CARRIAGE OF ANTIBIOTIC-RESISTANT STAPHYLOCOCCUS AUREUS BY LIVESTOCK WORKERS AND HOUSEHOLD MEMBERS IN NORTH CAROLINA
Maya Nadimpalli
A thesis submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Masters of Science in the Department of
Environmental Sciences & Engineering.
Chapel Hill 2012
Approved by: Dr. Jill Stewart
ii
Abstract
MAYA NADIMPALLI: Carriage of Antibiotic-Resistant Staphylococcus aureus by Livestock Workers and Household Members in North Carolina
(Under the direction of Dr. Jill Stewart)
iii
Acknowledgements
This thesis would not have been possible without the help of several individuals. First, I would like to thank my family and friends for their advice and constant support. I would like to thank my advisor, Dr. Jill Stewart, for patiently helping me learn microbiology from scratch, and members of my committee, Dr. Christopher Heaney and Dr. Rebecca Fry for their advice as I worked on this project. Dr. Heaney in particular has acted as a second advisor to me here at UNC.
I would like to thank current and former members of the Stewart lab group for helping me throughout this project, particularly Jennifer Shields, Kevin Myers, and Sarah Hatcher. I am especially grateful to members of the Department of Epidemiology for their support and
guidance. I would particularly like to thank Jessica Rinsky, Dr. Steven Wing, and Alan Kinlaw. I am also grateful to several individuals at the Staphylococcal Laboratory at the Statens Serum Institut for welcoming me into their lab for a few days this past November. I learned much more there than I ever could have on my own.
iv
v
Table of Contents
List of Tables ...vii
List of Figures ... viii
List of Common Abbreviations ... ix
C
HAPTER1:
I
NTRODUCTION... 1
OBJECTIVES ... 3
B
ACKGROUND... 4
INDUSTRIAL ANIMAL PRODUCTION IN THE UNITED STATES ... 4
INDUSTRIAL ANIMAL PRODUCTION:PUBLIC HEALTH CONCERNS ... 5
Environmental and health concerns ... 5
Propagation of antimicrobial resistance ... 7
FOOD ANIMAL PRODUCTION IN NORTH CAROLINA ... 8
STAPHYLOCOCCUS AUREUS ... 9
Epidemiology of S. aureus carriage ... 10
History of antibiotic resistance in S. aureus ... 12
Mechanisms for antibiotic resistance ... 14
Molecular typing of S. aureus ... 15
METHICILLIN-RESISTANT STAPHYLOCOCCUS AUREUS ... 19
History of MRSA ... 21
LIVESTOCK-ASSOCIATED ST398 ... 24
Origins of MRSA ST398 ... 24
vi
ST398 Infection ... 25
Colonization versus Contamination ... 26
ST398 in North America ... 26
Unanswered Questions ... 29
MULTIDRUG-RESISTANT S. AUREUS ... 30
MDRSA Associated with CAFOs ... 30
RESEARCH RATIONALE ... 31
C
HAPTER2:
M
ETHODS... 34
STUDY DESIGN AND TIMEFRAME ... 34
ADMINISTRATION OF QUESTIONNAIRES AND NASAL SWAB SAMPLE COLLECTION ... 34
ISOLATION OF MSSA AND MRSA FROM NASAL SWABS ... 35
MOLECULAR CONFIRMATION OF MSSA AND MRSA ... 37
ANTIBIOTIC SUSCEPTIBILITY TESTING ... 38
CONFIRMATION OF CC398 AND ADDITIONAL GENE TARGETS ... 39
C
HAPTER3:
R
ESULTS... 41
PREVALENCE OF S. AUREUS,MDRSA, AND MRSA ... 41
RESISTANCE PHENOTYPES ... 43
PREVALENCE AND DISTRIBUTION OF CC398 ... 44
Resistance phenotypes of CC398 isolates ... 46
SENSITIVITY AND SPECIFICITY COMPARISONS ... 47
C
HAPTER4:
D
ISCUSSION... 49
LIMITATIONS AND FUTURE RESEARCH ... 51
vii
List of Tables
Table 1: Shift in NC hog production from 1982-2007 ... 9
Table 2: Growth in NC organic operations from 1997-2007 ... 9
Table 3: Primer sequences used for multiplex PCR ... 37
Table 4: Antibiotics used in susceptibility testing via disk diffusion ... 38
Table 5: Summary of study participants ... 41
viii
List of Figures
Figure 1: Proliferation and fate of antibiotic resistant bacteria
during CAFO waste storage and disposal ... 6
Figure 2: Skeletal representation of the SCCmec element ... 14 Figure 3: Protocol for isolation and characterization of MSSA
and MRSA from nasal swabs ... 36 Figure 4: Carriage of S. aureus, multidrug-resistant S. aureus (MDRSA),
and methicillin-resistant S. aureus (MRSA) in each study group ... 42 Figure 5: Antibiotic resistance profiles of 8 MRSA isolates collected during study ... 42
Figure 6: Percentage of S. aureus-positive ABF, IND, and PP participants
demonstrating full resistance to 9 antibiotic classes ... 44
Figure 7: Prevalence of CC398 among S. aureus-positive IND, ABF,
and PP participants ... 45
Figure 8: Distribution of CC398 among S. aureus-positive IND, ABF,
and PP participants ... 46
Figure 9: Antibiotic resistance profiles of 20 CC398 isolates collected
ix
List of Common Abbreviations
ABF Antibiotic-free
BP Baird-Parker with Egg Yolk Tellurite Enrichment
CA Community-associated
CAFO Concentrated animal feeding operation
CC Clonal complex
CoNS Coagulase-negative staphylococci
CS CHROMagar™ Staph aureus
HA Hospital-associated
IND Industrial
LA Livestock-associated
MDRSA Multidrug-resistant Staphylococcus aureus
MGE Mobile genetic element
MLST Multilocus sequence typing
MRSA Methicillin-resistant Staphylococcus aureus
MSSA Methicillin-susceptible Staphylococcus aureus
PBP Penicillin binding protein
x
PP Processing plant
pvl Panton-Valentine leukocidin
REACH Rural Empowerment for Community Help
SCCmec Staphylococcal cassette chromosome mec
ST Sequence type
spa Surface protein A
CHAPTER 1: INTRODUCTION
Staphylococcus aureus is a ubiquitous human commensal carried by approximately one third of the American population, primarily in the nostrils (Gorwitz et al., 2008). S. aureus is also an adept human pathogen capable of causing a wide range of infections, including skin and soft tissue infections, bacteremia, endocarditis, gastrointestinal illness, necrotizing pneumonia, post-operative infections, and toxic shock syndrome (Loir et al., 2003; Lowy, 1998). Antibiotics have been used to treat S. aureus infections for decades, but rapidly disseminating multidrug-resistant strains of S. aureus (MDRSA) have made treatment more protracted, more burdensome, and less successful in recent years (Chambers et al., 2009). Methicillin-resistant S. aureus (MRSA) has proven particularly difficult to treat.
2
of community-acquired MRSA infections are now rising faster than hospital-acquired MRSA infections in the United States. Community-acquired MRSA is currently considered epidemic in the United States (Kennedy et al., 2009).
Most recently, the evolution of a community-acquired MRSA strain associated with intensive livestock production, primarily swine production, has been discovered. Although this novel strain of MRSA is believed to have originated from human-associated methicillin-susceptible S. aureus (MSSA) strains (Price et al., 2012), it now persists in a livestock reservoir and can be transmitted between humans and animals. Multilocus sequence type 398 (ST398) was first detected in association with hog-farming in the Netherlands in 2004, and since then has been identified in several European countries as well as South America, China, Canada, and as of 2009, the United States (Arriola et al., 2011; Khanna et al., 2008; Smith et al., 2009; Wagenaar et al., 2009). Livestock-associated MRSA is transmitted not only to farm workers, with observed carriage rates between 23% and 49%, but their families as well (Huijsdens et al., 2006; R. Köck et al., 2009). MRSA and MSSA ST398 infections have also been documented in the greater community (Jiménez et al., 2011), including in individuals with no clear connection to livestock production (Bhat et al., 2009; Golding et al., 2010; Mediavilla et al., 2012; Skov, 2011; Uhlemann et al., 2012).
3
North Carolina, however, is also home to a burgeoning sustainable farming movement (Greene, 2010). These farms also raise pork and poultry, along with other animals and produce, without the use of antibiotics, and with limited reliance on external inputs. Given that the persistent, subtherapeutic dosing of animals with antibiotics could create a selective pressure for antibiotic-resistant bacteria which could then be transmitted to humans (Khachatourians, 1998; Pew Charitable Trusts, 2008), a comparison of MDRSA and MRSA carriage rates among industrial workers and their household members to carriage rates among antibiotic-free farmers and their household members merits investigation.
Objectives
The goals of this research were four-fold:
1) To compare the prevalence of carriage of MDRSA and MRSA in 100 consenting industrial livestock workers and household members to 100 antibiotic-free livestock workers and household members in North Carolina;
2) To compare antibiotic resistance patterns among S. aureus-positive individuals in both groups;
3) To use lineage-specific PCR to determine whether CC398 is present in North Carolina; and 4) To compare the sensitivity and specificity of two differential media, Baird Parker and BBL™
CHROMagar™ Staph aureus, in their capacity to detect S. aureus from nasal swabs.
4
B
ACKGROUNDIndustrial Animal Production in the United States
5
Industrial Animal Production: Public Health Concerns
Environmental and health concerns
Animal feeding operations in the United States produce an estimated 500 million tons of waste per year, three times more than the human population (United States Environmental Protection Agency, 2003). Roughly 47% to 60% of this waste is generated by CAFOs (Greger et al., 2010). Waste is collected and stored in large pits beneath caged animals or flushed into outdoor, open-air lagoons, which may serve as optimal breeding grounds for the bacteria that propagate antimicrobial resistance. In North Carolina, no further treatment is required of CAFO liquid manure before it is eventually sprayed onto fields. Over application of manure can lead to runoff laden with harmful pollutants, including nutrients, pesticides, heavy metals, veterinary pharmaceuticals, resistance genes, and infectious pathogens (Figure 1); these pollutants can also leach through permeable soil into aquifers. Lagoons themselves often leak, overflow, or rupture, which can also contaminate groundwater and surrounding surface waters (Sapkota et al., 2007; Steve Wing et al., 2002). Exposure to waterborne contaminants can be particularly serious for vulnerable populations, including children, pregnant women, and the elderly (Reynolds et al., 2008).
6
dust from animal dander and waste pose further health risks. Acute occupational exposure to any combination of these irritants can result in shortness of breath, coughing, and inflammation of the lungs and mucous membranes; chronic exposures have been linked to bronchitis, asthma, and wheezing (Cole, et al., 2000). These health effects are also observed in the communities that surround CAFOs, due to both direct emissions and aerosolization of particulates during spraying. In combination with degraded mental and social health, lower quality of life measurements are often reported in such communities (S. Wing et al., 2000).
7
populations are often most susceptible to the environmental and public health harms produced by CAFO emissions and the land application of swine waste, due to lack of access to medical care, poor nutrition, and other risk factors. Furthermore, these groups often have the least political power to impact policies and practices related to industrial livestock operations in their communities.
Propagation of antimicrobial resistance
8
In response to these concerns, leaders in Congress have called for an end to the use of clinically-relevant antibiotics in food animal production in the “Preservation of Antibiotics for Medical Treatment Act” (H.R. 965/S. 1211; PAMTA). In 2012, the FDA announced plans to place controls on the use of cephalosporins in animal production. More recently, the FDA was also court mandated to withdraw approvals for most non-therapeutic uses of penicillins and tetracyclines in food animals. However, PAMTA is still being debated, and any further regulation by the FDA may take years. Unfortunately, legislative and regulatory action could be outpaced by the evolution and dissemination of novel antimicrobial mechanisms.
Food animal production in North Carolina
Industrial animal production has grown rapidly in North Carolina in the past few decades, with the state moving from fifteenth to second in hog production nationally (Table 1), and from fourth to third in poultry production. North Carolina’s hog CAFOs are concentrated in the southern Piedmont and in the eastern coastal plain of the state, with the highest densities occurring in Duplin and Sampson counties. Together, hog production in these two counties accounts for nearly half of the state’s total production. Growth has leveled off in the last several years due to a moratorium on permitting new hog CAFO liquid waste storage lagoons (North Carolina Department of Agriculture and Consumer Services, 2010). Meanwhile, poultry production in North Carolina has increased in recent years. Turkey production is highest in Duplin and Sampson counties, but egg production occurs in the central part of the state and broiler production is increasing throughout the state.
9
or 1.7% of total US sales (United States Department of Agriculture, 2010). North Carolina is the largest retailer of organic agricultural products in the Southeastern US.
Table 1. Shift in NC hog production from 1982-2007
Year Number of Operations Number of hogs
1982 11,390 2.0 million
2007 2,836 10.1 million
Source: (Chee-Sanford, et al., 2009)
Table 2. Growth in NC organic operations from 1997-2007
Year # of operations # of cows, pigs, and sheep # of poultry
1997 4 0 29,700
2007 112 156 1,088,860
Source: (Greene, 2010)
Staphylococcus aureus
Staphylococci are ubiquitous, gram-positive cocci which tend to cluster in grape-like bunches or in short chains. Most staphylococci are 0.5 to 1 µm in diameter, nonmotile, halotolerant to concentrations of 10% w/v NaCl, grow at temperatures between 18° and 40°C, and are aerobic or facultatively anaerobic (Murray et al., 2002). Typically, staphylococci are found in association with the skin, skin glands, and mucous membranes of warm-blooded animals (Crossley et al., 1997). Staphylococci are one of the most common bacteria to cause disease in humans, and in the United States, staphylococci are the most common cause of nosocomial infection (Crossley, et al., 1997). Of at least 32 species, Staphylococcus aureus is the most pathogenic and consequently the best researched (Crossley, et al., 1997).
10
(Murray, et al., 2002). S. aureus can be distinguished from other species of staphylococci by its ability to produce coagulase, a protein which binds fibrinogen in the blood and converts it to insoluble fibrin, thus localizing an infection and protecting the bacteria from immune response cells. S. aureus also uniquely produces surface protein A, a cell-wall bound protein which is involved in increased pathogencity of the organism. S. aureus’ fitness is further enhanced by its production of staphyloxanthin, a carotenoid pigment that acts as a protective antioxidant while imparting a characteristic golden color to S. aureus colonies. Several strains of S. aureus are capable of producing a variety of exotoxins, including TSST-1 (associated with toxic shock syndrome), exfoliative toxins (associated with staphylococcal scaled skin syndrome), staphylococcal enterotoxin B (associated with food poisoning), and α toxin, β toxin, and Panton-Valentine leukocidin (all associated with necrotizing pneumonia) (Lowy, 1998). The production of these proteins and toxins allows S. aureus to cause a wide diversity of benign and lethal infections.
Though humans and animals are its primary reservoir, S. aureus has also been detected in air (Bassetti et al., 2005), in drinking and waste waters (LeChevallier et al., 1980; Rosenberg-Goldstein, 2010), and in food (Loir, et al., 2003). It has been demonstrated that S. aureus can persist in river and sea water for up to two weeks (Tolba et al., 2008), as well as on dry surfaces for days to months (Kramer et al., 2006). S. aureus has also been detected in hog and poultry waste (Dimitracopoulos et al., 1977; Graham et al., 2009).
Epidemiology of S. aureus carriage
11
site for S. aureus colonization. An additional 20% of the population carry it on their skin, hair, or in their throats (Bhatia et al., 2007). Nasal carriage of S. aureus is not evenly distributed among the US population. Rather, colonization with S. aureus has been found to be significantly more common among males, compared with females (p<.001), among non-Hispanic whites and Mexican Americans, compared with non-Hispanic blacks (p<.001 and p<.01, respectively), and among persons aged <20 years, compared with older persons (p<.001) (Gorwitz, et al., 2008). Rates of staphylococcal colonization are higher than average among intravenous drug users, individuals with type 1 diabetes, and AIDS patients (Lowy, 1998). Significant associations between obesity and S. aureus carriage have also been described (Gorwitz, et al., 2008). Importantly, carriage of S. aureus can be transient. Longitudinal studies have described three patterns of S. aureus carriage in the population: persistent carriers, who are thought to comprise 20% of the population, persistent non-carriers, who are thought to comprise an additional 20% of the population, and intermittent carriers, who are thought to comprise the remaining 60% of the population (Peacock et al., 2001). Though concrete population determinants of carriage remain speculative, race, genetics, immune characteristics, strain diversity, and environment may play a role. While nasal carriage may be a risk factor for infection, most individuals who are colonized never become infected (Gorwitz, et al., 2008). Eradication of nasal colonization via mupirocin treatment may or may not prevent subsequent S. aureus infection (Wertheim et al., 2005).
12
imprisonment (Aiello et al., 2006), use of work-out gyms (Kirkland et al., 2008), and pig farming (Voss et al., 2005). While hospital-acquired MRSA strains were once predominant in the US, they are largely being replaced by community-acquired strains. Longitudinal studies have demonstrated that increasing rates of MRSA infection in the US in the early 2000s were linked to an expanding reservoir of such community-associated strains (Carleton et al., 2004).
Historyof antibiotic resistance in S. aureus
In the 1940s, penicillin became the first commercially available antibiotic, and its discovery heralded a new age of pathogen control. Almost simultaneously, however, there came reports of certain staphylococci that could destroy the antibiotic through production of an enzyme then termed “penicillinase.” Penicillin acts by binding to and inhibiting penicillin binding proteins (PBP), which are integral in catalyzing the peptidoglycan cross-links in bacterial cell walls. As bacteria continue to naturally degrade peptidoglycan links without the capacity to construct new ones, their cell structure weakens, resulting in lysis. Penicillinase, encoded by the blaZ gene, hydrolyzes the β-lactam ring of penicillin before it can act, thus protecting bacteria from the antibiotic’s mode of action (Lowy, 1998). As early as 1946, it was estimated that 60% of hospital S. aureus isolates in the UK were resistant to penicillin; current estimates top 90% (Crossley, et al., 1997). Penicillin-resistance is now nearly ubiquitous in S. aureus (Crossley, et al., 1997).
13
to all β-lactam antibiotics, including carbanepems and cephalosporins. The gene responsible for this resistance, mecA, encodes for an altered penicillin-binding protein (PBPa), which has a lower affinity for all β-lactam antibiotics. When bacteria produce PBPa, cell wall synthesis is able to proceed unaffected in the presence of β-lactams. At least 4 distinct waves of mecA-encoded methicillin-resistance have been observed, resulting in the global MRSA pandemic occurring today.
Vancomycin-intermediate and vancomycin-resistant S. aureus (VISA and VRSA) have emerged in the last decade as a result of the rising use of vancomycin to treat MRSA infections. However, VISA and VRSA infections remain few in number and have been isolated exclusively in the healthcare setting (Chambers, et al., 2009). VRSA carry the vanA gene cassette, presumably obtained from vancomycin-resistant Enterococcus (Chang et al., 2003). Carriage of this cassette appears to present a significant burden on the bacteria, which may account for the limited spread of VRSA in hospitals and its absence from the community (Foucault et al., 2009). The mechanisms for low-level and intermediate vancomycin resistance remain unclear, though certain morphological changes in VISA, such as cell wall thickening and intensive pigmentation, have been observed (Renzoni et al., 2010).
14 Mechanisms for antibiotic resistance
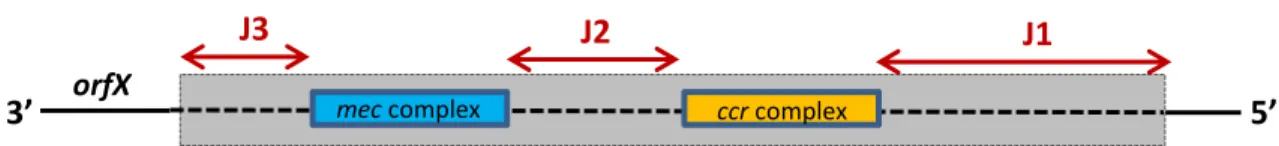
One of the most important mobile genetic elements (MGEs) encoding resistance in S. aureus is the Staphylococcal Cassette Chromosome mec (SCCmec; see Figure 2). SCC elements are genomic islands that are ubiquitous among staphylococci; SCCmec contains the mecA gene and is most commonly associated with MRSA (although partial, nonfunctional SCCmec sequences – known as “ghost sequences” - have been detected in MSSA) (Corkill et al., 2004). SCCmec elements are 20 - 66 kb in length and are integrated at the 3’ end of the orfX gene, whose function remains unknown. These elements carry two major complexes: the mec complex, encoding the mecA gene and two regulatory genes (mecI and mecRI), and the ccr complex, which encodes for one or two site-specific cassette chromosome recombinases, allowing the SCCmec element to be excised and inserted into new genomes (Deurenberg et al., 2007). Between these complexes and the 3’ and 5’ ends of the SCCmec lie three junkyard, or joining regions: J3, located between orfX and mec, J2, located between mec and ccr, and J1, located between ccr and the chromosomal region flanking the SCCmec. Several allotypes of the mec and ccr complexes have been identified, and SCCmec elements can be typed based on these differences. SCCmec types I-VIII are the most well-known, though several other types likely exist. Subtyping of SCCmec is also possible by classifying structural differences in the J1, J2, and J3 regions. In addition to the mecA gene, certain SCCmec complexes (namely II and III) encode additional drug resistance genes on integrated mobile elements, such as plasmids and transposons (Skov, 2011).
Figure 2. Skeletal representation of the SCCmec element. orfX
3’
5’
J3
J2
J1
15
S. aureus can carry resistance genes inserted at other sites on the chromosome besides SCCmec, as well as on plasmids (Deurenberg, et al., 2007). Additionally, S. aureus can develop resistance through random mutation of existing genes. Such mutations require selective pressure to maintain themselves in subsequent generations.
Molecular typing of S. aureus
Molecular typing originally evolved as an epidemiological tool to track the dissemination of MRSA strains into regional and global populations. However, any strain of S. aureus can be typed by any of these same methods. The typing methods that will be discussed here are: pulsed-field gel electrophoresis (PFGE), multilocus sequence typing (MLST), typing of the variable tandem repeat region of surface protein A (spa typing), and whole genome sequence typing (WGST).
Pulsed-Field Gel Electrophoresis
16
are difficult if standardized protocols are not used. Thus, standardized PFGE databases have only been achieved at the national level (Deurenberg, et al., 2007). Additional drawbacks of PFGE typing include high cost and time requirements, and the need for trained lab personnel. ST398 - the strain associated with intensive livestock production, is also “nontypeable” by PFGE using Sma1 (Voss, et al., 2005).
Multilocus Sequence Typing
MLST is a relatively new typing method that also demonstrates excellent discriminatory power (Maiden et al., 1998). MLST involves sequence analysis of 0.5-kb internal fragments of seven S. aureus housekeeping genes: arcC, aroE, glpF, gmk, pta, tpi, and yquiL. Differences in these sequences correspond to distinct alleles for each housekeeping gene. Each allele is assigned a unique number; any S. aureus isolate can be identified by its allele profile for these seven genes, also known as its “sequence type.” A major advantage of MLST over PFGE is that sequence data are truly commutable across laboratories and across countries. Currently, there exists one global database containing all known sequence types (http://mlst.net), to which any researcher can upload new alleles. Global epidemiological investigations of MRSA and MSSA infection are thus possible using MLST. Unfortunately, like PFGE, MLST is also time-consuming and expensive.
17
and may diversify independently to produce their own set of single-locus and double-locus variants (Deurenberg, et al., 2007). A constantly updated diagram depicting all known clonal complexes and their relationships to one another is available at http://eburst.mlst.net.
Spa typing
Spa typing is even more discriminatory than MLST, and is much less costly since it involves sequencing of only a single locus. The gene encoding surface protein A is unique to S. aureus and contains a polymorphic region X, which is characterized by one to twenty-five 24-bp tandem repeats. Diversity in repeats is attributed to duplications of repeats, deletions, and less commonly, mutations (Deurenberg, et al., 2007). However, the first repeat in region X always begins with the sequence GAG, while subsequent repeats always begin with AAA (M. Sørum, personal communication, November 26, 2011). Sequencing the spa gene reveals how many tandem repeats are present in a strain, which repeats they are, and in what order they occur; this corresponds to a certain spa type. Researchers can identify spa types as well as upload novel spa types using a central spa server, available at http://www.spaserver.ridom.de. Because of its simplicity, low cost, and the fact that it can be performed using “in-house” technologies, this typing method is used widely in hospitals and research labs (Deurenberg, et al., 2007; Sørum, 2011).
18
including its spa gene. As a result, spa-CC012 has a heterogeneous clonal structure that includes both CC8 and CC30, and its ancestry cannot be inferred. To be absolutely sure of a strain’s sequence type and clonal complex, MLST must be performed in addition to spa typing (M. Sørum, personal communication, November 26, 2011).
Additionally, there exist two competing nomenclature systems for spa types which are used worldwide (Harmsen et al., 2003; Koreen et al., 2004). This renders comparison of published spa typing results considerably more difficult than comparison of MLST typing results.
Whole genome sequence typing
Whole genome sequence typing (WGST) represents the newest, best available method for tracking S. aureus subtypes at both the global and regional scale. Though PFGE is considered the gold standard for typing S. aureus isolates and for epidemiological investigations, it is only able to discriminate interregional isolates – i.e., it lacks the power needed to discriminate subtle intraregional variability (Harris et al., 2010). Such discriminatory power is needed to understand the microevolutionary events that lead to clonal differentiation. Unlike full-genome sequencing, which is economically unfeasible and too time-consuming for large population samples, WGST instead maps single nucleotide-polymorphisms (SNPs) in given strains to a reference genome. Next-generation sequencing technologies allow multiple bacterial isolates at be mapped at once, thus making it feasible to quickly generate whole-genome sequence data for a large number of bacteria (Harris, et al., 2010). From this sequence data, the SCCmec type, sequence type, and spa type for any given bacterial isolate can be determined. Additionally, resistance genes, virulence genes, and other MGEs that may be incorporated in the accessory genome are revealed.
19
of sequenced bacteria. S. aureus isolates from specific geographical regions tend to cluster on such trees; clades that are basally located elucidate where an S. aureus strain may have originated (Harris, et al., 2010). Phylogenetic trees can also explain whether or not a certain strain was introduced to a country or if it evolved independently. If it was introduced, its origin can typically be determined. Phylogenetic trees constructed from WGST data were used to determine the source of the recent cholera outbreak in Haiti (Hendriksen et al., 2011), as well as to corroborate the assumed source of MRSA ST398 in two adopted Chinese children in Denmark (Stegger et al., 2010). WGST data potentially has sufficient discriminatory power to reveal fine-scale transmission events between or within single hospitals (Harris, et al., 2010). PFGE and spa typing findings do not always correspond with evolutionary relationships delineated by WGST (Harris, et al., 2010), suggesting WGST may be a superior tool for epidemiological investigations. WGST is currently a more expensive S. aureus typing method, although prices for next-generation sequencing technology are declining rapidly. Bioinformatics expertise is also required to interpret WGST data and to construct phylogenetic trees. Therefore, WGST may not yet be suitable for routine use outside of highly specialized research laboratories.
Methicillin-resistant
Staphylococcus aureus
20
may be decreasing; from 2005-2008, there was approximately a 28% decrease in hospital-onset MRSA infections and a 17% decrease in community-onset infections (Kallen et al., 2010). It is estimated that MRSA infections cost the healthcare system up to an additional $9.7 billion annually (Klein, et al., 2007). Many MRSA infections tend to be multiply drug-resistant (Bordon, et al., 2010), further compounding the costs of treatment.
As described previously, methicillin resistance is conferred by acquisition of the mecA gene, which is carried on the SCCmec element. The source of SCCmec is unknown, although coagulase-negative staphylococci (CoNS) are thought to have first transferred this MGE to S. aureus (Skov, 2011). Evidence suggests that MRSA clones do not have a single ancestor; instead, SCCmec was introduced multiple times into different S. aureus lineages, resulting in the clonal diversity observed today (Deurenberg, et al., 2007). It should be noted that carriage of the SCCmec element is not inherently beneficial to S. aureus, as it is associated with a weaker cell wall (Skov, 2011). Therefore, antibiotic pressure is required to facilitate dissemination of SCCmec into MSSA.
In 2011, a novel mecA gene, homologue mecALGA251 and herein termed “LGA251”, was
21
2008-2010 was statistically significant (Holmes, 2011). The LGA251 gene homologue has only been detected in “animal-associated strains” thus far, and has not been detected in any S. aureus isolate from the United States (Holmes, 2011). Efforts are now underway to allow for the simultaneous detection of mecA and its homologue using clinical diagnostic tools (M. Stegger et al., 2011). The discovery of LGA251 highlights the importance of scrutinizing S. aureus isolates that demonstrate methicillin-resistance but are PCR-negative for the mecA gene. Other homologues of the mecA gene, besides LGA251, may in fact exist.
History of MRSA
Wave 1: 1960s – mid 1970s
The first wave of MRSA to hit Europe was essentially monoclonal, with all archived isolates belonging to a single subgroup of CC8 (Enright et al., 2002). Isolates from this subgroup carried the first characterized SCCmec (SCCmec I), and most were ST250. This sequence type is now archaic; a defective ccr complex prevented SCCmec I from transferring effectively to MSSA, causing the first MRSA strains to die out completely by the 1980s (Skov, 2011). While ST250 isolates no longer cause epidemic MRSA disease, a minor variant (ST247–MRSA-I; also known as the “Iberian clone”) is one of the major MRSA clones detected in European hospitals and has evolved resistance to most antimicrobial agents (Enright, et al., 2002). MRSA infections from this first wave were largely contained to Europe, with isolated reports of MRSA infection in the US (Chambers, et al., 2009; Skov, 2011).
Wave 2: Late 1970s – present
22
the 1990s (Skov, 2011). Both of these elements are very large (52 and 66 kb, respectively) due to the accumulation of multiple genes encoding resistance to antibiotics and heavy metals. While selective pressure for these elements exists within hospitals, their large size likely precluded them from ever gaining success in community-associated (CA) MSSA strains (Ma et al., 2002). Both of these SCCmec types continue to circulate globally, primarily in hospitals.
Wave 3: Late 1980s – present
The third wave of methicillin-resistance in S. aureus was marked by the emergence of the new, relatively small SCCmec type IV (20-24 kb). This MGE is uniquely devoid of genes encoding resistance to antibiotics or metals. However, its small size allows strains carrying this element to efficiently compete with natural human flora. SCCmec IV first emerged in novel hospital-associated (HA) clones CC22 and CC45, and then began replacing SCCmec II and III in CC5 and CC8 (Skov, 2011). By the 1990s, approximately 90% of the HA-MRSA circulating worldwide belonged to only five clonal complexes: CC5, CC8, CC22, CC 30, and CC45 (Skov, 2011). Many of these clonal complexes are still associated with HA infections.
Next, SCC mec IV began to transfer to MSSA lineages completely separate from typical HA strains. These new MRSA strains were able to multiply and spread outside of the hospital environment. Reports of CA-MRSA infections date back to the 1980s, but these strains were only able to truly establish themselves in the 1990s. Today, CA-MRSA infections are considered epidemic in many parts of the world, including the US (Kennedy, et al., 2009).
23
close contact with an individual colonized with CA-MRSA, contact sports, crowding or living in close quarters, intravenous drug use, and homosexual activity (Skov, 2011). CA-MRSA also tends to be less drug resistant than HA-MRSA, though this is highly strain-dependent. Importantly, CA-MRSA infections can easily occur in healthy individuals. Due to its increased fitness and virulence, CA-MRSA has begun to replace HA-MRSA in the hospital and long-term care facility settings, therefore blurring the traditional distinctions between “HA-MRSA” and “CA-MRSA.” Nosocomial outbreaks of CA-MRSA have been reported since 2003 (Otter et al., 2006).
Wave 4: 2004 – present
24
Livestock-associated ST398
Origins of MRSA ST398
Though MRSA has been described in livestock since 1972, ST398 represents the first known transmission of MRSA from livestock to humans (Price, et al., 2012). Evidence suggests that MRSA ST398 emerged due to the repeated introduction of SCCmec IV and V into precursor MSSA ST398 strains that existed in pigs and in their environment. These precursor strains are thought to be human in origin and were transmitted to pigs an indeterminable length of time ago (Price, et al., 2012). It is not clear whether or not the emergence of MRSA ST398 at varied geographic locations can be attributed to de novo emergence on multiple farms from multiple MSSA ST398 reservoirs, or whether transmission among farms and animals is the more probable cause (Smith, et al., 2011).
Characteristics of MRSA and MSSA ST398
25
rate nosocomial transmission of MRSA ST398 as 72% less likely than transmission of non-ST398 MRSA strains (Wassenberg et al., 2011). However, the possibility that transmission of ST398 will increase as the strain becomes better adapted to humans is of great concern. This is particularly alarming since the swine reservoir of ST398 is enormous, and likely cannot be eliminated. Importantly, pigs may not be the only livestock reservoir of ST398; carriage has also been detected in poultry, cattle, and horses (Nemati et al., 2008; Van den Eede et al., 2009; Vanderhaeghen et al., 2010).
ST398 Infection
MRSA and MSSA ST398 have been responsible for a wide diversity of minor and localized infections, including abscesses, urinary tract infections, wound infections, mastitis, and conjunctivitis (Smith, et al., 2011). Much more serious infections include bacteremia, pneumonia, osteomyelitis, otomastoiditis, and post-operative infections (Smith, et al., 2011). Currently, most ST398 infections present as skin and soft tissue infections (Smith, et al., 2011). The vast majority of ST398 infections have occurred in the last five years, although a disease-causing MSSA ST398 isolate was archived in Denmark in 1992 (Smith, et al., 2011).
26
ST398 better suited for human infection than traditional livestock-associated isolates (Uhlemann, et al., 2012).
Colonization versus Contamination
It has not clear whether occupational exposure to colonized pigs results in persistent ST398 carriage, or if individuals only become transiently contaminated. Several studies from the Netherlands suggest the latter, while a more recent study from Germany suggests the former. A 2009 study was the first to find that veterinarians who worked with pigs often tested positive for MRSA carriage, but only on the day of their visit (Van Den Broek et al., 2009). A longitudinal study conducted by Graveland et al. later found that MRSA carriage in veal calf farmers was strongly related to duration of animal contact, with odds of carriage significantly reduced during periods of low exposure (Graveland et al., 2011). A third study found veal farmers were 94% less likely to carry MRSA 24 hours after animal exposure, compared to immediately after exposure (van Cleef et al., 2011). However, a 2012 study found that 59% of German pig farmers did not clear MRSA colonization following summer holidays, suggesting that regular contact with swine may lead to persistent carriage (Robin Köck et al., 2012). Overall, these studies present conflicting evidence regarding whether or not MRSA ST398 “carriage” is an artifact of short-term exposure to MRSA-contaminated bioaerosols, or whether carriage truly represents persistent colonization.
ST398 in North America
ST398 and hog farms
27
though not all with ST398. Though a few ST398 infections have been reported, infection by ST398 remains rare in the greater Canadian population (estimated at 0.14%) (Golding, et al., 2010).
The only published report of MRSA ST398 on a US pig farm was from Iowa in 2009. Smith et al. investigated two conventional CAFO systems in Iowa, each comprising over 25,000 animals in total. In the first production system, 209 pigs were swabbed, 70% of which were colonized with ST398. Nine out of 14 workers also carried this same strain. Curiously, in the second production system, no pigs (90 sampled) and no workers (6 sampled) carried ST398 (Smith, et al., 2009). Hypotheses provided for this discrepancy included the confirmed importation of pigs from Canada by the first production system, the presence of nearly twice the number of hogs in the first production system compared to the second, and the older, more established nature of the first production system (Smith, et al., 2009). However, the authors did not research this discrepancy further. Prevalence of MRSA ST398 has not been further investigated in pigs or workers in CAFOs elsewhere in the United States.
ST398 and retail meat
28
(raised without the use of antibiotics) (O'Brien, et al., 2012). Over a quarter of the pork samples that tested positive for MRSA had spa types associated with ST398. The authors speculated that the presence of MRSA on alternative pork products might be a result of contamination during processing, as the study’s results were incongruous with previous findings in Europe (O'Brien, et al., 2012). Although S. aureus is destroyed by cooking, contact with contaminated meat could result in nasal colonization, cross-contamination of surfaces, and/or introduction of ST398 into the home environment.
Prevalence of ST398 in the US population
Carriage of MRSA ST398 in the general US population is unknown due to poor national surveillance, particularly in rural areas. However, no symptomatic infections in the US have been reported (Smith, et al., 2011), and all evidence suggests that prevalence of MRSA ST398 is sharply below that in many other Western, industrialized countries. This may be because the US has a high prevalence of CA- and HA non-ST398 MRSA, and these strains are outcompeting MRSA ST398 in the community European countries like Denmark suffered greatly from the emergence of ST398 because pre-existing MRSA rates were extremely low, due to effective “search and destroy” policies. However, these policies did not target pig farming as a risk factor for MRSA colonization prior to 2006, leaving healthcare systems vulnerable once ST398 emerged. Now, ST398 comprises 30% of Denmark’s MRSA cases, and the country is experiencing the highest rates of MRSA since 1985 (Statens Serum Institut et al., 2010).
29
contaminated with MSSA ST398, either as a human colonizer or on environmental surfaces. More than one human was colonized with MSSA ST398 in 20% of multi-person households contaminated with this strain; there were fewer concordant dyads for all other S. aureus strains examined (Uhlemann, et al., 2012). Another study from the Northeastern US recently described several infections in the Manhattan area due to MSSA ST398 and MSSA ST291, a novel sequence type belonging to CC398 (Mediavilla, et al., 2012). Most isolates were not multidrug-resistant, and all were acquired from individuals with no history of livestock exposure. Both of these studies suggest at least low-level prevalence of human-associated MSSA ST398 in the New York City area, and efficient transmission among households independent of livestock contact.
Unanswered Questions
30
risk to global public health. Some evidence shows that accretion of human-specific MGEs, including virulence genes, is already occurring (Skov, 2011). If transmission of ST398 among humans begins to rise, resistance to human-specific antibiotics may also become more prevalent. Serious infections by ST398, including endocarditis, bloodstream infections, and necrotizing pneumonia, may become more common if the fitness, virulence, and/or resistance of ST398 increase.
Multidrug-resistant
S. aureus
Multidrug-resistant S. aureus (MDRSA) is typically defined as S. aureus that demonstrates resistance to three of more classes of drugs. MDRSA can include MRSA, but multiply-resistant S. aureus that do not exhibit methicillin-resistance are common. Infections with such bacteria can present treatment challenges comparable to MRSA, and multidrug-resistant, pvl-positive MSSA strains are thought to be contributing to the ongoing epidemic of CA-MRSA infections in the US (Orscheln et al., 2009). However, prevalence and characteristics of general MDRSA carriage in the US population are unknown. Risks for MDRSA acquisition are also unknown, but are likely similar to risks for MRSA acquisition (See: Epidemiology of S. aureus carriage).
MDRSA Associated with CAFOs
31
facilities. Resistance to 4 antibiotic classes was evaluated; a significant difference was detected in S. aureus resistance to oxytetracycline (p=.010), tetracycline (p=.014), ampicillin (p=.007), and erythromycin (p=.035) when samples taken upwind and samples taken immediately downwind of the facilities were compared (Gibbs et al., 2006). Several other studies have found S. aureus to be the most prevalent microorganism captured by filtration and impaction of bioaerosols inside CAFO barns, although none of these studies have measured antibiotic resistance in S. aureus isolates (Butera et al., 1991; Crook et al., 1991; Predicala et al., 2002).
MDRSA has also been isolated from flies collected near poultry CAFOs in Maryland (Graham, et al., 2009). Antibiotic-resistant bacteria can be ingested by flies as they feed on decomposing waste and animal carcasses, or they can stick to their legs, feet, wings, or proboscis. Bacteria can then be transferred to new surfaces through regurgitation or physical contact. A 2009 study found that 10-15% of S. aureus isolates collected from flies trapped 100-200 m from poultry CAFOs were MDR (n=27), while approximately 30% of S. aureus isolates collected from nearby poultry litter piles were MDR (n=118) (Graham, et al., 2009). While the species of flies collected in this study have a typical traveling radius of 2 miles, individual flies can fly up to 20 miles. Flies carrying MDRSA present an additional route for human exposure.
Research rationale
32
strains of S. aureus resistant to methicillin (MRSA) have proven increasingly difficult to treat (Chambers, et al., 2009).
Industrial livestock operations may be a source of exposure to antibiotic-resistant S. aureus, including MDRSA and MRSA. In addition to confining hundreds of animals in enclosed spaces, industrial livestock operations administer 80% percent of all antibiotics sold in the US to food animals for the purposes of treatment, prophylaxis, and growth promotion (Food and Drug Administration, 2009). These practices have been associated with a growing animal and environmental reservoir of novel antibiotic-resistant pathogens, including antibiotic-resistant S. aureus (Frank M. Aarestrup, 1995; Gibbs, et al., 2006; Graham, et al., 2009). Some clones of antibiotic-resistant S. aureus can be zoonotically transmitted to individuals in close contact with livestock (Catry et al., 2010), who may then be at an increased risk for infection (Wertheim, et al., 2005). Of particular concern is the recent emergence of a new livestock-associated S. aureus clone, clonal complex 398 (CC398), which is responsible for an increasing percentage of MRSA infections in several countries, including Denmark, Germany, and the Netherlands (Smith, et al., 2011; Statens Serum Institut, et al., 2010).
33
of carrying antibiotic-resistant S. aureus, including MDRSA and MRSA, than workers and household members exposed to antibiotic-free livestock operations.
CHAPTER 2: METHODS
Study design and timeframe
All participants were recruited for this study by snowball sampling, beginning in early 2011. Industrial workers and household members were recruited with the help of two community organizations based in eastern North Carolina, the Rural Empowerment Association for Community Help (REACH) and the North Carolina Environmental Justice Network (NCEJN). ABF farmers and household members were mainly recruited by a team of researchers at the UNC Gillings School of Global Public Health, though a small number of ABF famers and household members were also recruited by REACH. Enrollment for this study began in May, 2011 and ended in December, 2011.
All workers employed in North Carolina were eligible to participate in this study, providing they reported contact with swine or poultry, were eighteen years or older, and spoke either English or Spanish. Up to two household members, including minors (ages 7-17), were also invited to participate if they reported living in the same household as a livestock worker for the past three months.
This study and all of its protocols were approved by UNC Chapel Hill’s Institutional Review Board. Recruitment materials and questionnaires were also available in Spanish.
Administration of Questionnaires and Nasal Swab Sample Collection
35
the case of ABF participants) to obtain questionnaire data and a nasal swab. The questionnaire assessed risk factors for MRSA colonization, including occupational exposures, household and environmental exposures, personal activities, and medical history, as well as basic demographics, including race, sex, and education level. Each member of a household completed a separate questionnaire.
Next, a swab of each participant’s nostrils was taken by the interviewer. A single, sterile BD BBLTM CultureSwabTM was used to swab both nostrils. Most interviewers were trained in proper swabbing protocol by a member of the nursing staff at UNC hospitals prior to the start of the study; all other interviewers were trained by an individual who had attended that training session. Swabs were immediately inserted into Stuart’s transport medium and stored at 4°C until arrival at the UNC School of Public Health. Swabs were transported by courier on ice in an insulated storage container within five days of collection.
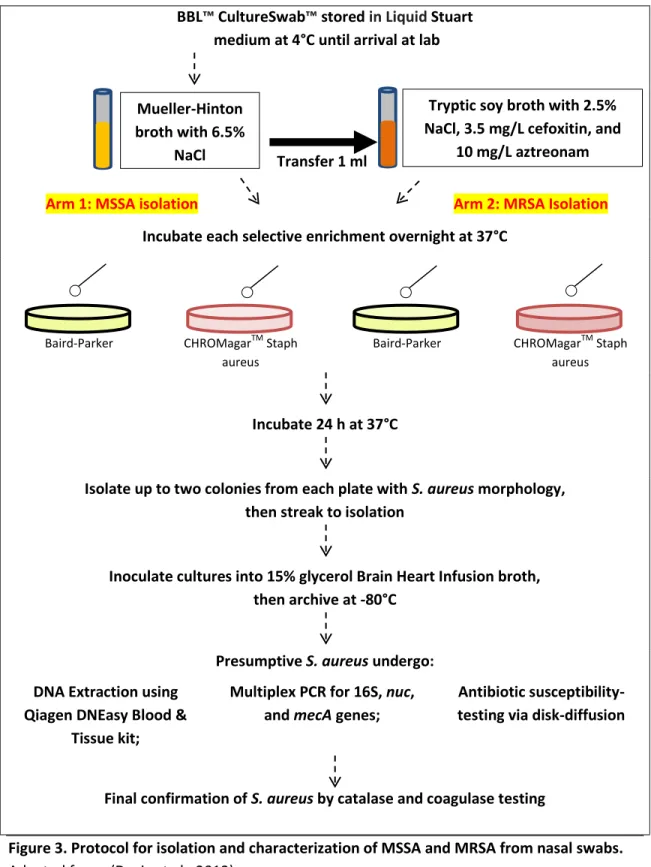
Isolation of MSSA and MRSA from Nasal Swabs
36
Tryptic soy broth with 2.5% NaCl, 3.5 mg/L cefoxitin, and
10 mg/L aztreonam Mueller-Hinton
broth with 6.5% NaCl
BBL™ CultureSwab™ stored in Liquid Stuart medium at 4°C until arrival at lab
Arm 1: MSSA isolation Arm 2: MRSA Isolation Incubate each selective enrichment overnight at 37°C
Baird-Parker CHROMagarTM Staph aureus
Baird-Parker CHROMagarTM Staph aureus
Incubate 24 h at 37°C
Isolate up to two colonies from each plate with S. aureus morphology, then streak to isolation
Inoculate cultures into 15% glycerol Brain Heart Infusion broth, then archive at -80°C
Presumptive S. aureus undergo: DNA Extraction using
Qiagen DNEasy Blood & Tissue kit;
Multiplex PCR for 16S, nuc, and mecA genes;
Antibiotic susceptibility-testing via disk-diffusion
Final confirmation of S. aureus by catalase and coagulase testing
Figure 3. Protocol for isolation and characterization of MSSA and MRSA from nasal swabs. Adapted from: (Davis et al., 2012)
37
exactly the same procedure as isolation of presumptive MSSA. A more detailed schematic of this process is provided in Figure 3. Approximately 10 swabs were processed each week using this method between May and December, 2011. This protocol was largely adapted from Davis et al. (2012).
Molecular Confirmation of MSSA and MRSA
38 Table 3. Primer sequences used for multiplex PCR
Gene target Direction Sequence (5’ -- 3’) Product Size (bp)
16S F GTG CCA GCA GCC GCG GTA A 886
R AGA CCC GGG AAC GTA TTC AC
nuc F TCA GCA AAT GCA TCA CAA ACA G 527
R CGT AAA TGC ACT TGC TTC AGG
mecA F GGG ATC ATA GCG TCA TTA TTC 255
R AAC GAT TGT GAC ACG ATA GCC
Source: (Poulsen, et al., 2003)
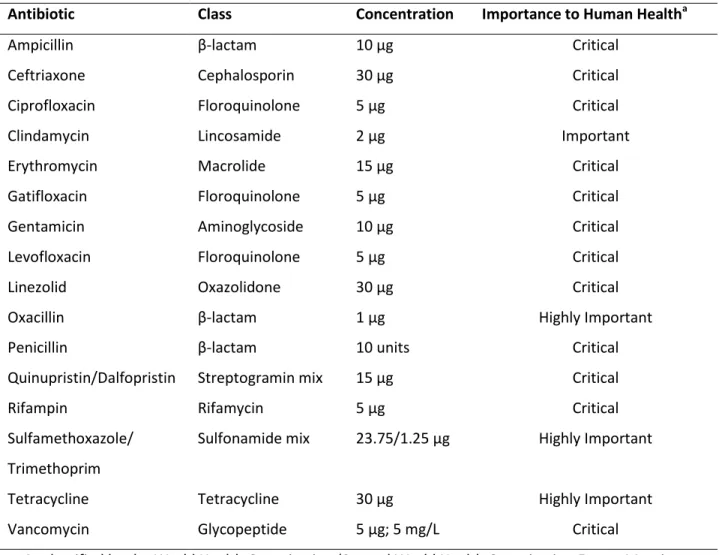
Antibiotic Susceptibility Testing
At least one isolate per S. aureus-positive individual (92 total) was assessed for its susceptibility to 12 classes of antibiotics using the Kirby-Bauer disk diffusion method (Table 4). Standard procedures were used and diameter interpretations were based on Clinical and Laboratory Standards Institute guidelines (Clinical and Laboratory Standards Institute, 2010). Inducible clindamycin resistance in erythromycin-resistant isolates was evaluated using the D-zone test (Steward et al., 2005). Additionally, vancomycin resistance was assessed in at least one isolate per S. aureus-positive individual (92 total) on Brain Heart Infusion agar supplemented with 5 mg/L vancomycin hydrochloride (Brown et al., 2005).
39
Table 4. Antibiotics used in susceptibility testing via disk diffusion
Antibiotic Class Concentration Importance to Human Healtha
Ampicillin β-lactam 10 µg Critical
Ceftriaxone Cephalosporin 30 µg Critical
Ciprofloxacin Floroquinolone 5 µg Critical
Clindamycin Lincosamide 2 µg Important
Erythromycin Macrolide 15 µg Critical
Gatifloxacin Floroquinolone 5 µg Critical
Gentamicin Aminoglycoside 10 µg Critical
Levofloxacin Floroquinolone 5 µg Critical
Linezolid Oxazolidone 30 µg Critical
Oxacillin β-lactam 1 µg Highly Important
Penicillin β-lactam 10 units Critical
Quinupristin/Dalfopristin Streptogramin mix 15 µg Critical
Rifampin Rifamycin 5 µg Critical
Sulfamethoxazole/ Trimethoprim
Sulfonamide mix 23.75/1.25 µg Highly Important
Tetracycline Tetracycline 30 µg Highly Important
Vancomycin Glycopeptide 5 µg; 5 mg/L Critical
a. As classified by the World Health Organization (Second World Health Organization Expert Meeting, 2007). Antibiotics were classified from lowest to highest priority: Important, Highly Important, and Critical.
Confirmation of CC398 and additional gene targets
40
PCR for the czrC gene was performed on all MRSA CC398 isolates identified by the lineage-specific PCR described above. Two µl of template was used in 2.5x 5 PRIME MasterMix (5 PRIME Inc., Gaithersburg, MD), with final primer concentrations of 2 pM. Primer sequences are described elsewhere (L. M. Cavaco et al., 2010). Amplification was carried out on a Bio-Rad Gene CyclerTM thermal cycler with the following program: 4 min at 94°C; then 30 cycles of 1 min at 94°C, 1 min at 55°C and 1 min at 72°C; followed by elongation for 10 min at 72°C. Amplified gene products were visualized as described above. A ST398 MRSA isolate (courtesy of Dr. Tara Smith, University of Iowa) was used a positive control and purified PCR-grade water was used as a negative control.
CHAPTER 3: RESULTS
Prevalence of
S. aureus
, MDRSA, and MRSA
In total, 105 antibiotic-free (ABF) farmers and household members and 99 industrial (IND) workers and household members participated in this study. Additionally, 30 industrial processing plant (PP) workers and household members participated; these individuals were separated from the IND category post-enrollment due to workers’ lack of contact with live animals. A detailed summary of study participants is presented in Table 5.
Table 5. Summary of study participants
Antibiotic-free (ABF)
Industrial (IND)
Processing plant (PP) Participant
Worker 92 80 18
Adult household member 4 10 8
Minor household member 9 9 4
TOTAL 105 99 30
42
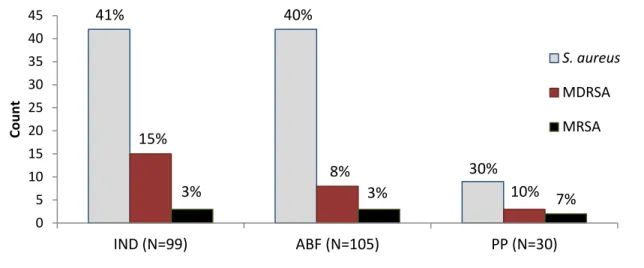
Figure 4. Carriage of S. aureus, multidrug-resistant S. aureus (MDRSA), and methicillin- resistant S. aureus (MRSA) in each study group.
Note: MDRSA and MRSA are sub-categories of S. aureus and are not independent of one another.
who had additional risk factors for MRSA carriage. MRSA prevalence in all three groups was higher than MRSA prevalence in the general US population, 1.5% (Gorwitz, et al., 2008).
Not all MRSA detected in this study were multidrug-resistant (Figure 5). Both positive individuals from the PP group carried multidrug-resistant MRSA while 2/3 of MRSA-positive individuals from the IND group carried multidrug-resistant MRSA. None of the MRSA isolates collected from the ABF group were multidrug-resistant.
MRSA isolated from one PP participant was pvl-positive (data not shown). No other tested S. aureus isolate carried the pvl gene.
Figure 5. Antibiotic resistance profiles of 8 MRSA isolates collected during study.
SXT: sulfamethoxazole/trimethoprim; VAN: vancomycin; LZD: linezolid; GEN: gentamicin; CLI: clindamycin; GAT: gatifloxacin; LEV: levofloxacin; CIP: ciprofloxacin; SYN: quinupristin/dalfopristin; ERY: erythromycin; TET: tetracycline; CEF: ceftriaxone; OXA: oxacillin; PEN: penicillin; AMP: ampicillin; MDR: multidrug-resistant 0 5 10 15 20 25 30 35 40 45
IND (N=99) ABF (N=105) PP (N=30)
Co u n t S. aureus MDRSA MRSA 8%
41% 40%
30% 15%
3% 3% 10% 7%
SXT VAN LZD RIF GEN CLI GAT LEV CIP SYN ERY TET CEF OXA PEN AMP MDR
43
It should be noted that the protocol used in this study failed to differentially isolate MRSA and MSSA from nasal swabs, as it was intended to do. Much of the “presumptive MRSA” isolated in Arm 2 of the study protocol (Figure 3) was in fact MSSA, as confirmed by PCR. It is hypothesized that MSSA were not completely eliminated during the overnight incubation in TSB with 2.5% NaCl, 3.5 mg/L cefoxitin, and 10 mg/L aztreonam. Once TSB enrichments were plated on to antibiotic-free BP and CS plates, any MSSA still remaining were able to flourish.
Resistance phenotypes
The antibiotic resistance profiles of S. aureus isolated from each study population were also assessed. A broad range of antibiotic resistance was observed in all three groups. Of the twelve classes of drugs tested (Table 5), resistance was observed to all except oxazolidones (linezolid), glycopeptides (vancomycin), and sulfonamides (sulfamethoxazole/trimethoprim). Rifamycin resistance was observed in only one ABF worker, while streptogramin resistance was observed in only one PP household member. Aminoglycoside resistance was unique to the IND group and was observed in two separate workers and an unrelated household member.
44
Figure 6. Percentage of S. aureus-positive ABF, IND, and PP participants demonstrating full resistance to 9 antibiotic classes.
Tetracycline resistance was unique to the IND group (χ²=21.92, df=2, p-value <0.0001) with over 40% of all S. aureus-positive IND participants demonstrating resistance to this antibiotic, compared with 22% of S. aureus-positive PP participants and 3% of S. aureus-positive ABF participants (Figure 6). Of the 19 IND participants who carried tetracycline-resistant S. aureus, 79% were workers who had direct contact with swine, while the remaining 21% reported direct contact with poultry.
Aminoglycoside resistance was also unique to the IND group, with resistance occurring in 2 IND workers and 1 IND household member, and no other study participant (Figure 6). 67% of individuals carrying gentamicin-resistant S. aureus reported direct or indirect contact with poultry.
Prevalence and Distribution of CC398
PCR was used to determine if any S. aureus isolated in this study belonged to CC398, the clonal complex commonly associated with zoonotic transmission. CC398 was detected in both workers and household members in this study, providing the first evidence of CC398 in humans
0 10 20 30 40 50 60 70 80 90 100 %
Antibiotic Free (N=42) Industrial (N=41) Processing Plant (N=9) 34
1
1 3 1 2 1
2 8 14 1 2 19 9 4 13
45
Figure 7. Prevalence of CC398 among S. aureus-positive IND, ABF, and PP participants.
in North Carolina (Figure 7). Seventeen of 41 S. aureus-positive IND participants carried CC398, as did 3/42 S. aureus-positive ABF participants. Ninety-four percent of CC398-positive participants were workers who reported direct contact with livestock. No CC398 was identified in the PP group.
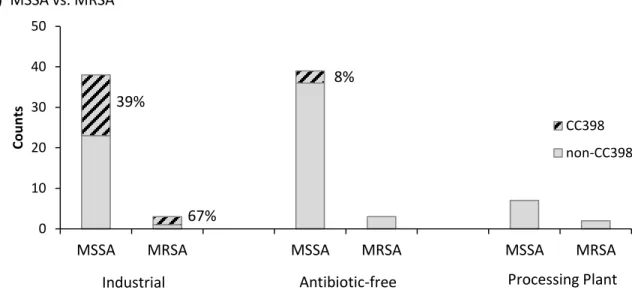
No MRSA CC398 was detected among ABF or PP participants, and only 2 cases of MRSA CC398 were identified among IND participants. Neither of these isolates carried the czrC gene. However, MSSA CC398 was present in both groups (Figure 8).
In the IND group, over 1/3 of all CC398 was resistant (Figure 8). No multidrug-resistant CC398 occurred in the ABF group.
0 10 20 30 40 50
Industrial Antibiotic-free Processing Plant
C
o
un
ts
CC398
non-CC398
41%
46 a) MSSA vs. MRSA
b) MDRSA vs. resistant to ≤2 classes of drugs
Figure 8. Distribution of CC398 among S. aureus-positive IND, ABF, and PP participants.
Resistance phenotypes of CC398 isolates
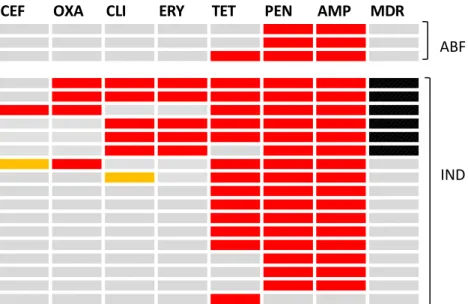
Nearly 1/3 of CC398 isolates collected in this study exhibited resistance patterns inconsistent with patterns characteristic of livestock-associated ST398. 1 ABF worker, 1 ABF household member, and 4 IND workers in this study were colonized with tetracycline-susceptible MSSA CC398, despite reporting direct contact with livestock (Figure 9). All six tetracycline-susceptible MSSA CC398 isolates demonstrated resistance to penicillin, and isolates from one IND worker demonstrated additional resistance to clindamycin and erythromycin.
0 10 20 30 40 50
MSSA MRSA MSSA MRSA MSSA MRSA
Co u n ts CC398 non-CC398 39% 67% 8% 0 10 20 30 40 50
non MDRSA MDRSA non MDRSA MDRSA non MDRSA MDRSA
Co u n ts CC398 non-CC398 40% 42% 9%
Industrial Antibiotic-free Processing Plant
47
Figure 9. Antibiotic resistance profiles of 20 CC398 isolates collected during study.
CEF: ceftriaxone; OXA: oxacillin; CLI: clindamycin; ERY: erythromycin; TET: tetracycline; PEN: penicillin; AMP: ampicillin; MDR: multidrug-resistant
Sensitivity and specificity comparisons
Results from two different media used to isolate S. aureus during this study are shown in Table 6. The sensitivity and specificity for detecting S. aureus from human nasal swabs using Baird-Parker media were 80.7% and 95.5%, respectively. Results from 216 individuals were included in these calculations because BP media was unavailable during one week of sample collection. The sensitivity and specificity of CHROMagar™ Staph aureus was 90.1% and 95.4%, respectively. Results from 222 individuals were included in these calculations for the same reason (unavailability of media). Nasal swabs were incubated overnight in selective enrichment broth prior to plating to increase sensitivity on both types of media (Safdar et al., 2003).
CEF OXA CLI ERY TET PEN AMP MDR
ABF
48
Table 6. Sensitivity and specificity of (a) Baird-Parker and (b) CHROMagar Staph aureus media for isolation of Staphylococcus aureus from nasal swabs
Baird-Parker CHROMagar™ Staph aureus
PCR + PCR - Total PCR + PCR - Total
Morphology + 67 6 73 Morphology + 82 6 88
Morphology - 16 127 143 Morphology - 9 125 134
Total 83 133 216 Total 91 131 222
Identifying colonies as presumptive S. aureus based on expression of only 2 out of 3 morphological characteristics was critical in ensuring that no S. aureus were missed. Of the 67 individuals correctly identified as S. aureus-positive using BP, 17.9% would not have been identified if only those colonies expressing all three characteristics (black, shiny, with clearing) were archived. Of the 82 individuals correctly identified using CS, 26.9% would not have been identified if only those colonies expressing all three characteristics (mauve, matte, with halo) were archived.
CHAPTER 4: DISCUSSION
This study provides the first report of CC398 in livestock workers and household members in North Carolina. MSSA CC398 was detected in both IND and ABF participants, although MRSA CC398 only occurred in industrial workers. We observed only 2 cases of MRSA CC398 among 99 sampled industrial participants, a sharply lower incidence than what has been observed in previous studies in North America (Khanna, et al., 2008; Smith, et al., 2009). Most cases of CC398 occurred in individuals with direct exposure to livestock.
Interestingly, 30% of CC398 detected in this study was tetracycline-susceptible. Tetracycline resistance is commonly considered a hallmark of livestock-associated ST398 (MSSA or MRSA) (Frank Møller Aarestrup et al., 2010), though resistance is rare among human-associated ST398 isolates (Uhlemann, et al., 2012). Human-human-associated ST398 detected in the US thus far has consistently demonstrated resistance to penicillin, and resistance to clindamycin and erythromycin is common as well (Mediavilla, et al., 2012; Uhlemann, et al., 2012). These findings suggest that CC398 in North Carolina may be different from livestock-associated CC398 isolated in previous studies. Whole genome sequence typing will be used to elucidate the evolutionary relationship of all CC398 identified in this study to a diverse collection of ST398 isolates described elsewhere (Price, et al., 2012).
50
the study groups were highly dissimilar – lacking collapsibility. The ABF group was mostly Caucasian, middle class, and college-educated, while individuals in the IND and PP group were largely Hispanic, of lower socioeconomic status, and less educated (data not shown). Additionally, there were wide differences in the distribution of traditional risk factors for MDRSA and MRSA colonization. Members of the ABF group were more likely to have used antibiotics within the past year, to have a pet inside the home, and to have recently visited a gym or played contact sports. However, it is important to that note our findings are in accordance with previous European studies which showed a higher prevalence of antibiotic-resistant S. aureus among industrial livestock workers compared with antibiotic-free livestock workers (Blaha et al., 2010; Graveland et al., 2010; Wulf, Nes, et al., 2008). Furthermore, despite having more risk factors for MDRSA and MRSA colonization, ABF participants had a lower prevalence of MDRSA than either the IND or PP group, and had a similar prevalence of MRSA.