Bryan
L
Roth
ChemogenetictechnologiessuchasDesignerReceptors ExclusivelyActivatedbyDesignerDrugs(DREADDs)arewidely usedtoremotelycontrolneuronalandnon-neuronalsignaling. DREADDsexistformostofthecanonicalGprotein-coupled receptorsignalingpathways,andprovideasyntheticbiology platformusefulforelucidatingtheroleofneuronalsignalingfor brainfunction.Here,afocusedreviewisprovidedthatshows howrecentinsightsobtainedfromGPCRstructuralstudies informourunderstandingofthesechemogenetictoolsfroma structuralperspective.
Address
DepartmentofPharmacologyandDivisionofMedicinalChemistryand
ChemicalBiology,UniversityofNorthCarolina,ChapelHillSchoolof
Medicine,ChapelHill,NC27514,UnitedStates
Correspondingauthor:Roth,BryanL([email protected])
CurrentOpinioninStructuralBiology2019,57:9–16
ThisreviewcomesfromathemedissueonEngineeringanddesign:
syntheticsignaling
EditedbyAndreasMo¨glichandHaraldJanovjak
ForacompleteoverviewseetheIssue andtheEditorial
Availableonline25thFebruary2019
https://doi.org/10.1016/j.sbi.2019.01.016
Introduction
Gprotein-coupledreceptors(GPCRs)—withmorethan 800 members — represent the largest family of mem-brane proteins in the human genome [1]. GPCRs also represent the single largest family of therapeutic drug targets in the druggable genome [2], with 20–30% of approveddrugshaving GPCRsastheirmajormolecular target [3,4]. GPCRs are expressed in essentially every majororganinthebody,withapproximately80%being expressed in the brain [5]; many GPCRs have their highestlevelofexpressioninnervoustissue[5].GPCRs transduce their signals via binding to and activating hetereotrimericGproteins;Gproteinscoupletovarious downstream effectors to modulate second messengers including cAMP, inositol trisphosphate (IP3), calcium (Ca2+)andothers[3].GPCRsalsocansignalviaarrestin proteins [6] which bind to activated GPCRs following phosphorylation by G protein receptor kinases (GRKs) [7].ArrestinsalsoarrestG-proteinsignalingandinteract with clathrintopromote endocytosis[8].
Withinneurons, GPCRs,viaGproteinactivation, regu-late neuronal excitability [9], vesicle release [10], ion channel activity [11,12], and a variety of intracellular secondmessengers[seeRef.[13]forreview].Howthese alterations in neuronal signaling and firing ultimately result in various brain processes such as perception, cognition,emotion,motivation,andsoonisyetunclear. Someyearsago, FrancisCrickproposedthatinorderto ultimately understandthe neuronalbasisof thesebrain activities[14,15]wewouldneed:
“ ...to identifythemanydifferenttypesof neu-rons in the cerebral cortex and other partsof the brain. One of the next requirements...is to be abletoturnthefiringofoneormoretypesofneuron on and off in the alert animal ...One way-out suggestion is to engineer these neurons so that when one of them fires it would emit a flash of lightof aparticularwavelength...”[15]
The past decade has seen remarkable progress on this frontandessentiallyalloftheitemsinCrick’s‘wishlist’ are in place. First, RNA-seq based technologies have begun cataloging the many different types of neurons [16] and large numbersof engineered mouse lines are availabletoultimatelyprovidegeneticaccessto individ-ual types of neurons [17,18]. Second, optogenetic [19,20] and chemogenetic [21,22] technologies have providedtools‘toturnthefiringofoneormoretypesof neurononandoffinthealertanimal’.Finally,genetically encodedcalcium[23]andvoltage[24]sensorsare avail-able to image neurons that ‘emit a flash of light of a particular wavelength’ [25]and bevisualized with fiber optics [26]andminiaturemicroscopes [27].
signaling cascade for a particular process (Figure 1). Although these pioneering chemogenetic technologies were useful for archetypical proof-of-concept studies [31,32], they were notwidely adopted.To circumvent problemsinherentwiththefirst-generationchemogenetic tools(lackofinertligandsandhighbasalactivityof recep-tors),mylabdevelopedDREADDs(DesignerReceptors ExclusivelyActivatedbyDesignerDrugs)[21],whichare nowroutinelyusedbyneuroscientistsandothersto modu-late neuronal activity and cellular signaling [22]. DREADDsarealsousedtoclarifyhowGPCRsignaling processesareimportantformediatingnormalandabnormal physiology[33,34,35,36].
DREADDs[22]areGPCRsthathavebeenengineeredto: (a)lackappreciableresponsestotheirendogenousligand;(b) haveminimalbasal (constitutive) activityand(c)beactivated by a pharmacologically inert ligand. Many types of DREADDscurrentlyexist,includingthosebasedon mus-carinic[22],k-opioid[37]andtheFFA2fattyacidGPCRs [38]. All of these DREADDs activate the canonical G proteinstowhichtheyarecoupledafterstimulationbytheir otherwise inert cognate ligand. Thus, the M2 and M4-DREADDs(hM2DiandhM4Di,respective)activate Gi-familyproteins,M1-DREADDs,M3-DREADDsand M5-DREADDs (hM1Dq,hM3Dq and hM5Dq respectively)
activateGq/11.Thek-opioidDREADD(KORD)activates Gi-familyreceptors,whiletheFFA2DREADDactivatesGi andGqfamilyGproteins.AparticularlyusefulDREADD hasbeendeveloped,inwhichthehighlyconservedarginine in the conserved ‘DRY’motif essential for receptor activation [3,39,40]ismutated.This particularmutation—R3.50L— yields an M3-(R3.50L)-DREADD that is able to recruit arrestin, but can no longer activate G protein signaling [41].ThisM3-(R3.50L)-DREADDcanbeusedto deter-minetheroleofarrestinsignalingincellularprocesses.
DREADDscanbeexpressedviavirallymediated transduc-tion[42]orcanbegeneticallyencodedinessentiallyanycell type[43,44]andactivatednon-invasivelyviadrug-likesmall molecules[45]torevealthephysiologicalconsequencesof GPCRsignalingindefinedcells.Accordingly,DREADDs provideasyntheticbiological platformforselectivelyand non-invasively modulating GPCR signaling. Here, I will summarizeemergingstructuralinsightsintotheactionsof DREADDsthatpromisetotransformandextendthe use-fulnessofthischemogeneticplatform.
Structural
insights
into
muscarinic
DREADD
actions
Thefirst DREADDswerebasedonhuman muscarinic acetylcholine receptors. These muscarinic DREADDs
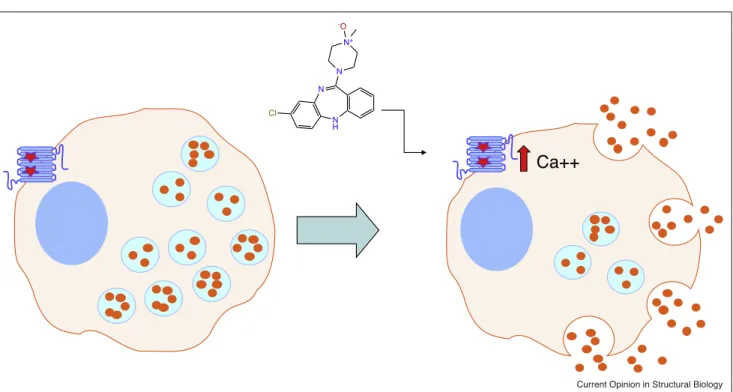
Figure1
N
N
N H
N+
-O
Cl
Ca++
Current Opinion in Structural Biology
DREADD-basedchemogenetictoolsmodulatecellularfunction.
ShownisahypotheticalpancreaticbetacellwhereinaGq-coupledDREADDcanbeactivatedbyCNOtoinduceinsulinrelease(orangecircles).
RedstarsshowtheapproximatelocationsoftheY3.33CandA3.56GmutationswhichrendertheM3-muscarinicreceptorinsensitiveto
created by directed molecular evolution in yeast [22,46]tobeactivatedbytheinertclozapine metabo-lite clozapine-N-oxide (CNO). To date, muscarinic DREADDs are the most widely used chemogenetic tools in the neurosciences. Two of these — hM2Di and hM4Di — inhibit neuronal activity [22] via Gb/
g-mediatedactivationofhyperpolarizingGIRKchannels and inhibition of synaptic release of neurotransmitters [47]. The Gq-coupled DREADD (hM3Dq) is most frequently used to enhance neuronal activity [48], via Gq-mediateddepolarizationandmodulationofion chan-nel activity [44]. When combined with Cre-responsive viruses and genetically encoded Cre-recombinase expressedindiscreteneurons,DREADDs canbeused toremotelycontrolneuronalactivityindefinedneurons and,consequently,modulateanumberofbehaviorsand physiologicalresponses [49,44].
Althoughwidelyused,theseDREADDsarenotwithout liabilities.Thus,forinstance,CNOmaybemetabolized toclozapineatlowlevelsinmice[50]andrats[51]andtoa greaterextentinguineapigsandhumans[52].As cloza-pineisanantipsychoticdrugwithhighaffinityforalarge number of neurotransmitter receptors [53], such a con-versionmaybeproblematicandvariouscontrolsarenow recommended when using CNO [see Ref. [44] for review].Alternatively,CNOanalogswithimproved bio-availabilityandwithoutmetabolicliabilitiessuchasC21 and perlapinecanalsobeused[45,54].Going forward, novel DREADD chemogeneticactuators having differ-ent chemotypes with enhanced bioavailabilty will be useful.
the numbering system developed by Ballesteros and Weinstein [55]. Thus, for the muscarinic DREADDs the conserved amino acids of interest are Y3.33 in TM3andA5.46inTM5(numberedaccordingto Balles-teros and Weinstein [55]). When these are mutated to Y3.33C and A5.46G they yield muscarinic receptors insensitive to the endogenous ligand acetylcholine and potently activatedbyCNO[22](Figure 2aand e).As these particular mutations were arrived at via several cycles of directed evolution and employed unusual [non-conservative]substitutions(e.g.Y!CandA!G) itwas notimmediatelyclearwhytheseparticular muta-tionswereessentialforthedesiredchemicaland biologi-calphenotype.
Several years after the invention of the muscarinic DREADDs,bothinactive[56]andactive-statestructures of the M2 muscarinic receptor were published [57]. Figure 2 also shows an active state structure of the wild-type M2 muscarinic receptor (2D [57]), a model of anactive state hM2Di-DREADD(Figure 2b) and a summaryofkeytransitions(Figure2c).Althoughitisnot entirelyevidentfromthestructurewhythesemutations transformCNOfromaweak antagonistintoanagonist, ourrecentstudiesprovideaclue.Thus,wefoundthata similarA5.46Gmutationofthe5-HT2Bserotonin recep-tortransformstheantagonistmethysergideintoapotent agonist [58]. This is accomplished by providing bulk-tolerancefortheN-methylgroupofmethysergide,which allows for the inward movement of TM5 required for stabilizingtheactive state.Presumably,asimilar transi-tion in the hM2Di-DREADD (Figure 2b–d) is key for
Figure2
hM2Di hM2Di 4MQS hM3Dq
MODEL
2 Å hM2Di
INACTIVE ACTIVE
(a) (b) (c) (d) (e)
Current Opinion in Structural Biology
StructuralinsightsintoDREADDactions.
ShownaresnakeplotsoftheM2(a)andM3(e)[72]muscarinicreceptorswiththelocationsoftheY3.33CandA3.56Gmutationshighlightedin
red.Panel(d)showstheactivestatestructureoftheM2muscarinicreceptorwhile(b)and(c)showamodeloftheM2-DREADDandthe
accommodating bulky constituents of CNO, although direct structural studies will ultimately be essential to determiningthisfor certain.
Structure-inspired
design
of
a
k
-opioid
DREADD
The first DREADDs were developed using an unbi-ased, directed evolution approach and, although suc-cessful for many applications, they suffered from the inability to ‘multiplex’. To be able to use synthetic biological approaches to separately activate distinct GPCR signaling cascades sequentially or simulta-neouslyinthe sameneuronswouldsolve the problem ofmultiplexing.Withtheexplosioninstructural infor-mation related to GPCRs [59] we set out to use this information to design a new DREADD. We choseas ourtemplate the k-opioidreceptor(KOR) forwhicha high resolution structure was available [60] and for whichNMR-basedstructuralinformationwasavailable forthe activestate[61].Wehadpreviouslydiscovered that the non-nitrogenous natural product salvinorin A wasapotentandselectiveKORagonist[62]andthatit did not rely on the conserved Asp3.32 found in the bindingpocket ofmanybiogenicaminereceptors [63] forbindingoractivation.Indeed,SalvinorinAbinding and functional potency were enhanced by a D3.38N mutation while binding of the native peptide ligand dynorphin A(1–17) wasabolished[63].
Accordingly,wecreatedaD3.38Nmutantandfoundthat notonly wasSalvinorin A’spotencyenhanced,but that therelativelyinertmetabolite SalvinorinB(SALB)was transformed into a potent agonist [37]. In a series of studies,wewereabletoshowthatSALBwasinertatthe tested doses in vivo but was able to afford potent and
efficacious neuronal silencing in vivo in genetically defined neurons [37]. We dubbed this the k-opioid receptor DREADD(KORD) and note thatit has been usedwidely in theneurosciences to silence genetically identified neurons [64–66].Recentstructural studies of theactiveKORhaverevealedapotentialmechanismfor SALB’spotencyatKOR[40].
Inaccordancewithanewactive statestructureof KOR [40], we found that the epoxymorphinan-derivative MP1104 bound to KOR at some distance (2.6A˚ ) from the highly conserved D3.38 (Figure 3b). The D3.38A mutationofthisresiduehadamodesteffectonbindingof MP1104, although it abolishes binding of all known endogenous KOR ligands [37]. The D3.38N mutation ispredictedtoresultinamovementof3A˚ outwardfrom the binding pocket due to repulsive forces with the positivelychargedgroupsofmorphinansandopioid pep-tides. Such a movement ispredicted, based on simula-tions, to enhance SALB binding by desolvation [37] thereby providing a molecular mechanism for SALB’s remarkable efficacy at KORD. The D3.38N mutation alsoprovides repulsive energyto hinder thebinding of endogenousopioidpeptides,whichareknowntointeract with this residue in opioid receptors for high affinity binding[61,67]in theactive state.
Future
potential
developments
DREADDsandotherchemogenetictools[68]alongwith optogenetictechnologies[69]havetransformed neurosci-enceandotherdisciplines.Thecurrently available che-mogenetic and optogenetic technologies provide many orthogonal approaches to manipulate cellular signaling andneuronalfiring.Withthecurrentrevolutionoccurring in membrane protein structural determination via
Figure3
D3.32
D3.32N
(a) (b) (c)
6B73
Current Opinion in Structural Biology
Structuralfeaturesofthek-opioidDREADD.
Shownin(a)istheinactivestatestructureofthek-opioidreceptorindicatingthelocationofD3.32showninsphererepresentation.(b)showsa
close-upofthebindingpocketoftheactivestatek-opioidreceptorhighlightingthechangeinrotomerofD3.32frominactive(purple)toactive
(green).(c)showsamodelofthek-opioidDREADDwhereintheD3.32Nmutatedresidueispushedoutofthebindingpocket,freeingupenergyof
crystallography and cryo-electronmicroscopy, the fields are poised to deliveranewgeneration of toolsinspired anddesignedwithhighresolutionstructuralinsights(see Refs.[37,70]forexamples).Combininghighresolution structures with automated docking, for instance, could provide novel chemogenetic actuators for engineered receptors, as has been done for native GPCRs [71] (Figure 4).
Inaddition todevelopingnovelchemogeneticactuators andnewDREADDs,itcontinuestobeusefultoengineer GPCRstoactivatedesignedsignalingnetworks.Thus,for instance, DREADDsareavailablefor Gi[22,37],Gq
[22],Gs[33]andarrestin[41]signaling,althoughnone are available that are selective for G12/13, gustducin, transducin or Golf. As well, although the Gi and Gs-selective DREADDs are currently available, they also mobilize arrestin to at least some extent, and to have thosethatdonotinteractwitharrestinatall(ashasbeen done for a Gq/11-DREADD [36]) will be useful for interrogatingarrestin-dependentandindependent path-waysincells andintactanimals.
Conclusion
Fromtheprecedingexamples,itshouldbeclearthatthe field of chemogenetics is maturing, and that, when
N H N N N
N
!
ON H2N
N NH2
O
N
N N H N S
O N
Cl N N
N
O O
Cl N N N O O N
O N
High resolution structures identify potential selectivity motifs
Docking millions of compounds against mutant receptor to identify
chemogenetic actuators N
O O N N S
O N
O HO
O O
LECL2
S5.42
DRD4/Nemonapride
DRD3
D3.32
Y7.43
F2.61V
L3281F
Current Opinion in Structural Biology
Computationalapproachesforstructure-guideddiscoveryofnovelchemogeneticactuators.
InthetoppanelisshownacomparisonoftheD3-(purple)[73]andD4-(green)[71]dopaminereceptorstructureshighlightingtheselectivityfilter
whichprovidesatemplateforlarge-scaledockingcampaigns[71].Thelowerpanelillustrateshowasimilarprocesscouldbeusedonamutant
ofspecialinterest
ofoutstandinginterest
1. FredrikssonR,LagerstromMC,LundinLG,SchiothHB:The G-protein-coupledreceptorsinthehumangenomeformfive mainfamilies.Phylogeneticanalysis,paralogongroups,and fingerprints.MolPharmacol2003,63:1256-1272.
2.
HopkinsDiscov2002,AL,Groom1:727-730.CR:Thedruggablegenome.NatRevDrug
Aseminalpaperwhichforthefirst-timecategorizedgenesinthegenome
basedontheirpotentialabilitytobetargetdbysmallmoleculessuchas
drugs.
3. WackerD,StevensRC,RothBL:HowligandsilluminateGPCR molecularpharmacology.Cell2017,170:414-427.
4. RothBL,IrwinJJ,ShoichetBK:DiscoveryofnewGPCRligands toilluminatenewbiology.NatChemBiol2017,13:1143-1151.
5.
Regardprotein-coupledJB,SatoIT,receptorCoughlinexpressionSR:Anatomical.Cell2008,profiling135:561-571.ofG
Thefirstcomprehensivemap ofGPCR expression invariousmouse
tissueswhichcontinuestobehighlyuseful.
6. LuttrellLM,FergusonSS,DaakaY,MillerWE,MaudsleyS,Della RoccaGJ,LinF,KawakatsuH,OwadaK,LuttrellDKetal.: Beta-arrestin-dependentformationofbeta2adrenergic receptor-Srcproteinkinasecomplexes.Science1999,283:655-661.
7. DeWireSM,AhnS,LefkowitzRJ,ShenoySK:Beta-arrestinsand cellsignaling.AnnuRevPhysiol2007,69:483-510.
8. GoodmanO,KrupnickJ,SantiniF,GurevichV,PennR,GagnonA, KeenJ,BenovicJ:b-arrestinactsasaclathrinadaptorin endocytosisoftheb2-adrenergicreceptor.Nature1996,
383:447-450.
9. AndradeR,MalenkaRC,NicollRA:AGproteincouples serotoninandGABABreceptorstothesamechannelsin hippocampus.Science1986,234:1261-1265.
10. BlackmerT,LarsenEC,BartlesonC,KowalchykJA,YoonEJ, PreiningerAM,AlfordS,HammHE,MartinTF:Gprotein betagammadirectlyregulatesSNAREproteinfusion machineryforsecretorygranuleexocytosis.NatNeurosci 2005,8:421-425.
11. KrapivinskyG,KrapivinskyL,WickmanK,ClaphamDE:Gbeta gammabindsdirectlytotheGprotein-gatedK+channel, IKACh.JBiolChem1995,270:29059-29062.
12. KrapivinskyG,GordonEA,WickmanK,VelimirovicB, KrapivinskyL,ClaphamDE:TheG-protein-gatedatrialK+ channelIKAChisaheteromultimeroftwoinwardlyrectifyingK (+)-channelproteins.Nature1995,374:135-141.
13. RoganSC,RothBL:Remotecontrolofneuronalsignaling. PharmacolRev2011,63:291-315.
14. CrickFH:Thinkingaboutthebrain.SciAm1979,241:219-232.
15.
CrickPhilosF:TransTheRimpactSocLondofmolecularBBiolScibiology1999,354on:2021-2025.neuroscience.
AlongwithRef.[14]representtheblueprintonhowtousemolecular
technologiestodeconstructthebrain.
16. ZeiselA,HochgernerH,LonnerbergP,JohnssonA,MemicF,van derZwanJ,HaringM,BraunE,BormLE,LaMannoGetal.:
Moleculararchitectureofthemousenervoussystem.Cell 2018,174:999-1014e1022.
17. GongS,ZhengC,DoughtyML,LososK,DidkovskyN, SchambraUB,NowakNJ,JoynerA,LeblancG,HattenME, HeintzN:Ageneexpressionatlasofthecentralnervous systembasedonbacterialartificialchromosomes.Nature 2003,425:917-925.
18. DaigleTL,MadisenL,HageTA,ValleyMT,KnoblichU,LarsenRS, TakenoMM,HuangL,GuH,LarsenRetal.:Asuiteoftransgenic driverandreportermouselineswithenhancedbrain-cell-type targetingandfunctionality.Cell2018,174:465-480e422.
19.
Millisecond-timescale,BoydenES,ZhangF,BamberggeneticallyE,NageltargetedG,DeisserothopticalcontrolK: of neuralactivity.NatNeurosci2005,8:1263-1268.
ThefirstuseofoptogenetictechnologyinvitroalongwithRef.[20]which
providedthefirstuseofoptogeneticsinvivotocontrolneuronalfiringwith
laserpulses.
20. LiX,GutierrezDV,HansonMG,HanJ,MarkMD,ChielH, HegemannP,LandmesserLT,HerlitzeS:Fastnoninvasive activationandinhibitionofneuralandnetworkactivityby vertebraterhodopsinandgreenalgaechannelrhodopsin.Proc NatlAcadSciUSA2005,102:17816-17821.
21. ArmbrusterB,RothB:Creationofdesignerbiogenicamine receptorsviadirectedmolecularevolution.
Neuropsychopharmacology2005,30:S265.
22.
ArmbrusterlocktofittheBN,keyLiX,toPauschcreateMH,afamilyHerlitzeofS,GRothprotein-coupledBL:Evolvingthe receptorspotentlyactivatedbyaninertligand.ProcNatlAcad SciUSA2007,104:5163-5168.
ThefirstpaperdescribingtheinventionandutilityofDREADDs.
23. NakaiJ,OhkuraM,ImotoK:Ahighsignal-to-noiseCa(2+)probe composedofasinglegreenfluorescentprotein.NatBiotechnol 2001,19:137-141.
24. SiegelMS,IsacoffEY:Ageneticallyencodedopticalprobeof membranevoltage.Neuron1997,19:735-741.
25. TianL,HiresSA,MaoT,HuberD,ChiappeME,ChalasaniSH, PetreanuL,AkerboomJ,McKinneySA,SchreiterERetal.:
Imagingneuralactivityinworms,fliesandmicewithimproved GCaMPcalciumindicators.NatMethods2009,6:875-881.
26. FlusbergBA,CockerED,PiyawattanamethaW,JungJC, CheungEL,SchnitzerMJ:Fiber-opticfluorescenceimaging. NatMethods2005,2:941-950.
27. FlusbergBA,JungJC,CockerED,AndersonEP,SchnitzerMJ:In vivobrainimagingusingaportable3.9gramtwo-photon fluorescencemicroendoscope.OptLett2005,30:2272-2274.
28. StraderCD,GaffneyT,SuggEE,CandeloreMR,KeysR, PatchettAA,DixonRA:Allele-specificactivationofgenetically engineeredreceptors.JBiolChem1991,266:5-8.
29. CowardP,WadaHG,FalkMS,ChanSD,MengF,AkilH, ConklinBR:Controllingsignalingwithaspecificallydesigned Gi-coupledreceptor.ProcNatlAcadSciUSA1998,95:352-357.
30. WestkaemperR,GlennonR,HydeE,ChoudharyM,KhanN, RothB:Engineeringinaregionofbulktoleranceintothe 5-HT2Areceptor.EurJMedChem1999,34:441-447.
31.
RedfernHennighausenCH,CowardL,BujardP,DegtyarevH,FishmanMY,GI,ConklinLeeEK,BR:KwaConditionalAT, expressionandsignalingofaspecificallydesigned Gi-coupledreceptorintransgenicmice.NatBiotechnol1999,
17:165-169. combinedwithdetailedstructuralstudies,toolswill
con-tinue to be developed that will allow a much more sophisticated understanding of intracellular, cellular, and especially neuronal signaling in vitro and in
vivo. The ability to multiplex these tools will allow a morenuanced viewofsuch signalingin thefuture,and thuswillallow ustoextendCrick’svision yetfurther.
Conflict
of
interest
statement
Nothingdeclared.Acknowledgements
Workintheauthor’slabissupportedbytheNIH(R37DA045657, RO1MH112205,U24DK116195)andtheMichaelHookerDistinguished ProfessorshipinPharmacology.TheauthorthanksWesKroeze,PhDfor helpfuledits.
References
and
recommended
reading
Papersofparticularinterest,publishedwithintheperiodofreview,
32. MuellerKL,HoonMA,ErlenbachI,ChandrashekarJ,ZukerCS, RybaNJ:Thereceptorsandcodinglogicforbittertaste.Nature 2005,434:225-229.
33. GuettierJM,GautamD,ScarselliM,RuizdeAzuaI,LiJH, RosemondE,MaX,GonzalezFJ,ArmbrusterBN,LuHetal.:A chemical-geneticapproachtostudyGproteinregulationof betacellfunctioninvivo.ProcNatlAcadSciUSA2009,
106:19197-19202.
34. FarrellMS,PeiY,WanY,YadavPN,DaigleTL,UrbanDJ,LeeHM, SciakyN,SimmonsA,NonnemanRJetal.:AGasDREADD mouseforselectivemodulationofcAMPproductionin striatopallidalneurons.Neuropsychopharmacology2013,
38:854-862.
35.
JainChronicS,RuizactivationdeAzuaofI,aLudesignerH,WhiteG(q)-coupledMF,GuettierJM,receptorWessJ: improvesbetacellfunction.JClinInvest2013,123:1750-1762.
AnimportantpaperwhichshowshowDREADDscanbeusedtoidentify
GPCRsignalingcascadeswhichcouldpotentiallybeusedtherapeutically
toimprovecellularfunction.
36. HuJ,SternM,GimenezLE,WankaL,ZhuL,RossiM,MeisterJ, InoueA,Beck-SickingerAG,GurevichVV,WessJ:AG protein-biaseddesignerGprotein-coupledreceptorusefulfor studyingthephysiologicalrelevanceofGq/11-dependent signalingpathways.JBiolChem2016,291:7809-7820.
37.
VardySassanoE,FM,RobinsonHuangJE,XP,LiZhuC,OlsenH,UrbanRH,DJDiBertoetal.:JF,AnewGiguereDREADDPM, facilitatesthemultiplexedchemogeneticinterrogationof behavior.Neuron2015,86:936-946.
An important paper which shows how structure-based design of
DREADDscanleadtoenhancedcontrolandmultiplexing.
38. HudsonBD,ChristiansenE,TikhonovaIG,GrundmannM, KostenisE,AdamsDR,UlvenT,MilliganG:Chemically engineeringligandselectivityatthefreefattyacidreceptor 2basedonpharmacologicalvariationbetweenspecies orthologs.FASEBJ2012,26:4951-4965.
39. WackerD,WangC,KatritchV,HanGW,HuangXP,VardyE, McCorvyJD,JiangY,ChuM,SiuFYetal.:Structuralfeaturesfor functionalselectivityatserotoninreceptors.Science2013,
340:615-619.
40. CheT,MajumdarS,ZaidiSA,OndachiP,McCorvyJD,WangS, MosierPD,UpretyR,VardyE,KrummBEetal.:Structureofthe nanobody-stabilizedactivestateofthekappaopioidreceptor. Cell2018,172:55-67e15.
41.
Nakajimaanovel,arrestin-biasedKI,WessJ:DesigndesignerandfunctionalGprotein-coupledcharacterizationreceptorof. MolPharmacol2012,82:575-582.
Thisshowshowcarefullydesignedmutantscanbecreatedtoalterthe
signalingpropertiesofDREADDs.
42. KrashesMJ,KodaS,YeC,RoganSC,AdamsAC,CusherDS, Maratos-FlierE,RothBL,LowellBB:Rapid,reversibleactivation ofAgRPneuronsdrivesfeedingbehaviorinmice.JClinInvest 2013,121:1424-1428.
43. ZhuH,AryalDK,OlsenRH,UrbanDJ,SwearingenA,ForbesS, RothBL,HochgeschwenderU:Cre-dependentDREADD (DesignerReceptorsExclusivelyActivatedbyDesignerDrugs) mice.Genesis2016,54:439-446.
44. RothBL:DREADDsfor neuroscientists.Neuron2016,89:683-694.
45. ChenX,ChooH,HuangXP,YangX,StoneO,RothBL,JinJ:The firststructure-activityrelationshipstudiesfordesigner receptorsexclusivelyactivatedbydesignerdrugs.ACSChem Neurosci2015,6:476-484.
46. DongS,RoganSC,RothBL:Directedmolecularevolutionof DREADDs:agenericapproachtocreatingnext-generation RASSLs.NatProtoc2010,5:561-573.
47. StachniakTJ,GhoshA,SternsonSM:Chemogeneticsynaptic silencingofneuralcircuitslocalizesa
hypothalamus!midbrainpathwayforfeedingbehavior. Neuron2014,82:797-808.
controlofneuronalactivityintransgenicmiceexpressing evolvedGprotein-coupledreceptors.Neuron2009,63:27-39.
49. KrashesM,KodaS,ChiangPingY,RoganSC,AdamsA, Maratos-FlierE,RothBL,LowellB:Rapid,reversibleactivation ofAgRPneuronsdrivesfeedingbehavior.JClinInvest2011,
121:1424-1428.
50. BenderD,HolschbachM,StocklinG:Synthesisofn.c.a. carbon-11labelledclozapineanditsmajormetabolite clozapine-N-oxideandcomparisonoftheirbiodistributioninmice.Nucl MedBiol1994,21:921-925.
51. GomezJL,BonaventuraJ,LesniakW,MathewsWB,Sysa-ShahP, RodriguezLA,EllisRJ,RichieCT,HarveyBK,DannalsRFetal.:
Chemogeneticsrevealed:DREADDoccupancyandactivation viaconvertedclozapine.Science2017,357:503-507.
52. JannMW,LamYW,ChangWH:Rapidformationofclozapinein guinea-pigsandmanfollowingclozapine-N-oxide
administration.ArchIntPharmacodynTher1994,328:243-250.
53. RothBL,ShefflerDJ,KroezeWK:Magicshotgunsversusmagic bullets:selectivelynon-selectivedrugsformooddisorders andschizophrenia.NatRevDrugDiscov2004,3:353-359.
54.
SlocumThompsonS,JinKJ,J,KhajehaliLiuJ,XiongE,BradleyY,OlsenSJ,RHNavarreteetal.:DREADDJS,HuangagonistX-P, 21Isaneffectiveagonistformuscarinic-basedDREADDsin vitroandinvivo.ACSPharmacolTranslSci2018,1:61-72.
ThefirstusefulnonCNO-basedDREADDligandforinvivostudies.
55. BallesterosJA,WeinsteinH:Integratedmethodsforthe constructionofthree-dimensionalmodelsandcomputational probingofstructure-functionrelationsinGprotein-coupled receptors.MethodsNeurosci1995,25:366.
56. HagaK,KruseAC,AsadaH,Yurugi-KobayashiT,ShiroishiM, ZhangC,WeisWI,OkadaT,KobilkaBK,HagaT,KobayashiT:
StructureofthehumanM2muscarinicacetylcholinereceptor boundtoanantagonist.Nature2012,482:547-551.
57. KruseAC,RingAM,ManglikA,HuJ,HuK,EitelK,HubnerH, PardonE,ValantC,SextonPMetal.:Activationandallosteric modulationofamuscarinicacetylcholinereceptor.Nature 2013,504:101-106.
58. McCorvyJD,WackerD,WangS,AgegnehuB,LiuJ,LansuK, TriboAR,OlsenRHJ,CheT,JinJ,RothBL:Structural determinantsof5-HT2Breceptoractivationandbiased agonism.NatStructMolBiol2018,25:787-796.
59. HauserAS,AttwoodMM,Rask-AndersenM,SchiothHB, GloriamDE:TrendsinGPCRdrugdiscovery:newagents, targetsandindications.NatRevDrugDiscov2017,16:829-842.
60. WuH,WackerD,MileniM,KatritchV,HanGW,VardyE,LiuW, ThompsonAA,HuangXP,CarrollFIetal.:Structureofthehuman kappa-opioidreceptorincomplexwithJDTic.Nature2012,
485:327-332.
61. O’ConnorC,WhiteKL,DoncescuN,DidenkoT,RothBL, CzaplickiG,StevensRC,WuthrichK,MilonA:NMRstructureand dynamicsoftheagonistdynorphinpeptideboundtothe humankappaopioidreceptor.ProcNatlAcadSciUSA2015,
112:11852-11857.
62. RothBL,BanerK,WestkaemperR,SiebertD,RiceKC, SteinbergS,ErnsbergerP,RothmanRB:SalvinorinA:apotent naturallyoccurringnonnitrogenouskappaopioidselective agonist.ProcNatlAcadSciUSA2002,99:11934-11939.
63. VardyE,MosierPD,FrankowskiKJ,WuH,KatritchV, WestkaemperRB,AubeJ,StevensRC,RothBL: Chemotype-selectivemodesofactionofkappa-opioidreceptoragonists.J BiolChem2013,288:34470-34483.
64. DenisRG,Joly-AmadoA,WebberE,LangletF,SchaefferM, PadillaSL,CansellC,DehouckB,CastelJ,DelbesASetal.:
PalatabilitycandrivefeedingindependentofAgRPneurons. CellMetab2015,22:646-657.
amygdalacontrolslearninginthelateralamygdala.Nat Neurosci2017,20:1680-1685.
66. StarkweatherCK,GershmanSJ,UchidaN:Themedialprefrontal cortexshapesdopaminerewardpredictionerrorsunderstate uncertainty.Neuron2018,98:616-629e616.
67. KoehlA,HuH,MaedaS,ZhangY,QuQ,PaggiJM,LatorracaNR, HilgerD,DawsonR,MatileHetal.:Structureofthemicro-opioid receptor-Giproteincomplex.Nature2018,558:547-552.
68. SternsonSM,RothBL:Chemogenetictoolstointerrogatebrain functions.AnnuRevNeurosci2014,37:387-407.
69. YizharO,FennoLE,DavidsonTJ,MogriM,DeisserothK:
Optogeneticsinneuralsystems.Neuron2011,71:9-34.
70.
KatoInoueHE,K,ItoKimS,YS,RamakrishnanPaggiJM,EvansC,FennoKE,LEAllenetWE,al.:StructuralRichardsonC, mechanismsofselectivityandgatinginanion
channelrhodopsins.Nature2018,561:349-354.
Animportantpaperwhichprovidesaroadmapforthedesignofnew
opsinsbasedonstructuralconsiderations.
71.
VenkatakrishnanWangS,WackerAJ,D,LevitHuangA,XP,CheDrorT,BetzRO,RM,ShoichetMcCorvyBK,JD,RothBL:
D4dopaminereceptorhigh-resolutionstructuresenablethe discoveryofselectiveagonists.Science2017,358:381-386.
Afoundationalpaperwhich describeshowstructure-baseddrug
dis-covery can be used to discover novel chemotypes for dopamine
receptors.
72. IsbergV,MordalskiS,MunkC,RatajK,HarpsoeK,HauserAS, VrolingB,BojarskiAJ,VriendG,GloriamDE:GPCRdb:an informationsystemforGprotein-coupledreceptors.Nucleic AcidsRes2017,45:2936.
![Figure 2 also shows an active state structure of the wild-type M2 muscarinic receptor (2D [57]), a model of an active state hM2Di-DREADD (Figure 2b) and a summary of key transitions (Figure 2c)](https://thumb-us.123doks.com/thumbv2/123dok_us/8205849.2175825/3.918.91.838.748.1040/figure-structure-muscarinic-receptor-figure-summary-transitions-figure.webp)
