Rifampicin Mediated Cell Death in Staphylococcus aureus
By Seth Alexander The Brian Conlon Lab
Senior Honors Thesis Department of Biology
University of North Carolina at Chapel Hill
April 17th, 2018
Abstract
Antibiotic tolerance and resistance create significant barriers to pharmacologic treatment of bacterial infections. Rifampicin, a commonly used rifamycin, is a bactericidal antibiotic that binds to RNA polymerase in the cell and inhibits transcription elongation after the synthesis of a strand of two or three nucleotides. It is not understood how this ultimately leads to cell death as opposed to stasis. Previous studies have shown that the metabolic state of S. aureus can alter the degree of tolerance to antibiotics but have not provided information regarding how this relates to the mechanism by which rifampicin induces cell death. Rifampicin mediated cell death is potentially the result of a futile cycle (continued synthesis of short strands of nucleotides), which could exhaust the cell of available nucleotides. Alternatively, DNA damage could be caused by the inhibition of transcriptional enzymes causing irreversible collapse of the cell’s nucleoid and/or double stranded DNA breaks.
regards to the metabolic state of the cell, it was found that S. aureus cultured in glucose containing media exhibits an increase in resistance to rifampicin when compared to media containing amino acids as an alternative carbon source.
These results suggest that rifampicin mediated cell death is not the result of a futile cycle nor improper nucleoid condensation. These findings leave open the possibility that lethal, double-stranded DNA breaks are being caused by collision between inhibited transcriptional enzymes and replication enzymes. This possibility should be the subject of future study along with studies pertaining to how metabolism of glucose may alter the state of the cell so as to increase resistance to rifampicin. Determination of the mechanisms leading to rifampicin mediated cell death could aid in the development of more robust antibiotics and treatments which could more effectively combat tolerant and resistant strains of bacteria.
Introduction
Clinical Background Staphylococcus aureus is a common commensal organism that lives in the nares and on the skin. In some people, particularly the immunocompromised or chronically ill, S. aureus infection can lead to skin infections, osteomyelitis, endocarditis, and other deep tissue infections[1]. Several S. aureus strains have developed antibiotic resistance resulting in Methicillin-resistant S. aureus and Vancomycin-resistant S. aureus. Understanding the mechanisms by which antibiotics act upon the cell provides information on how resistant strains arise and allows for development of more efficacious antibiotics.
selection pressures act on underlying genetic variation in a bacterial population and favor proliferation of resistant mutants[2]. However, tolerance is an innate mechanism by which the bacteria are able to tolerate antibiotic challenges without acquisition of genetic resistance[3]. It has been shown that properties such as the metabolic activity of the cell contribute to the degree of antibiotic tolerance[4].
Futile Cycle Hypothesis Rifamycins are bactericidal antibiotics that bind to RNA polymerase (RNAP) in the cell. Rifampicin, a rifamycin, is a transcription elongation inhibitor. After binding to RNAP, rifampicin allows RNAP to synthesize a transcript of two or three nucleotides before terminating the process[5]. If this process continues consistently, it would produce a futile cycle and could potentially deprive the cell of free nucleotides. Without available nucleotides to contribute to transcription or replication, the cell would be susceptible to death. Rifabutin, a transcription initiation inhibitor, will be used to investigate this hypothesis. By inhibiting initiation, there is no possibility of generating a futile cycle. If rifabutin results in bacteriostatic effects or lesser bactericidal effects than rifampicin, it would suggest that a futile cycle is occurring and contributing to cell death. This would serve as a foundational finding upon which to further investigate a futile mechanism.
to oxidative stress[7]. If decondensing the nucleoid is the mechanism of cell death induced by rifampicin, overexpression of dps should condense the nucleoid and thus protect it from further decondensation, which would increase tolerance to rifampicin. Similarly, in a PerR mutant, the nucleoid should be more compact due to constitutive expression of dps and thus be more tolerant to rifampicin mediated death.
Another hypothesis is that rifampicin causes cell death by causing double stranded DNA breaks. If RNAP is prevented from terminating its connection with the chromosome, it is possible that RNAP could collide with DNA polymerase trying to replicate the chromosome. This collision could subsequently result in double stranded DNA breaks. By breaking the circular bacterial chromosome, the cell would not be able to maintain vital functions and would likely die. Detection of double stranded DNA breaks via quantitative analysis or constitutive expression of DNA repair mechanisms in a mutant strain would reveal whether such breaks occur with higher frequency in antibiotic treated cells. This can be the subject of future studies.
Materials and Methods
Unless otherwise stated, all experiments were performed in biological triplicates; indicated values are averages of these triplicates. A result was determined to be significant if a two-tailed t-test yielded a p-value < 0.05.
cultures were grown in chemically defined media (CDM) which was prepared as specified by Vitko and Richardson[8]. Broth cultures were incubated while shaking at 225 rpm and 37 °C.
Prior to the author’s involvement on this project, a genetically modified strain of S. aureus was constructed. To construct this mutant, a low copy plasmid containing a xylose inducible promoter was transduced into HG003. When working with this strain, the cells were exposed to xylose for 30 minutes prior to being challenged with any antibiotic.
A perR mutant was constructed by transducing a plasmid containing an erythromycin resistant cassette from the Nebraska transposon library into homologous regions within the perR genetic sequence. Donor cells were grown on a BHI slant overnight and then resuspended in 1 mL of Tryptic Soy Broth (TSB) and 5 mM CaCl2 to facilitate phage binding and lysis. Afterwards, 100 µL of transducing bacteriophage suspension (80ɑ and ɸ11), 10 µL of cell suspension, and 4 mL of 0.5% TSA were combined, plated on TSA and 5 mM CaCl2, and incubated at 37 °C overnight. The following day, the loosened agar was combined with 3 mL of TSB and centrifuged before being separated with a 0.45 µm filter. The recipient strain was grown on a BHI slant overnight and then resuspended in 1 mL of TSB and 5 mM CaCl2. A 500 µL aliquot of cells were transferred into 1.5 mL of TSB and 5 mM CaCl2 before adding 500 µL of filtered phage. This solution incubated while shaking for 20 minutes before being centrifuged and the supernatant disposed of. The pellet of cells was then resuspended in 1 mL of ice-cold sodium citrate to chelate the calcium. The cells were then plated on TSA, sodium citrate (50 µg/ mL), and erythromycin (10 µg/mL) and incubated at 37 °C.
desired media. Stationary phase S. aureus was diluted 1:1000 in each media and 100 µL of culture was added to each antibiotic well and to the positive control wells. Plates were sealed with breathe easier® strips (Fisher) to allow aeration and prevent contamination. Plates were incubated at 37 °C for 24 hours. The lowest concentration at which there was no visible growth was considered the MIC for each of the tested media.
Each individual image was imported into the Java based photo editing software, Fiji. Each fully visible S. aureus cell was circled using the in-software drawing tool and a Wacom Intuos Photo Pen and Touch Digital Photo Editing tablet. Cells were circled to include all cellular area within the peptidoglycan cell wall. The nucleoid was also circled. Using the scale bar provided on each microscopy image, the area of the cell and nucleoid were determined using Fiji’s measure function. A ratio of nucleoid area to cell area was then calculated for each of the cells analyzed. The average ratio from rifampicin treated cells (N=47) was then compared to the average ratio for untreated control cells (N = 50).
Antibiotic Challenges Cultures were grown to mid-exponential phase. A time point to determine initial colony forming units per mL (CFU/mL) was taken and then the maximum achievable serum concentration (Cmax) of each antibiotic was added to each culture. Rifampicin (Cmax = 10 µg/mL)[10] was tested in the presence of trimethoprim (Cmax = 0.8 µg/mL)[11] to prevent growth of rifampicin resistant cells. Rifabutin was challenged at the Cmax concentration of 0.46 µg/mL[12].
Results
Figure 1. Antibiotic challenge of S. aureus with rifampicin and rifabutin exhibits no difference in tolerance. Mid-exponential phase S. aureus HG003 cells were challenged with trimethoprim (0.8 µg/mL) and rifampicin (10 µg/mL) or rifabutin (0.46 µg/mL) at T0 and then surviving cells were enumerated at T18 and T24. Error bars have been omitted where the standard deviation does not exceed the height of the data point symbol.
Figure 2. The nucleoid of rifampicin treated S. aureus appears larger than that of control cells in transmission electron microscopy (TEM). When reviewing TEM images of control cells (A) and rifampicin treated cells (B), the nucleoid appears more diffuse in the antibiotic treated cultures.
Figure 3. The nucleoid to cell area ratio is almost two-fold higher in rifampicin treated cells.
Quantitative analysis of TEM images of control (N = 46) and rifampicin (N=50) treated S. aureus cells was performed using Fiji photo editing software to quantify the ratio of nucleoid area to total cell area.
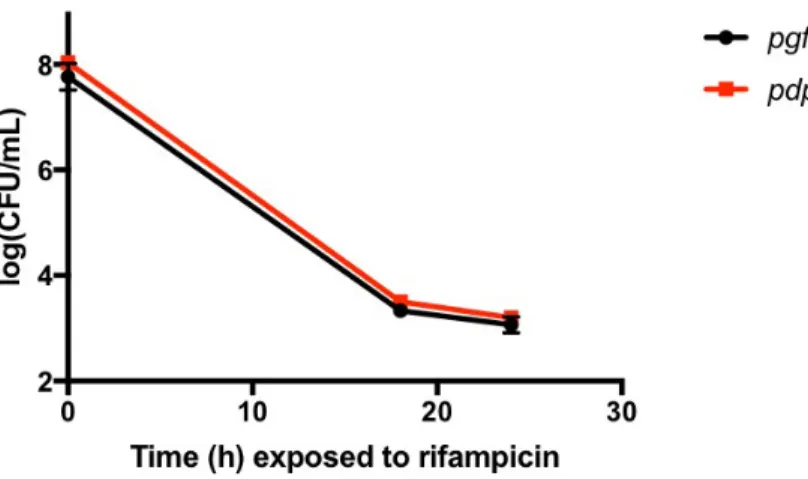
Figure 4. Overexpression of dps does not alter antibiotic tolerance in S. aureus. S. aureus strain HG003 harboring plasmids pEPSA5::igfp (pgfp) or pEPSA5::dps (pdps) were grown to mid-exponential phase and challenged at T0 with rifampicin (10 µg/mL) and trimethoprim (0.8 µg/mL) and surviving cells were enumerated at T18 and T24. Error bars have been omitted where the standard deviation does not exceed the height of the data point symbol.
Figure 5. Disruption of perR does not alter antibiotic tolerance in S. aureus. S. aureus strain HG003 or HG003 perR::erm were grown to mid-exponential phase and challenged at T0 with rifampicin (10 µg/ mL) and trimethoprim (0.8 µg/mL) and surviving cells were enumerated at T18 and T24. Error bars have been omitted where the standard deviation does not exceed the height of the data point symbol.
cultures grown in glucose containing CDM, the MIC was found to be eight-fold higher (p = 0.0317). This suggests that the cells possess some additional form of innate resistance to rifampicin when catabolizing glucose as a carbon source as opposed to solely amino acids.
Table 1. The minimum inhibitory concentration (MIC) of S. auerus cultured in CDM is elevated.
Media Rifampicin MIC (µg/mL)
Mueller-Hinton Broth 0.006
Chemically Defined Media 0.048
When comparing the MIC of S. aureus in non-glucose containing MHB to glucose containing CDM, it
can be seen that the MIC is significantly elevated in the CDM (p = 0.0317).
Discussion
Based on the above data, the mechanism of antibiotic mediated cell death in cultures treated with rifampicin cannot yet be determined. The data suggest that rifampicin does not induce a futile transcription elongation cycle. While nucleoid collapse occurs in rifampicin treated cultures, evidence has not yet been found to support that the enlarged nucleoid contributes to cell death. Future experimentation should seek to fully elucidate the mechanism if the collapsed nucleoid contributes to cell death and to investigate the eight-fold increase in MIC that S. aureus exhibits when cultured in the presence of glucose.
an artifact of the images due to cell death. This could be accomplished utilizing high resolution microscopy images of the cells in real time or TEM images of cells removed from the antibiotic at a number of earlier timepoints. Additionally, construction and rifampicin challenge of a mutant that cannot synthesize dps and therefore cannot condense its nucleoid, would provide an additional piece of data on which to base this conclusion.
Further experimentation should be done to understand why the cells exhibited varying degrees of resistance in different media. Based on the data presented above, it appears that the metabolic state of the bacteria when glucose is present provides some level of intrinsic resistance to rifampicin. This finding can be verified utilizing CDM that lacks glucose to minimize confounding variables. If the result remains consistent, this would indicate that the catabolic pathway in which S. aureus breaks down glucose varies considerably from that in which the cells are catabolizing amino acids as in MHB. This would have far reaching implications for the field when experimenting with rifampicin given that a large portion of experimental cultures are grown in glucose-lacking media such as MHB. Additionally, this innate resistance would be significant in the in vivo environment where glucose is readily available and would have far reaching clinical implications.
Acknowledgements
I would like to thank Dr. Brian Conlon and Dr. Sarah Rowe-Conlon for their direct advice and oversight in the completion of this project. I would also like to thank Lauren Radlinski and Nikki Wagner for their assistance and advice. I would like to thank the UNC School of Medicine Microscopy Services Labratory for providing the TEM images and protocol referenced above. I would like to thank Dr. Amy Shuab-Maddox for her advice regarding the composition and presentation of this thesis. I would finally like to thank the Department of Microbiology and Immunology at UNC School of Medicine for providing the internal funds to Brian Conlon which made this project possible.
References
1. Conlon, B. P. (2014), Staphylococcus aureus chronic and relapsing infections: Evidence of a role for persister cells. BioEssays, 991–996. doi:10.1002/bies.201400080
2. Antibiotic / Antimicrobial Resistance. (2017, September 19). Retrieved December 06, 2017, from https://www.cdc.gov/drugresistance/about.html
3. Brauner, A., Fridman, O., Gefen, O., & Balaban, N. Q. (2016). Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nature Reviews Microbiology, 320-330. doi:10.1038/nrmicro.2016.34
5. Unissa, A. N. (2017). Molecular mechanisms of action, resistance, detection to the first-line anti tuberculosis drugs: Rifampicin and pyrazinamide in the post whole genome sequencing era. Tuberculosis. 96-107. doi:https://doi.org/10.1016/j.tube.2017.04.008 6. Ohniwa, R. L., Morikawa, K., Takeshita, S. L., Kim, J., Ohta, T., Wada, C., & Takeyasu, K.
(2007). Transcription-coupled nucleoid architecture in bacteria. Genes to Cells,
1141-1152. doi:10.1111/j.1365-2443.2007.01125.x
7. Ushijima, Y., Ohniwa, R. L., Maruyama, A., Saito, S., Tanaka, Y., and Morikawa, K. (2014). Nucleoid compaction by MrgAAsp56Ala/Glu60Ala does not contribute to staphylococcal cell survival against oxidative stress and phagocytic killing by macrophages. FEMS Microbiology Letters. 144-151. doi:10.1111/1574-6968.12598
8. Vitko, N. P. and Richardson, A. R. 2013. Laboratory Maintenance of Methicillin-Resistant Staphylococcus aureus (MRSA). Current Protocols in Microbiology. 28:C:9C.2:9C.2.1– 9C.2.14.
9. Reynolds, E.S. 1963. The use of lead citrate at high pH as an electron opaque stain in electron microscopy, J. Cell Biol. 17, 208.
10. Barcia-Macay M, Seral C, Mingeot-Leclercq M-P, Tulkens PM, Bambeke FV. Pharmacodynamic Evaluation of the Intracellular Activities of Antibiotics against Staphylococcus aureus in a Model of THP-1 Macrophages. Antimicrobial Agents and Chemotherapy. 841-851. doi:10.1128/aac.50.3.841-851.2006.