapan (F5) Filtrat
Gambar 9. Skema proses fraksinasi enzim dengan ammonium sulfat
Endapan protein enzim yang didapatkan pada tiap fraksi kejenuhan
ammonium sulfat, dipisahkan dari filtratnya dengan sentrifugasi pada
kecepatan 5000 rpm selama 20 menit. Kemudian endapan yang diperoleh
37
b. Dialisis
Endapan enzim dari tiap fraksi hasil fraksinasi dimurnikan dengan cara
dialisis melalui membran semipermeabel (kantong selofan). Endapan
tersebut dimasukkan kedalam kantong selofan dan didialisis menggunakan
buffer fosfat pH 5 0,01 M selama 24 jam pada suhu dingin (Pohl, 1990).
Selama dialisis, dilakukan pergantian buffer selama 4-6 jam agar konsentrasi
ion-ion di dalam kantong dialisis dapat dikurangi.
Untuk mengetahui bahwa sudah tidak ada lagi ion-ion garam dalam kantong,
maka diuji dengan menambahkan larutan Ba(OH)2atau BaCl2. Bila masih
ada ion sulfat dalam kantong, maka akan terbentuk endapan putih BaSO4.
Semakin banyak endapan yang terbentuk, maka semakin banyak ion sulfat
yang ada dalam kantong. Selanjutnya dilakukan uji aktivitas dengan metode
Mandels dan diukur kadar proteinnya dengan metode Lowry.
4. Uji Aktivitas Enzim Selulase Metode Mandels(Mandels et al., 1976).
a. Pembuatan pereaksi untuk pengukuran aktivitas enzim selulase metode
Mandels (Mandels et al., 1976).
Pada labu takar 100 mL, dimasukkan 1 g DNS, selanjutnya ditambahkan 1 g
NaOH lalu dikocok hingga larut, kemudian ditambahkan 1 mL larutan
Na(K)-tartarat 40 %, 0,2 g fenol, dan 0,05 g Na2S2O3kemudian dilarutkan dengan
100 mL akuades hingga tanda batas.
b. Uji aktivitas enzim selulase metode Mandels (Mandels et al., 1976)
38
Sebanyak 0,25 mL enzim ditambah 0,25 mL larutan CMC 0,5% dalam
bufferfosfat pH 5,0 dan kontrol (0,25 mL enzim), yang masing-masing
diinkubasi selama 60 menit dalam waterbath incubator pada suhu 50oC.
Kemudian kontrol ditambahkan dengan 0,25 mL (larutan CMC 0,5% dalam
buffer fosfat pH 5,0) dan selanjutnya sampel dan kontrol ditambahkan 1 mL
pereaksi DNS dan dididihkan selama 10 menit pada penangas air. Kemudian
masing-masing ditambahkan 1,5 mL akuades lalu didinginkan. Setelah
dingin, serapannya diukur menggunakan spektrofotometer UV-VISpada λ
510 nm. Kadar glukosa yang terbentuk ditentukan dengan mengunakan kurva
standar glukosa.
5. Penentuan Kadar Protein Metode Lowry (Lowry et al., 1951).
a. Pembuatan pereaksi untuk penentuan kadar protein enzim selulase metode Lowry
1. Pereaksi A : 2 g Na2CO3dilarutkan dalam 100 mL NaOH 0,1 N
2. Pereaksi B : 5 mL larutan CuSO4.5H2O 1% ditambahkan kedalam 5 mL
larutan Na(K) tartarat 1 %
3. Pereaksi C : 2 mL pereksi B ditambahkan 100 mL pereaksi A
4. Pereaksi D : reagen folin ciocelteau diencerkan dengan akuades 1:1.
5. Larutan standar : larutan BSA (Bovine Serum Albumin) dengan kadar 0,
20, 40, 60, 80, 100, 120, dan 140 ppm.
b. Penentuan kadar protein
Sebanyak 0,1 mL enzim selulase ditambahkan 0,9 mL akuades lalu
39
selama 10 menit pada suhu ruang, setelah itu ditambahkan dengan cepat 0,5
mL pereaksi D dan diaduk dengan sempurna, didiamkan selama 30 menit
pada suhu kamar. Untuk kontrol, 0,1 mL enzim diganti dengan 0,1 mL
akuades, selanjutnya perlakuannya sama seperti sampel. Serapannya diukur
menggunakan spektrofotometer UV-Vis pada panjang gelombang 750 nm.
Untuk menentukan konsentrasi protein enzim digunakan kurva standar BSA.
6. Amobilisasi Enzim Selulase Hasil Pemurnian dengan Zeolit
a. Preparasi matriks zeolit
Serbuk zeolit diayak menggunakan ayakan berukuran 120 mesh. Aktivasi
zeolit alam dilakukan dengan cara mencampurkan 30 gram zeolit alam dan
100 mL HCl 3 M. Campuran dipanaskan sambil diaduk pada suhu 90oC
selama 2 jam, kemudian didinginkan, disaring dan dicuci dengan akuades
sampai zeolit tidak berwarna kekuningan lagi, lalu keringkan dalam oven
pada suhu 105oC selama 5 jam, dan disimpan dalam desikator (Septiani dan
Lisma, 2011).
b. Penetapan pH untuk proses pengikatan enzim selulase pada zeolit
Enzim selulase diikatkan dengan matriks dengan variasi pH 4; 4,5; 5; 5,5;
6; 6,5; dan 7 dengan menggunakan buffer fosfat 0,1 M. Kemudian matriks
diisi dengan 0,5 mL larutan enzim dan dielusi dengan buffer yang sesuai,
diaduk kemudian disentrifugasi selama 5-10 menit. Selanjutnya supernatan
40
c. Amobilisasi enzim selulase
Sebanyak 0,25 mL larutan enzim selulase diamobil dengan zeolit pada pH
optimum pengikatan. Enzim selulase diikatkan pada zeolit dengan cara
mencampurkan 0,25 mL enzim selulase dengan 0,25 gram zeolit. Kemudian
campuran diaduk hingga rata dan disimpan dalam freezer selama 30 menit.
Selanjutnya dicuci dengan akuades sebanyak tiga kali. Lalu dikeringkan pada
suhu kamar (Oktariani,2018).
d. Pemakaian berulang enzim amobil
Enzim amobil yang telah dipakai (direaksikan dengan substrat), dipakai
kembali untuk direaksikan kembali dengan substrat dengan uji metode
Mandels. Pemakaian berulang ini dilakukan hingga 4 kali.
7. Karakterisasi Enzim Selulase Murni dan Amobil
a. Penentuan suhu optimum
Untuk mengetahui suhu optimum kerja enzim dilakukan dengan
memvariasikan suhu yaitu 50; 55; 60; 65; 70; 75; dan 80oC. Selanjutnya
aktivitas enzim diukur dengan metode Mandels.
b. Penentuan KM dan Vmaks
Konstanta Michaelis-Menten (KM) dan laju reaksi maksimum (Vmaks) enzim
selulase ditentukan dengan memvariasikan konsentrasi substrat 0,1; 0,2; 0,4;