PROBLEMATIC AND NON-
PROBLEMATIC ECSTASY (MDMA)
USAGE: COGNITIVE AND
PSYCHOPATHOLOGICAL ASPECTS
KIRSTIE SOAR
A thesis submitted in partial fulfilment of the
requirements of the University of East London
for the degree of Doctor of Philosophy
Kirstie Soar
ACKNOWLEDGEMENTS
I would like to take this opportunity to thank Professor Andy Parrott and Dr John Turner for supervising my PhD thesis over the past years and for their useful comments and corrections. Thanks also go to Margherita Milani and Helen Fox for their valuable advice and support; providing a forum for discussion, and for commenting on some chapters; as well as their friendship and support during the earlier years of this research.
Special thanks go to my parents Graham and Anne Soar for their love and support when the times were bad and especially for keeping me financially afloat for the first years of this thesis. Additional thanks to Graham for proof-reading the final version of my thesis. Thanks also go to Professor Irvine Gersch for the part-time work, which helped the financial situation, but also for the valuable moral support he has provided throughout the years.
Thanks to all my colleagues in the School of Psychology at the University of East London for their encouraging support, especially the 'girls' (you know who you are) and Alan Martin from room GN141. Lastly, I would also like to thank everyone who volunteered to
participate in these research studies and those at the Big Issue magazine for facilitating contact with many participants through allowing me advertising space.
Kirstie Soar
ABSTRACT
This research thesis aimed to explore the apparent dichotomy of ecstasy (MDMA) users who report cognitive and psychopathological problems which they attribute to their use of this drug ("problematic" users), and those who report no adverse ecstasy-related effects ("non- problematic" users). In the first study, possible psychological sequalae linked to past ecstasy use were assessed in problematic and non-problematic ecstasy users using the modified Brief Symptom Inventory, aspects of the Rivermead Behavioural Memory Test, Tower of London and Auditory Verbal Learning Task. Problematic ecstasy users displayed higher
psychopathological symptoms and a small number of selective cognitive deficits compared to non-problematic ecstasy users and polydrug controls. However, problematic ecstasy use did not appear to be related to patterns of ecstasy use or polydrug use. Using the same
assessment measures, a case study based on a heavy problematic ecstasy user (RW), who had been abstinent for seven years, was presented. RW displayed cognitive deficits and extensive psychological problems suggesting that heavy ecstasy consumption may be associated with irreversible problems. The persistence of possible psychological and cognitive problems was further investigated in the second group study, using the same battery of tests. However no significant differences in cognitive and psychopathological performances were found between polydrug controls, current and ex-ecstasy users. It is argued that impairments in performance were possibly masked by poor cognitive performance in polydrug controls. The validity of the polydrug control group was addressed (in the third study) by assessing 20 drug-naive participants on the same measures. The introduction of a drug-naive control group only
suggested that problematic and non-problematic ecstasy users were exhibiting more errors on the Tower of London task compared to polydrug and drug-naive controls. The final study assessed psychopathological symptoms in problematic and non-problematic ecstasy users relative to drug-naive and polydrug controls, and explored factors which may be integral in the development of problematic ecstasy use, including certain pre-existing factors. Users were assessed on the BSI and Locus of Control scale. Pre-existing psychiatric histories, the intensity of ecstasy dosing and the role of polydrug use in relation to ecstasy use, appeared to contribute in higher psychopathological symptoms in problematic ecstasy users. Together these studies suggest that only self-reported problematic ecstasy users consistently display cognitive and psychopathological problems. For these vulnerable individuals the intensity of ecstasy use, patterns of other drug use and pre-existing psychiatric histories are thought to contribute to the development of these problems.
Kirstie Soar
CONTENTS
ACKNOWLEDGEMENTS ABSTRACT
ABBREVIATIONS
CHAPTER 1 - HISTORY, CULTURE & CHEMISTRY OF MDMA/ECSTASY History of MDMA/Ecstasy
Epidemiology
Patterns of ecstasy use Acute effects of MDMA
Psychopharmacology of MDMA MDMA Neurotoxicity in animals
Behavioural consequences of MDMA-induced neurotoxicity in animals Recovery from MDMA neurotoxicity in animals
Human neurotoxicity
1. CSF Assessment
2. Pharmacological Challenges 3. Neurological assessment
4. Electrophysiological Assessment 5. Dopaminergic functioning
Strength of evidence for neurotoxicity in ecstasy users Recovery from MDMA neurotoxicity in humans
CHAPTER 2 - POSSIBLE FUNCTIONAL CONSEQUENCES OF ECSTASY- INDUCED SEROTONERGIC NEUROTOXICITY
Long term psychopathological consequences associated with ecstasy use Long term cognitive consequences associated with ecstasy use
Cognition and altered serotonin functioning Recovery of cognitive abilities
Other important confounds/contributory factors Problematic ecstasy use
RATIONALE
CHAPTER 3 - RESEARCH STUDY 1
Cognitive and psychological profiles of non-problematic and problematic ecstasy users
CHAPTER 4 - CASE STUDY
Case study of persistent psychobiological problems attributed to ecstasy after seven years of abstinence
CHAPTER 5 - RESEARCH STUDY 2
Persistency of cognitive and psychological effects of recreational ecstasy use CHAPTER 6 - RESEARCH STUDY 3
Kirstie Soar
Addressing the methodological issue of a valid and reliable control group: comparisons with a drug-naive control group
CHAPTER 7 - RESEARCH STUDY 4
Differences in attributional styles between problematic and non-problematic ecstasy users: locus of control and drug attributions
CHAPTER 8 - OVERVIEW OF THESIS RESEARCH FINDINGS
- Overview of problematic and non-problematic ecstasy user profiles
Overview of psychopathology findings Overview of cognitive findings
Alternative interpretations and limitations Summary
CONCLUSIONS
REFERENCES APPENDICES
Kirstie Soar
FIGURES AND TABLES
Page CHAPTER 1
Table 1: Summary of pharmacological, imaging, physiological and CSF studies 23
investigating neurotoxicity in human ecstasy users
CHAPTER 2
Table 2: A summary of clinical case studies, where ecstasy appears to be a prominent feature 27
Table 3: Summary of empirical research assessing the long-term psychopathological effects 31
associated with ecstasy use
Table 4: Summary of empirical research assessing the long-term cognitive effects associated 42
with ecstasy use
CHAPTER 3
Table 5: Professional organisations where help/advice was sought by problem ecstasy users 74
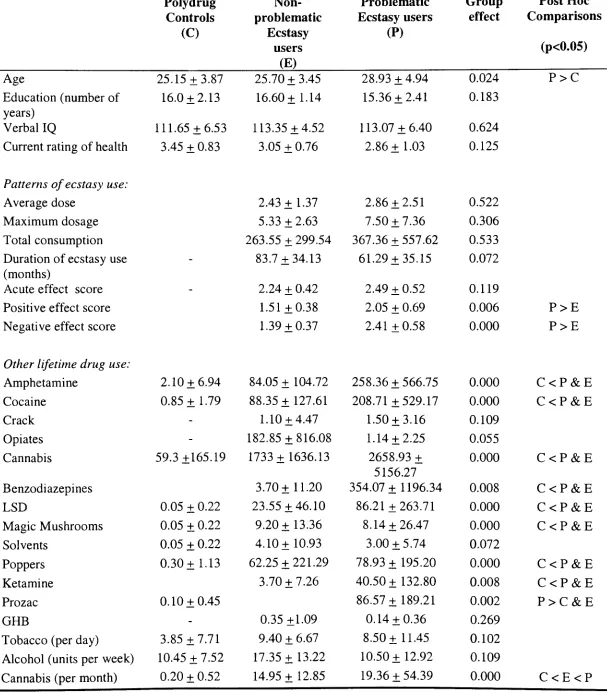
Table 6: Participant demographics, levels of illicit drug use and patterns of ecstasy use 76
consumption in polydrug controls, non-problematic and problematic ecstasy users
Table 7: Percentages of non-problematic and problematic ecstasy users reporting changes in 77
ecstasy use consumption and perceptions of their patterns of use
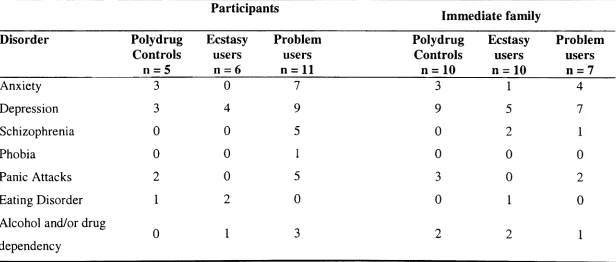
Table 8: Number of psychiatric disorders reported in polydrug, non-problematic and 77
problematic ecstasy users of those who reported individual psychiatric and family psychiatric histories
Table 9: Modified BSI subscale scores for polydrug controls, non-problematic and 78
problematic ecstasy users
Figure 1: BSI: mean somatisation scores 79
Figure 2: BSI: mean interpersonal sensitivity scores 79
Figure 3: BSI: mean depression scores 80
Figure 4: BSI: mean anxiety scores 80
Figure 5: BSI: mean phobic anxiety scores 81
Figure 6: BSI: mean paranoid ideation scores 81
Figure 7: BSI: mean psychoticism scores 82
Table 10: RBMT component and test scores, reaction times, TOL times and AVLT scores 85
by trial for polydrug controls, non-problematic and problematic ecstasy users
Figure 8: RBMT: mean scores for 'immediately remembering to deliver a message' 86
Figure 9: RBMT: mean recall scores of first and second name 86
CHAPTER 4
Table 11: Group mean scores for the BSI for polydrug controls, heavy ecstasy users and 105
individual case study scores.
Table 12: Group mean scores for the 9 primary symptom dimensions of the BSI for 106
normative data (psychiatric outpatients, psychiatric inpatients and non-patient normals) and individual case study scores.
Table 13: Group mean scores for the battery of cognitive tests, for polydrug controls, non- 107
problematic and problematic ecstasy users, normative data and individual case study scores.
Figure 10: Mean recall scores on the AVLT across all trials for RW and experimental groups 108
Figure 11: Mean planning times on TOL for RW and experimental groups 109
Figure 12: Mean solution times on TOL in seconds for RW and experimental groups 109
Figure 13: RBMT mean profile scores for RW and experimental groups 110
CHAPTER 5
Table 14: Participant demographics, levels of illicit drug use and patterns of ecstasy use 125
consumption in polydrug controls, current and ex-ecstasy users
Table 15: Percentage of group participants who answered 'yes' to questions concerning 126
patterns of ecstasy use: current and ex-ecstasy groups
Table 16: Reported drug use whilst using ecstasy, for current and ex-ecstasy using groups 127
Table 17: Modified BSI subscale scores for polydrug controls, current ecstasy and ex- 127
ecstasy users
Table 18: Cognitive Assessment data for polydrug controls, current and ex-ecstasy users 128
Figure 14: Mean AVLT recall scores, across all trials, for polydrug controls, current ecstasy 131
Kirstie Soar
users and ex-ecstasy users
Figure 15: Group mean planning times (in seconds) on the TOL 132
Figure 16: Group mean errors on the TOL I-* 2
CHAPTER 6
Table 19: Comparison A: Group demographics and drug use data from chapter 3 and current 148
drug-naive controls.
Table 20: Comparison A: Group BSI Scores from chapter 3 data and current drug-naive 149
controls.
Table 21: Comparison A: Cognitive assessment data for all groups from chapter 3 data and 150
current drug-naive controls.
Figure 17: Comparison A; Mean MDMA side-effects in all four groups 151
Figure 18: Comparison A: Mean cognitive failures in all four groups 151
Figure 19: Comparison A: Mean BSI somatisation scores for all groups 152
Figure 20: Comparison A: Mean BSI phobic anxiety scores for all groups 152
Figure 21: Comparison A: Mean BSI positive mood state scores 153
Figure 22: Comparison A: Mean errors made on the TOL 154
Figure 23: Comparison A: Mean delayed recall on the AVLT for all four groups 154
Table 22. Comparison B: Group demographics and drug use data from chapter 5 data and 156
current drug-naive controls.
Table 23. Comparison B: Group BSI Scores from chapter 5 data and current drug-naive 157
controls.
Table 24. Comparison B: Cognitive assessment data for all groups from chapter 5 data and 158
current drug-naive controls.
Figure 24: Comparison B: Mean BSI somatisation scores for all four groups 159
Figure 25: Comparison B: Mean BSI phobic anxiety scores for all four groups 159
Figure 26: Comparison B: Mean cognitive failures in all four groups 160
CHAPTER 7
Table 25: Group demographics, drug use data and locus of control scores for all groups 178
Table 26: Reported psychiatric and family psychiatric histories 179
Table 27: Professional organisations where help/advice sought by problematic ecstasy users 179
Figure 27: Mean scores on the LOG scale for all four groups 180
Figure 28: Mean BSI somatisation scores for all four groups 180
Table 28: Mean BSI scores across all dimensions and for all groups 182
Figure 29: Mean BSI negative psychobiology scores for all four groups 183
Figure 30: Mean BSI depression scores for all four groups 183
Figure 31: Mean BSI anxiety scores for all four groups 184
Figure 32: Mean BSI sexual functioning scores for all groups 184
Table 29: Number and percentage of drug-naive, polydrug controls, non-problematic and 187
problematic ecstasy users that reported that they had experienced positive and negative life changes
Table 30: Number and percentage of polydrug, non-problematic and problematic ecstasy 189
users who attributed life changes to more than one drug
CHAPTER 8
Table 35 Polydrug controls mean BSI dimension scores from the current studies compared 204
to normative data consisting of non-patients, psychiatric out-patients and psychiatric inpatients
Table 36 Non-problematic ecstasy users mean BSI dimension scores from the current studies 205
compared to normative data consisting of non-patients, psychiatric out-patients and psychiatric inpatients
Table 37 Drug naive controls' mean BSI dimension scores from the current studies 206
compared to normative data consisting of non-patients, psychiatric out-patients and psychiatric inpatients
Table 38 Problematic, non-problematic and ex-ecstasy users mean AVLT recall scores from 214
the current studies compared to normative data based on 20 to 29 year olds of average intelligence
Kirstie Soar
Table 39: Polydrug and drug naive mean AVLT recall scores from the current studies 215
compared to normative data based on 20 to 29 year olds of average intelligence
Diagram 1: A diagrammatic model summarising the potential factors which influence the 237
development of problematic ecstasy use
APPENDICES
A: Personal history and drug use questionnaire B: Ecstasy questionnaire
C: Ecstasy questionnaire: questions pertaining to the acute effects experienced on ecstasy D: Ecstasy questionnaire: questions pertaining to the long term effects of ecstasy
E: Drug use questionnaire: altered questions pertaining to cannabis use
F: Questionnaire pertaining to positive and negative life experiences and drug attributions. G: Tables 31-34: Percentages of reported positive and negative life changes in drug-naive, polydrug, non-problematic and problematic ecstasy users and which, if any drug(s), they attributed this change to.
H: Table 40 Locus of Control means scores (SD) for drug-naive, polydrug controls, non- problematic ecstasy, problematic ecstasy users and problematic ecstasy users who reported seeking help.
I: Figure 33 Mean LOG scores for drug-naive, polydrug controls, non-problematic ecstasy, problematic ecstasy users and problematic ecstasy users who reported seeking help.
J: Table 41: Mean BSI scores for all groups, including assessment of problematic ecstasy users and 'help-seeking' problematic ecstasy users.
K: Table 42: Ethnicity diversity in drug-naive, polydrug controls, non-problematic and problematic ecstasy users.
L: Modified BSI and subscale definitions
M: List of publications and presentations related to this thesis N: Peer reviewed journal publications from this thesis
O: Big Issue Advertisement P: Big Issue Advertisement
Q. Poster Advertisement - Chapter 3 R: UEL posters - Chapter 7
S: Application for ethical approval and confirmation of approval: Long term psychological effects of recreational ecstasy/MDMA
T: Application for ethical approval and confirmation of approval: Persistent neuropsychological effects of ecstasy/MDMA, in recreational users
U: Application for ethical approval and confirmation of approval: Personality characteristics of problem versus non-problem ecstasy (MDMA) users
V: Table 43: Chapter 3 - Modified BSI subscale ANCOVA statistics, with age as covariant : Table 44: Chapter 6 - Modified BSI subscale ANCOVA statistics, with age as covariant X: Information sheet & consent form
Y: Information sheet & consent form Z: Information sheet & consent form
Kirstie Soar ABBREVIATIONS: AVLT BDI BSI BZs CANTAB mCPP (r)CSF DNMS DNTMP DXM EEG EMCDDA EMQ ERBMT GC/MS GH GHB H-MRS IVE IMS LOG LSD MDA MDEA MDMA MHLC MI MEET 20 MR NART OCD PET PMQ RBMT SCL-90 SCL-90-R SERT SPECT SPSS SOSS STAI STAXI TOL VMAT WAIS-R WMS-R 5-HIAA 5-HT
Auditory Verbal Learning Task Beck Depression Inventory
Brief Symptom Inventory
Benzodiazepines
Cambridge Neuropsychological Test Automated Battery
1 (3-Chlorophenyl)piperazine dihydrichloride Cerebrospinal fluid
Delayed non-matching to samples test Delayed non-match to place procedure pheneylethylamine dextromethorphan Electroencephalogram
European Monitoring Centre for Drugs and Drug Addiction Everyday Memory Questionnaire
Extended Rivermead Behavioural Memory Test Gas chromatography/mass spectrometry
Growth Hormone
Gamma-h y drox ybutyrate
Proton Magnetic Resonance Spectroscopy
Impulsivity, Venturesomeness and Empathy Questionnaire ion mobility spectrometry
Locus of Control Scale d-lysergic acid
3,4-methlyenedioxyamphetamine
3,4-methylenedioxy-N-ethylamphetamine 3,4-methlenedioxymethamphetamine Multi-dimensional Locus of Control Myo-inositol
Matching Familiar Figures Test 20 Magnetic Resonance Imaging National Adult Reading Test Obsessive-compulsive disorder Positron Emission Tomography Prospective Memory Questionnaire Rivermead Behavioural Memory Test Symptom Checklist
Revised Symptom Checklist Serotonin transporter
Single Photon Emission Computed Tomography Statistical Package for the Social Sciences
Self-ordered spatial search task State-trait anxiety inventory
State-trait anger expression inventory
Tower of London
Vesicular monoamine transporters
Wechsler Adult Intelligence Scale, Revised. Wechsler Memory Scale, Revised
5-hydroxyindoleacetic acid
5-hydroxytryptamine (Serotonin)
Kirstie Soar Chapter 1
CHAPTER 1
Kirstie Soar Chapter 1
HISTORY OF MDMA/ECSTASY
3,4-methlyenedioxymethamphetamine (MDMA) was first synthesised and patented by Merck in 1914. Many believe it was patented as an appetite suppressant, but according to Holland (2001) there was no use mentioned for MDMA within this patent application and such claims probably arose from MDMA's chemical cousin MDA (3,4-methlyenedioxyamphetamine), which was patented by SmithKline French in 1958 and tested as an appetite suppressant in humans.
MDMA was never marketed due to the lack of commercial interest and therefore did not become available on the public market. It resurfaced briefly during the 1950's, in a toxicological and behavioural report from a study conducted by the US Army Chemical
Centre when they secretly tested a number of psychoactive chemicals for military application (Shulgin & Nichols, 1978). It was not until 1976, that Alexander Shulgin synthesised
MDMA and suggested its potential use in psychotherapy (Holland, 2001). From Shulgin's experiences, it was suggested that MDMA's subtle mood modifying characteristics could be used as a adjunct to psychotherapy, with the strengthening of the therapeutic alliance by enhancing trust, freeing patients from defensive anxiety and making them more emotionally open. Its success in fostering introspection and verbalisation during therapy led to a slow spread of its use in underground psychotherapeutic work in the late 70s and early 80s (Greer and Tolbert, 1986). The drug's ability to alter consciousness and induce such subtle mood changes leaked out of the therapeutic community, and as a result MDMA was being used amongst students throughout the US under its new name ecstasy.
In 1977 MDMA and other similar psychedelic amphetamines such as MDA and MDEA (3,4- methlyenedioxyethylamphetamine) were listed as class A drugs under the Misuse of Drugs Act 1971, in the United Kingdom (UK). In the United States (US) the drug was still legal until the mid 1980s. However due to numerous reports of misuse in conjunction with a widely publicised report of brain damage in rats caused by a similar drug, MDA, the US Department of Justice's Drug Enforcement Agency (DEA) followed the British Government and placed MDMA and related derivatives on the list of substances under international
Kirstie Soar Chapter 1
Despite ecstasy's illegality within the UK, towards the end of the 1980s, it started to become a popular drug of choice at all-night dance parties, commonly known as 'raves', due to its
modulation of emotional state; inducing feelings of relaxation, fearlessness and happiness which, along with its stimulant properties, enhanced the ability and enjoyment of dancing. Since then ecstasy has grown in popularity as a recreational drug and according to Saunders (1997), has been used by about one to five million people within Britain.
EPIDEMIOLOGY
Worldwide estimates of ecstasy use within the general population are hard to establish, but according to Holland (2001) nearly one million people take ecstasy every weekend. Ecstasy along with amphetamine is the second most commonly used drug in Europe (EMCDDA, 2001). Since the re-emergence of ecstasy in the late 80s, its use increased in the 1990s but now appears to be stabilising (EMCDDA, 2001). In 1998 prevalence rates in the general population of EU (European Union) adults were 0.5-3% (EMCDDA, 1998) and by 2002 this had only risen to 5% (EMCDDA, 2002).
The UK accounts for most of the ecstasy use in the EU, with rates at 11% (EMCDDA, 2002). Outside the UK the highest European rates appear in Ireland 8.9% (EMCDDA, 2001) and Latvia 6% (ESPAD, 2001). Similar rates to the EU have been reported elsewhere in the world. In Australia, for example, the National Drug Strategy Household Survey reports rates at 3%, (Commonwealth Department of Health and Family Services, 1996).
In the US, ecstasy has also been reported as the second most frequently tried illicit drug after cannabis (Pope et al, 2001). However, despite the prevalence rates stabilising in the EU, reports from America indicate a dramatic increase in use in the later part of the 90s amongst
16-26 year olds (Johnston et al, 2002). Amongst American college students, rates increased significantly by 69% between 1997 and 1999, from 2.8% to 4.7% (Strote et al, 2002). Whilst one study showed an increase in prevalence rates from 4.1% in 1989 to 10.1% in 1999 (Pope et al, 2001), however these rates were based on usage within just the one college.
Prevalence rates are higher in younger more specific age groups, with 6% of 15-34 years olds having used the drug. In a school survey 5% of 15-16 year olds reported its usage
Kirstie Soar Chapter 1
16/17 years (Measham et al, 1998), which is thought to reflect the shift in leisure activities of the youth as they start reaching the age to attend licensed clubs, bars and pubs. This is
reflected in the greater use amongst university students at 13% (Webb et al, 1996).
Additionally, prevalence rates are again much higher amongst youngsters that attend the rave or 'party' scene, in which ecstasy is a popular drug of choice; this is fairly consistent across the world. Amongst rave attendees in the UK, reported use varied from 82% to 96% (Riley et al, 2001; Forsyth, 1996; Winstock et al, 2001). In Australia, reported use amongst people in the rave scene has been seen to vary between studies, with Topp et al (1999) reporting 76% compared to an earlier report of 89% (Lenton et al, 1997); in the US, Arria et al (2002) reported lifetime use by 89% of rave attendees; and in Canada rates were reported as 65.2% (Gross et al, 2002).
PATTERNS OF ECSTASY USE
Recreational ecstasy use has been commonly associated with the rave or dance scene, were its use has been seen as a dance drug, and has been used in this setting more than any other
recreational drug (Forsyth, 1996). However, its usage in more recent times has been seen to be shifting away from large dance events to more geographically diffuse club, bar and private settings (EMCDDA, 2000). In particular, a sub-cultural music preference for house/techno music has been shown to be the greatest predictor of ecstasy use (Pederson & Skrondal,
1999).
The assumption that drug users are unemployed, uneducated and come from socially deprived backgrounds does not fit the profile of a common recreational ecstasy user. Ecstasy users are not academic underachievers (Strote et al, 2002), but are generally employed or in higher education (Riley et al, 2001). In one survey of recreational ecstasy users, 80% reported having been in further education (Forsyth, 1996), and another survey reported 65% of users were currently employed (Winstock et al, 2001).
Kirstie Soar Chapter 1
The drug is usually consumed orally in multiple tablets (Winstock et al, 2001), but there is a subsample of users (16%), mostly found in Australia, that report injection of ecstasy (Topp et al, 1999). Boys et al (2001) examined the reasons why a sample of poly-substance users chose to use ecstasy, and they discovered that 91% of users reported that the main reason for using ecstasy was to 'keep going'. Also, 78% reported using for its euphoric/elation effects, 80% for its ability to enhance activity, 72% to stay awake and 68% to feel intoxicated.
Recreational ecstasy users rarely report sole use of the drug. In fact, Hammersley (1999) failed to find a single interviewee that reported sole use of ecstasy. Instead ecstasy users are more likely to be polydrug users, in that they often consume ecstasy in combination with other substances, such as tobacco, cannabis, speed, alcohol, d-lysergic acid (LSD), and amyl nitrate (Winstock et al, 2001; Topp et al, 1999; Atria et al, 2002). Ecstasy 'polydrug' users have also been shown to consume cannabis, alcohol, tobacco, amphetamine, heroin and benzodiazepines (BZs) whilst coming off the drug (Topp et al, 1999; Winstock et al, 2001). There are a number of potential reasons why people choose to consume ecstasy in the context of polydrug use. Firstly, taking several psychoactive compounds together may enhance the effects of ecstasy (Schifano, 2004). In particular, the use of alcohol prior to taking ecstasy is thought to enhance the 'high' from MDMA (Schifano, 2004), whilst stimulants, such as amphetamine and cocaine are thought to maintain the arousal and alertness of the ecstasy experience. Sedatives and relaxants, such as opiates, BZs and cannabis, are reported to
relieve the unpleasant sub-acute effects of ecstasy (Scholey et al, 2004). Secondly, it has been repeatedly reported that there is a decrease in the desired effects of ecstasy following repeated use (Scholey et al, 2004; Parrott, 2005). This chronic tolerance to ecstasy may lead to the use of other stimulants and hallucinogens in order to try and achieve the initial ecstasy effects (Schifano, 2004). Some ecstasy users have indeed reported that this is the reason behind the need to consume other psychoactive drugs in the context of ecstasy use (Scholey et al, 2004).
ACUTE EFFECTS OF MDMA
Kirstie Soar Chapter 1
effects in humans. The behavioural and psychological effects include increased positive mood, feelings of euphoria, increased physical and emotional energy, heightened sensual awareness, depersonalisation, derealisation, fear of loss of control, increased extroversion, mild perceptual alterations, increased self-confidence, accelerated thinking, thought blocking, difficulty in concentrating and impaired decision making (Downing, 1986; Liechti et al,
2000a; Liechti et al 2000b; Gamma et al, 2000; Cami et al, 2000; Liechti & Vollenweider, 2001). Reported physiological effects of MDMA include suppressed appetite, pupil dilation, jaw clenching (trismus), enhanced tendon reflexes, increased heart rate and blood pressure,
increased peripheral body temperature, dry mouth, impaired balance and dizziness (Downing, 1986; Grob et al, 1996; Liechti et al, 2000a; Gamma et al, 2000; Liechti & Vollenweider, 2001).
The subjective psychological and physiological effects from pharmaceutical grade MDMA are similar to the subjective acute effects of ecstasy. It has been repeatedly reported that ecstasy users experience elation, agreeableness, euphoria, increased energy, confidence,
exhilaration, warmth and friendliness, calmness and relaxation, increased perception of sound, colour and touch, confusion, increased heart rate, increased body temperature, sweating and dehydration, trismus, sexual arousal, papillary dilation, bruxism, lower back pain and nausea (Cohen, 1995; Davison & Parrott, 1997; Parrott & Stuart, 1997; Verheyden et al, 2003).
Following the acute effects of ecstasy, users report subacute effects in the following 24 to 48 hour period, which tend to be negative effects (Verheyden et al, 2003), characterised by symptoms such as muscle aches, lethargy, fatigue, moodiness, depression, anxiety,
Kirstie Soar Chapter 1
brooding and bad dreams. The fact that the subjective effects of acute and subacute ecstasy use are similar to the acute and subacute effects of MDMA administration in placebo
controlled studies, suggests that in most cases ecstasy contains the active compound MDMA more so than any other psychoactive drug.
PSYCHOPHARMACOLOGY OF MDMA
3,4-methylenedioxymethamphetamine (MDMA) is a completely synthetic substance. It is a ringed substituted amphetamine derivative, which resembles the structure of the
hallucinogenic material mescaline. As a result its pharmacological effects are a blend of those from amphetamine and mescaline, affecting 5-hydroxytryptamine (Serotonin; 5-HT) and
dopamine-containing neurons, and also other neurotransmitter systems; hence the frequent references to MDMA being a rather 'messy' drug.
MDMA acts as an indirect monoaminergic agonist, stimulating the release of 5-HT from the presynaptic neuron (Nixdorf et al, 2001) and inhibiting the reuptake of 5-HT (Iravani et al, 2000; Mechan et al, 2002), causing the synapse to be flooded with atypically large amounts of 5-HT. At the same time it also causes the release of dopamine; although this effect is weaker than the MDMA-induced efflux of 5-HT (Yamamoto and Spanos, 1988; Gough et al, 1991; Nixdorf et al, 2001) and, possibly, norepinephrine (Rothman et al, 2001). Additionally, MDMA produces a rapid inactivation of trytophan hydroxylase (Stone et al, 1989; Schmidt and Taylor, 1988), an enzyme necessary for the synthesis of 5-HT, thus slowing 5-HT
replenishment. For a more comprehensive review of the pharmacology of MDMA see Green et al (2003).
The acute boost in monoamine activity generates the unique reinforcing effects of the drug. Selective blocking studies show that the subjective effects of MDMA (feelings of elation, euphoria and well-being, emotional closeness and sensory pleasure; Liechti et al, (2000a & 2000b) and Cami et al, (2000)), are largely dependent on carrier-mediated 5-HT release, whilst the stimulant-like mood effects appear to be related, at least in part, to dopamine D2 receptor stimulation (see Liechti & Vollenweider, 2001). The mild hallucinogenic like perceptual effects appear to be due to serotonergic 5-HT2-receptor stimulation (Liechti & Vollenweider, 2001). Because MDMA's dopaminergic actions are similar to those of
amphetamine, whilst its serotonergic effects are closer to LSD, in behavioural terms MDMA
Kirstie Soar Chapter 1
displays similarities to both amphetamine and LSD. However, its strong euphoric properties appear to be unique to MDMA itself.
MDMA NEUROTQXICITY IN ANIMALS
There is currently a large body of preclinical research, which shows that MDMA has neurotoxic effects on brain serotonin neurons in animals (for a comprehensive review see Ricaurte et al, 2000 and Green et al, 1995 & 2003). Evidence has arisen from studies using a variety of experimental techniques and neurochemical, neuroanatomcial and functional
measures of 5-HT neurons, which have consistently demonstrated MDMA-induced
serotonergic injury. These include long-term decreases in levels of 5-HT, the metabolite 5- HIAA, 5-HT transporters, tryptophan hydroxylase activity, anterograde transporters and vesicular monoamine transporters (VMAT) and histological evidence of 5-HT axon
degeneration (Ricaurte, 2000). The areas of the rat brain that appear to be the most sensitive to these alterations in serotonergic activity are the striatum, hippocampus and prefrontal cortex, with smaller but significant effects in the brain stem and hypothalamus (Sabol et al,
1996).
This selective serotonergic neurotoxicity induced by MDMA has been demonstrated in a variety of animals, such as rats (Finnegan et al, 1988; Lew et al, 1996), guinea pigs (Battaglia et al, 1988), baboons (Scheffel et al, 1998) and monkeys (Ricaurte et al, 1988). Non-human primates have been shown to be considerably more sensitive to the serotonin depleting effects than rats (Ricaurte, 1989). The only animal to date that is resistant to this MDMA-induced serotonergic neurotoxicity is the mouse. In the mouse dopamine neurons are affected whilst serotonin neurons appear to be spared (Battaglia et al, 1988; O'Shea et al, 2001).
Studies have also suggested that regardless of the route of administration and dosage of MDMA, serotonin neurotoxicity is still evident. Orally administered MDMA has been seen to produce toxic effects that are comparable to those induced by subcutaneous administration in rats and monkeys (Finnegan et al, 1988; Ricaurte et al 1988). Serotonergic neurotoxicity also appears to be dose-dependent (Battaglia et al, 1988; Finnegan et al, 1988). However, even though multiple doses of MDMA are more effective than single doses at depleting serotonin, Ricaurte et al (1988) showed that even a single dose of MDMA could produce long-lasting depletions in monkey brain 5-HT.
Kirstie Soar Chapter 1
These effects in animals produced by oral administration and single and multiple dosing of MDMA, often at concentrations analogous to MDMA amounts in ecstasy users (Ricaurte, 2000), has raised legitimate concerns about possible MDMA-induced neurotoxicity in
humans (Parrott, 2000; Turner and Parrott, 2000). Furthermore, comparative animal data has shown that the level of neurotoxicity also increases in high ambient temperatures (Broening et al, 1995; Colado et al, 1998; Malberg and Seiden, 1980). However, caution needs to be taken in interpreting many of the animal findings and extrapolating to human MDMA use. The dosing regimens used in animals vary greatly between studies, and many studies look at
amounts that actually cause neurotoxic effects, rather than looking at doses that are equivalent to those used by human recreational ecstasy users. Many animals employed in these studies also have been used in previous studies assessing the acute effects of several other
psychoactive compounds (Frederick et al, 1995; Frederick et al, 1998; Taffe et al, 2001).
Behavioural consequences of MDMA-induced neurotoxicity in animals
Considering the evidence of MDMA-induced neurotoxicity in animals, it is important to determine whether such neurotoxicity has behavioural consequences, and if so, what areas of behaviour are affected. Compared to the number of animal studies demonstrating
neurotoxicity, there is a somewhat limited behavioural data in animals. Studies investigating various regimens of MDMA on animal behaviour have consistently failed to find any baseline changes in performance across a number of behavioural tests, despite marked reductions in 5- HT and 5-HIAA (up to 80% decrease, in the study of Winsaur et al, 2002) in areas including the hippocampus, striatum, neocortex, caudate, and thalamus (Ricaurte et al, 1993; Seiden et al, 1993; Marston et al, 1999; Dornan et al 1991; Frederick et al, 1995; Winsaur et al, 1993), which have, in some studies, lasted up to 4-7 months (Ricaurte et al, 1993; Taffe et al, 2001; Frederick et al 1998).
In a majority of the studies that have demonstrated cognitive and behavioural dysfunction relative to control animals, this has not persisted longer than 7 days post-MDMA treatment (Slikker et al, 1989; McNamara et al, 1995; Robinson et al, 1993; Taffe et al, 2001;
Maldonado & Navarro, 2001; Navarro et al, 2004). McNamara et al (1995) demonstrated behavioural changes in locomotor activity in rats, during the 4 days of MDMA
Kirstie Soar Chapter 1
controls, despite MDMA-induced depletions of 5-HT and 5-HIAA in the frontal cortex and amygdala, 7 days after treatment. Similarly, rats who showed 73% depletion in serotonin concentrations in the neocortex and 32% in the caudate nucleus relative to controls, only demonstrated a mild impairment in developing an efficient search strategy, on a spatial- navigation learning set-task, on the first three days of training, after MDMA-treatment.
However, once learnt, memory performance concerning this location was equivalent to that of controls (Robinson et al, 1993). Taffe et al (2001) reported behavioural impairments in
rhesus monkeys treated with MDMA. Performance in memory, on the delayed non-matching to samples test (DNMS) and the self-ordered spatial search task (SOSS); reinforcer efficacy and sustained attention on a progressive-ratio (PR) schedule of responding task; fine motor control on a bimanual motor task reaction; and reaction time were all impaired relative to controls. Task performance returned to pre-treatment baseline levels within one week after MDMA treatment, despite reports of a 44% reduction in 5-HIAA concentrations which
persisted for approximately 3 months after MDMA treatment. However, they did report one single animal's behavioural performance to be severely affected which persisted for up to two months.
These earlier studies suggest that any cognitive and behavioural dysfunction in MDMA treated animals is the result of the acute and sub-acute effects of MDMA, rather than
functional consequences of neurotoxicity. This evidence also suggests that the neurotoxic effects of the drug may not be manifested behaviourally, despite the underlying
neurochemical changes. However, more recent studies have shown behavioural effects related to MDMA-neurotoxicity. Martson et al (1999) reported a selective deficit in performance in rats on the delayed non-match to place procedure (DNTMP), 16 days following MDMA exposure. MDMA-treated rats did not show the improvement in
performance at the longer delays, as seen in the control rats. In addition, MDMA treated rats showed reductions of 5-HT function upon post-mortem analysis. Memory impairments, on object recognition tasks, in MDMA treated rats have also been shown, one week (Piper & Meyer, 2004) and 10-12 weeks after drug treatment (McGregor et al, 2003). Taffe et al (2002) have showed lasting behavioural sensitivity in monkeys. When pharmacologically challenged with l(3-Chlorophenyl)piperazine dihydrichloride (mCPP), vigilance and reaction time (5-choice reaction time), and reinforcer efficacy and sustained attention (PR), where disrupted in monkeys treated 13-months previously with MDMA . Taffe et al (2002) also
Kirstie Soar Chapter 1
showed 50% reductions in 5-HIAA, 2-17 weeks post MDMA treatment. However, cognitive dysfunction was quite small compared to the magnitude of serotonin depletion.
Studies also indicate that social interaction and anxiety-related behaviours are disrupted in MDMA-treated animals, which are lasting after the cessation of MDMA administration.
McGregor et al (2003) reported that rats pre-treated with MDMA displayed increased anxiety in a social interaction test and emergence test; demonstrating a shorter duration of social
interaction and fewer social interaction bouts, and took longer to emerge in the open field compared to control groups 8-10, weeks following drug administration. In addition, rats were reported to have shown depressive symptoms in the forced swim test, displaying reduced escape attempts and increased immobility. However, these symptoms were only evident on the third day of testing. Ho et al, (2004) did not provide indications of anxiety, reduced social interaction and depressive symptoms, using similar testing paradigms (open field, plus maze and forced swim test). However, Ho et al (2004) only injected animals with a single dose (7.5mg/kg) of MDMA compared with a dosing regimen of 5 mg/kg every 4 hours on 2 consecutive days (20 mg/kg per day) in McGregor et al's study. This, together with the animal studies which have demonstrated small, if any, changes in behaviour compared to the relatively large amounts of serotonin depletion, suggests that it may be the magnitude of neurotoxicity produced by MDMA that is crucial in behavioural studies. The research data suggests that there may be a threshold effect of 5-HT, below which no behavioural
consequences will be observed. In many of the studies which have not demonstrated behavioural disturbances or very limited and selective deficits, there may not have been sufficient neurotoxic damage over and above that 5-HT threshold, in order to interfere with behavioural and cognitive functioning.
The shortage and very selective nature of long-term behavioural dysfunction in these studies may be because of the specific tests employed and the differing paradigms: they may not be sensitive enough to detect changes in the 5-HT system; and the behaviour in question may not be influenced directly by the 5-HT system. What is certain is that the animal behaviour
research has utilised various behavioural measures, covering numerous behavioural domains which are known to be sensitive to small changes in monoamine neurotransmitter
concentrations (Seiden et al, 1993). One possibility to account for discrepancies in some of these tests is that the lasting effects of MDMA may depend on subject-dependent factors.
Kirstie Soar Chapter 1
Taffe et al (2001) drew attention to one monkey of the six that they assessed, which demonstrated marked deficits in performance which lasted for up to 2 months. Ho et al (2004) also demonstrated behavioural impairments 3 weeks after MDMA-treatment only in animals with low anxiety levels, but not high anxiety levels. This demonstrates certain individual vulnerabilities to MDMA-induced effects.
Recovery from MDMA neurotoxicitv in animals
Animal studies have shown that these neurodegenerative effects in the serotonin system are long lasting (up to one year). However, there is evidence which shows 'recovery' of this serotonergic function; although subsequent reorganisation and/or function may be abnormal. Recovery of serotonin reuptake sites has been shown in rats, with the concentration of 5-HT reuptake sites returning to control levels after 12 months (Battaligia et al, 1991; Sabol et al,
1996; Scheffel et al, 1998; Scanzello et al, 1993). Taffe et al (2001) demonstrated a 44% reduction in 5-HIAA concentrations and altered peak latencies in brainstem auditory evoked potentials in rhesus monkeys, which persisted for approximately 3 months post-MDMA treatment, yet in the fourth month both these measures normalised.
Ricaurte at al (1992) showed evidence of partial recovery in some brain regions of nonhuman primates (hippocampus, caudate nucleus, frontal cortex). However, after 18 months it was evident that recovery did not continue in all regions with the exception of the thalamus and hypothalamus. Thus the rate and degree of recovery appears to depend on the brain region (Lew et al, 1996; Battaglia et al, 1991; Sabol et al, 1996), with some brain regions showing an increase in 5-HT functioning, mainly the hypothalamus, but others showing persistent
decreases (Ricaurte et al, 1992; Scheffel et al, 1998); also, recovery was not always normal. Fischer et al (1995) demonstrated that the pattern of some of the serotonin axonal sprouting in both rats and monkeys was abnormal, especially in the amygdala and hypothalamus where neuron axons were reinnervated or hyperinnervated, suggesting that MDMA actually lead to a reorganisation of the serotonin system. Such abnormal patterns were also evident in monkeys seven years after MDMA treatment, although some regions were less severely affected than those observed at 18 months (Hatzidimitriou et al 1999). This abnormal reorganisation of 5- HT axons and axon terminals is synonymous to the 'pruning effect' seen with a number of neurotoxins (Ricaurte et al, 2000), where nerve cells will often grow replacement terminals,
Kirstie Soar Chapter 1
where there has been damage, resulting in a different dendritic pattern. This pruning effect provides further evidence of MDMA's neurotoxicity (Ricaurte, 2000), but also of potential attempts of the serotonergic system to recover from such damage.
Serotonergic recovery also depends on the animal treated. On inspection of experimental animals, Scanzello et al (1993) revealed a group of rats that did not show signs of recovery, whilst others did. Those that did not recover had severe and enduring serotonergic deficits in multiple brain regions. Thus, it appears that the recovery of serotonin neurons is region dependent and also varies between and within species. Monkeys tend to be more sensitive and damage appears to be permanent, whereas with rats there is some indication that recovery takes place, but the question is whether this is sustained? Such differences in recovery may be due to the severity of damage sustained from MDMA; the more severe the damage, or more highly arborized, the lower the probability of recovery (Fischer et al, 1995). Disparity may also be due to genetic differences, individual vulnerabilities and other parameters like age, health status, diet and fluid supply etc.
Whilst studies have demonstrated the potential for animals to show a certain level of recovery from neurotoxic effects of MDMA, this recovery is not always normal and there is a paucity of research indicating the behavioural consequences this has. As tests for neurotoxic effects become more refined, then the assessment of behavioural recovery should become possible.
HUMAN NEUROTOXITICY
That fact that nearly all animal species tested are sensitive to the neurotoxic potential of MDMA (with the exception of the mouse), suggests that humans too will be sensitive to the toxic effects. Applying the well-established principles of interspecies scaling also strengthens the case for human sensitivity to MDMA-induced serotonergic neurotoxicity. The principle of interspecies scaling is that smaller animals require higher doses of a psychoactive drug to achieve the equivalent effect. As demonstrated in the animal literature, rodents require higher doses of MDMA to produce the same neurotoxic effect as in non-human primates. Using this technique, the dosages of MDMA known to be neurotoxic in animals falls squarely in the range of dosages typically used by recreational ecstasy users: between 75-125mg of MDMA (Ricaurte et al, 2000). The fact that most recreational ecstasy users consume more than one
Kirstie Soar Chapter 1
dose of ecstasy on one occasion (Winstock et al, 2001), and that the acute effects of ecstasy mimic these controlled effects of pharmaceutical MDMA (Leicht & Vollenweider, 2001), strongly suggests that recreational ecstasy users are ingesting neurotoxic doses of MDMA.
A number of lines of evidence to support the notion that MDMA-induced serotonergic
neurotoxicity occurs amongst recreational ecstasy users has emerged in the last 10 years (see table 1). Methods of assessing serotonin neurotoxic changes in the living human brain
include analysis of cerebrospinal fluid (CSF) 5-hydroxyindoleacetic acid (5-HIAA - a major metabolite of serotonin) concentrations and pharmacological challenges using 5-HT agonists. Less invasive methods of assessing changes have included Positron Emission Tomography (PET), Single Photon Emission Computed Tomography (SPECT), Proton Magnetic
Resonance Spectroscopy (H-MRSI), Electroencephalograms (EEG) and assessment of auditory evoked potentials.
1. CSF Assessment
The first study which measured the concentration of 5-HIAA in cerebrospinal fluid (CSF) did not find any significant indication of neuronal alteration in ecstasy users compared to age- matched controls (Peroutka et al, 1987). However, subsequent studies using the same technique and larger participant numbers have consistently demonstrated significantly reduced concentration levels of CSF 5-HIAA in human ecstasy users compared to controls (Ricaurte et al, 1990; McCann et al, 1994; Bolla et al, 1998; McCann et al, 1999). Bolla et al, (1998) demonstrated a dose-response effect, with the concentration of 5-HIAA decreasing as the dose of reported ecstasy use increased. However, other studies which have looked at this correlation, have not found a significant relationship between the concentration levels of 5- HIAA and number of ecstasy exposures (Ricaurte at al, 1990; McCann et al, 1994).
Measuring levels of CSF does not indicate which areas of the brain are potentially affected by ecstasy, if at all; because CSF can also be a consequence of psychological changes; (e.g CSF 5-HIAA is lower in depressed individuals (Becker et al, 1995)).
2. Pharmacological Challenges
Pharmacological challenges using 5-HT modulators, such as L-tryptophan, d-fenfluramine and M-chlorophenylpiperazine, have also been used to assess possible MDMA-induced
Kirstie Soar Chapter 1
neuronal alterations but with less consistency in their findings. Price et al (1989) first
demonstrated altered 5-HT functioning compared to healthy controls, using the precursor L- tryptophan. Ecstasy users showed blunted responses to the effects of L-tryptophan, as
measured by prolactin concentration, yet the difference compared to healthy controls failed to reach significance. A subsequent study using L-tryptophan also did not find any significant differences in prolactin concentration between ecstasy users and controls (McCann et al,
1999). A more recent study by Curran & Verheyden (2003) showed that only ex-ecstasy
users (who had used more than 20 tablets, but not within the last year), and not current ecstasy users (who had used more than 20 tables within the last year), showed significantly higher levels of total and free plasma tryptophan following tryptophan manipulation compared to polydrug controls. However, such differences in these ex-users may reflect pre-morbid differences in their 5-HT function.
The studies by Gerra et al (1998 & 2000) and Verkes et al (2001) have indicated alterations in serotonin functioning in ecstasy users compared to control participants using the agonist d- fenfluramine, with ecstasy users showing significantly reduced prolactin and cortisol
responses compared to controls. Gouzoulis-Mayfrank et al (2002) failed to find any
significant effects of d-fenfluramine. However, one of their control groups consisted of heavy cannabis users, which is thought to be an important confound in endocrinological studies of ecstasy users.
McCann et al (1999) also found evidence of possible serotonergic neurotoxic change using another 5-HT agonist, M-chlorophenylpiperazine, which caused significant blunting of both prolactin and cortisol responses in heavy ecstasy users compared to non-using controls. It therefore appears that differences in the studies involving pharmacological challenges are related to the 5-HT activating drug used, with L-tryptophan appearing to be the least sensitive probe for demonstrating brain serotonergic alteration induced by MDMA. None of these pharmacological challenge studies have demonstrated any significant correlation between prolactin responses and the amount of ecstasy used (Price et al, 1989; Gerra et al 1998 & 2000), such that in summary pharmacological challenge techniques seem a somewhat crude and indirect assessment measure of MDMA neurotoxicity in humans.
Kirstie Soar Chapter 1
3. Neurological Assessment
More recently less invasive methods of evaluating the neurotoxic effects of MDMA in
humans have been developed, using measurements of biological markers by in vivo imaging techniques (see Table 1; for a comprehensive review see Reneman et al, 2001). PET studies using the 5-HT transporter ligand McN-5652 have shown decreases in both global and
regional 5-HT transporter binding in ecstasy users, compared with ecstasy-naive controls (McCann et al, 1998) and polydrug controls (Thomasius et al, 2003). This would appear to indicate actual structural changes in the serotonin neurons within the brain. Also the
decreases observed in the 5-HT labelled transporter sites correlated with the degree of
previous MDMA exposure (McCann et al, 1998), indicate dose-related 5-HT neurotoxicity. Further, using the same transporter ligand, Buchert et al (2003) showed that ecstasy users had significantly reduced distribution volume ratios of SERT (presynaptic serotonin transporter) availability in the mesencephalon and thalamus, compared to drug naive controls. However, Gamma et al (2001) did not find any indication of neuronal alterations using the same 5-HT ligand; although this method only used PET to detect possible deviant patterns of rCBF, not SERT densities or availability.
Neuronal activity using PET has also been studied by Obrocki et al (2000). They assessed alterations to the brain cerebral glucose metabolic rate using the ligand 2-[(18)F]-fluro-2- deoxy-D-glucose (FDG). They demonstrated lasting reductions in the metabolic uptake rate within the amygdala, hippocampus and Brodmann's area 11 in ecstasy users, compared to controls. However, no correlations were found between FDG uptake rates and cumulative ecstasy dosage. One further limitation of this study was that FDG PET does not selectively display activity of the serotonergic system; rather it reflects total neuronal activity and thus the possible effects of MDMA on other neurotransmitters in these areas, can not be
established. In addition, it is as yet impossible to conclude whether any disruption to 5-HT or other neurotransmitters, is due to MDMA and/or other drug use.
SPECT has also been used to demonstrate selective serotonin neurotoxicity, using a number of different radioactive ligands; in particular [ 123I]p-CIT and [ 123I]R91150 which are good in
vivo tracers for 5-HT transporters (Reneman et al, 2001). Using these markers, ecstasy users
were shown to have significantly reduced cortical SERT, particularly in the primary sensory cortex, compared to polydrug controls, but there were no correlations between lifetime dose
Kirstie Soar Chapter 1
and reductions in SERT binding (Semple at al, 1999). Such effects on cortical 5-HT receptor densities have been replicated by Reneman and colleagues (2000, 2001a, 2001b), who
demonstrated significantly lower binding ratios in recent ecstasy users compared to controls. This suggests down-regulation of receptors caused by MDMA-induced 5-HT release.
Reductions in receptor densities have also been demonstrated in all areas studied within the cerebral cortex (Reneman et al, 2002), but with no correlation between the level of cortical binding and extent of previous ecstasy use.
These alterations in SERT densities only appear to occur in heavy ecstasy users. In all of the above studies, reported lifetime consumption of ecstasy is above 140 tablets. Reneman et al (2001) only found significant decreases in overall binding ratios in heavy users (who had used over 50 ecstasy tablets) compared to non-ecstasy users, but not between binding ratios of moderate users (who reported use of a maximum of 50 ecstasy tablets) compared with non- users. This could be taken to suggest a possible dose-response effect shown in the previously discussed animal data. However, this could also be indicative of a neurotoxic 'threshold'. Below this threshold neurons can manage or self-protect against (or perhaps recover from) the neurotoxic effects of MDMA, but above this threshold neurons may sustain damage.
Magnetic resonance spectroscopic imaging (1H-MRSI) has also provided evidence for neuronal abnormality in human recreational ecstasy users. Reneman et al (2002) evaluated the MDMA-related alterations in metabolite ratios: N-Acetylaspartate (NAA) / creatine (Cr), NAA/Choline (Cho) and myo-inositol (MI)/Cr ratios (markers associated with neuronal loss or dysfunction) in the frontal cortex of ecstasy users. Neuronal abnormalities significantly correlated with the degree of ecstasy use: the higher the amount of ecstasy exposure the lower the metabolite ratios. Chang et al (1999) also demonstrated metabolic alterations in ecstasy users compared to normal controls using the same method. Cumulative lifetime dose showed significant effects on MI (a glial marker) in the parietal white matter and the occipital cortex of ecstasy users. However, using the same technique, Obergeisser et al (2001) failed to find any group differences in neuronal functioning in the hippocampus, between ecstasy users and controls, although they only assessed a small number of users (5) and also a differing region of the brain to that assessed by Reneman et al (2002), which might account for the difference in findings.
Kirstie Soar Chapter 1
Neuroimaging techniques are not without their criticism, especially when it comes to data interpretation as evidence for MDMA neurotoxicity. The binding specificity of ligands has been questioned, potentially causing an under or overestimation of binding density (see Cole et al, 2003 for further discussion). Also, the loss of markers indicated in these neuroimaging techniques does not necessarily equate to actual cell loss. Certain polymorphisms in the 5-HT transporter gene have shown to have reduced 5-HT transport activity. As such, these possible pre-existing differences in ecstasy users in previous studies are unknown (Kish, 2002).
Whilst there are limitations to neuroimaging studies, the converging line of evidence, using different techniques in conjunction with the preclinical animal data, does allow for a certain level of assessment of the potential effects of ecstasy.
4. Electrophysiological Assessment
Other evidence of MDMA-induced depletion of 5-HT functioning is provided by studies using electrophysiological assessment, involving the auditory evoked potentials and the intensity dependence paradigms. The intensity dependence of auditory evoked potentials is thought to be one index of 5-HT integrity. High intensity dependence has found to be
associated with a low functioning of serotonergic neurotransmission (Hegerl and Juckel, 1993). Croft et al (2001) and Tuchtenhagen et al (2000) have both demonstrated serotonin neuronal alterations via this method in ecstasy users. Ecstasy users exhibited significant increases in amplitude of the tangential N1/P2 source activity with higher stimulus intensities compared to drug-naive and cannabis users, indicating diminished serotonergic activity
specifically in ecstasy users. In addition, Croft et al (2001) demonstrated a significant positive relation between ecstasy users' N1/P2 slopes and total ecstasy consumption independent of cannabis use, suggesting a causative link between ecstasy and 5-HT dysfunction.
Further evidence for dose-related neuronal alterations is provided by Dafters et al (1999). They investigated whether there was a correlation between quantitative EEG variables (Spectral power and coherence) and the level of prior ecstasy use. Reported ecstasy use positively correlated with absolute power in alpha and beta frequency bands and negatively correlated with EEG coherence.
Kirstie Soar Chapter 1
Dopaminergic functioning
Most of the previously discussed studies addressing neurotoxicity have focused on the levels of serotonin within the brain of human recreational ecstasy users. Two further studies
focused primarily on the long-term effects, MDMA exposure may have, on the dopamine system. Gerra et al (2002) investigated dopaminergic function in ecstasy users compared to control subjects. In a pharmacological challenge study using bromocriptine (a specific D-2 receptor agonist), they found a negative correlation between dopamine receptor sensitivity and ecstasy exposure. This suggested possible reduced dopaminergic receptor sensitivity in heavy ecstasy users. Within the same study, there were no significant group differences on prolactin response, but there was a significant difference in growth hormone (GH) responses. Ecstasy users showed significantly reduced GH response compared to controls. However,
such alterations in dopamine could be related to the use of other drugs, which are known to affect dopaminergic neurons. Reneman et al (2002) demonstrated that the sole use of ecstasy was not related to dopaminergic neurotoxicity, but rather that the combined use of ecstasy and amphetamine were associated with reduced dopamine transporter densities. This appears to suggest that MDMA is not associated with human dopaminergic neurotoxicity; but rather it is a selective serotonergic neurotoxin.
Strength of evidence for neurotoxicity in ecstasy users
With the development of in vivo imaging in the human brain, there is now extensive evidence which suggests that MDMA may cause neuronal injury in some recreational ecstasy users. However, these human studies employ a retrospective design and thus evidence is indirect and based on associations. Experimental and/or longitudinal designs are needed to establish
whether there is actually a causal link between neurotoxic changes and MDMA. There are a number of methodological flaws with the neuroimaging studies. Kish (2002) argues that evidence employing the radioligands used to bind to the serotonin neuron, over-rely on one component, that of the serotonin transporter (SERT). He questions the reliability and validity of the SERT measure and whether this is actually proof of brain damage; since it has been established that drug-induced changes in the levels of brain neurotransmitter transporters can occur independently of any changes in the number of serotonin neurones. As such, brain levels of SERT might change following exposure to some drugs independently of any
changes in levels of nerve terminals. It is also suggested that SERT levels can also vary as a
Kirstie Soar Chapter 1
function of oestrogen status, gender and variations in a SERT promoter gene polymorphism, which may be unrelated to the actual number of serotonin neurons. Thus studies involving measurement of brain SERT might be confounded to some extent.
However, there is other evidence, as discussed, which suggests there are alterations in neuronal functioning which have been associated with MDMA exposure, which mirrors findings found in the animal data. The notion that cell loss of markers equates to real 5-HT cell loss, within the human brain can be obtained from post-mortem brain examination and to date there is only one published study that has done this. Kish et al (2000) reported that
striatal (putamen, caudate, nucleus accumbens) levels of serotonin and of its metabolite 5- HIAA were severely depleted by 50 to 80% in the brain of an ecstasy user compared to
controls, but that there were generally normal dopamine concentrations. However, it still can not be determined whether 5-HT depletion was caused by ecstasy use or other polydrug use. Though all methodologies described are flawed, taken together with the animal data there is an increasingly compelling case for the theory of MDMA-induced 5-HT injury in recreational ecstasy users. Thus taken together the evidence strongly supports the earlier animal findings of reductions in brain serotonin in ecstasy users as a result of MDMA-induced neurotoxicity.
Recovery from MDMA neurotoxicity in humans
As outlined above there is strong evidence to suggest that recreational ecstasy use can cause serotonergic injury within the human brain. This also raises the question as to whether
recovery of brain neurons can occur after continued abstinence from the drug or whether such changes are persistent. Theses questions have only been addressed in the last 5 years. Chang et al (1999) did not find a significant relationship between the recent timing of MDMA use and the concentration of any metabolites, nor did Reneman et al (2002). McCann et al (2000) also found no significant correlation between the duration of abstinence from MDMA and the extent of 5-HT transporter binding. However, Semple et al (1999) and Reneman et al (2002) observed a significant positive correlation between SERT binding and the duration of
abstinence; suggesting possible recovery from serotonergic neurotoxic injury over time. Obrocki et al (2000) also focused on the reversibility of PET FDG uptake on brain glucose metabolism and found a correlation between uptake and the time since ecstasy was last ingested; though as mentioned before, PET FDG only reflects total neuronal activity levels
Kirstie Soar Chapter 1
not selective serotonergic activity. The evidence of possible reversibility of MDMA neurotoxicity, is only based on associations using correlational analyses. Other studies
though, have actually looked at group differences between current ecstasy users, former or ex- ecstasy users and controls.
Using SPECT, Reneman et al (2000a and b) compared ecstasy users and ex-ecstasy users who reported using similar amounts of ecstasy but had not used in the last 2 months, and controls. Cortical binding ratios were lower in current ecstasy users compared to ex-users and controls. There was also a significant correlation between cortical binding and duration of abstinence from ecstasy; suggesting possible neuronal recovery in ex-ecstasy users. They also
demonstrated that there was higher cortical binding of the 5-HT ligand [ 123I]R91150 in the ex- ecstasy users compared to controls (though not a significant effect), possibly suggesting an up-regulation of postsynaptic receptors. Reneman et al (2002) replicated these findings in a later study, but only in female ecstasy users. Binding ratios were significantly higher in ex- female users compared to current female users, but not controls; again suggesting that in
several brain regions, MDMA-induced decreases in serotonin transporters could be reversible. This study also suggested a possible gender difference in recovery as this reversal was not observed in the male ecstasy users. Buchert at al (2003) have also indicated the reversibility of MDMA-induced SERT availability as measured by PET. Former users showed levels
close to that of drug-nai've controls in all brain regions assessed. However, using a tryptophan challenge, as an indirect measure of central 5-HT function, Curran & Verheyden (2003)
showed evidence of altered 5-HT functioning in ex-ecstasy users, but not current users, compared to controls. This indicates that neuronal alterations could further develop after cessation of ecstasy use. However, in light of previous research showing possible recovery, it is more than likely that such differences in 5-HT functioning in these users could reflect pre- morbid differences in 5-HT function.
When considering the persistence or reversibility of MDMA-induced neurotoxicity, there is a need for more longitudinal studies like that of Gerra et al (2000) who investigated possible reversibility of changes in the 5-HT system. They found that prolactin rises were
significantly impaired in ecstasy users compared to controls both 3 weeks after discontinuing ecstasy use and after prolonged abstinence (12 months). But in contrast, cortisol rises in ecstasy users were significantly impaired compared to controls at 3 weeks, but were restored after 12 months. Thus, the restored responses of cortisol after 12 months may represent the
Kirstie Soar Chapter 1
expression of an initial recovery of serotonergic functioning after MDMA-induced neurotoxicity.
Whether MDMA leads to irreversible or partly reversible impairment of serotonergic neurons within humans still remains controversial. The current studies do not allow for definite
Kirstie Soar Chapter 2
CHAPTER 2
Possible Functional Consequences of Ecstasy-Induced
Serotonergic Neurotoxicity
Kirstie Soar Chapter 2
The previous chapter highlighted a substantial body of evidence that supports the idea that recreational ecstasy use induces neurotoxic effects on serotonergic neurons and alters brain serotonergic functioning. The fundamental question then, is whether or not this MDMA- induced neurotoxicity results in alterations in human behaviour.
The role of serotonin has been implicated in the regulation of mood (Young et al, 1985), depression (Delgado et al, 1990), anxiety (Garvey et al, 1995), aggression and impulsiveness (Brown et al, 1979; Coccaro, 1989; Reist et al, 1996; Askenazy et al, 2000), sexual activity, appetite (Fernstrom, 1987), sleep (Oswald et al, 1964), pain (Messing et al, 1977; Akunne and Soliman, 1994), circadian and seasonal rhythms (Penev et al, 1995), motor activity
(Loubinoux et al, 2002) and body temperature (Blier et al, 2002). It is also thought to play a role in cognitive processes (Hunter, 1988), although the place of 5-HT in cognition is poorly understood. However, it has been proposed that serotonin may play an important modulating role in memory and attention and so it is possible that extreme deviations of 5-HT activity could result in biases in cognitive processing. Evidence to date suggests that 5-HT is
involved in learning, visuo-spatial memory, visual discrimination, associative functions and aspects of planning and general memory.
The consequences, therefore, of alterations in serotonergic functioning in recreational ecstasy users are expected in these psychological and behavioural domains, which are related to
serotonergic processes. This current chapter summarises the growing number of research reports which lend support to this notion, in particular concerning psychiatric,
psychobiological, and also cognitive effects in recreational ecstasy users. The idea that these psychological effects are associated with altered serotonergic functioning is also discussed, along with a discussion regarding evidence concerning the reversibility or permanence of these effects.
Long-term psychopathological consequences associated with ecstasy use
The first evidence which indicated that ecstasy might lead to chronic psychiatric symptoms came from individual case studies, where psychiatric complaints were reported to have appeared to develop in the context of ecstasy use (see Table 2 for a summary of case reports from the last 15 years). These clinical reports suggest that certain individuals appeared to have developed psychopathological symptoms, which manifested in a range of psychiatric
Kirstie Soar Table 2: A summary of clinical case studies, where ecstasy appears to be a prominent feature. Chapter 2 Reference Symptom/ Disorder
Age/ Sex
Psychiatric History Psychiatric illness among 1" degree relatives Ecstasy Use Other drug use Teggin(1992) Cohen & Cocores (1997)
Conen(1996) McGuire&Fahy<(1991) McGuire&Fahy(1991) Cassidy&Ballard(1994) Series
et
al
(1994)
Keenanetal(1993) Bone
et al (2000) Ellis & Schimmel (1989)