Supporting Information
Wu et al. 10.1073/pnas.1113409109
SI Materials and MethodsPlant Materials. The mutant alleles and previously published transgenic plants used in this study werelfy-5,lfy-6,lfy-1(1), brm-101 (2), brm-1 (SALK_033046), brm-3 (SALK_088462), syd-2
(3),syd-5(SALK_023209), sep3-2(4),ap1-1(5),clf-2(6), LFY-GFP (7), ap1-1 cal-1 p35S::AP1-GR (8), and pAP3::GUS and
pAG-I::GUSreporter lines (9, 10). Forsyd-2pLFY::aMIRBRM, nine independent single-insertion site lines were analyzed, and all showed qualitatively similar phenotypes.
Constructs.ForpLFY::aMIRBRM, MIR167awas amplified from L.ergenomic DNA using primers 167a-F and 167a-R (below) and used as the backbone for generatingaMIRBRM.aMIRBRM
was generated by PCR mutagenesis using the following primers: 464-F1, 464-R1, 464-F2, 464-R2, 167a-F, and 167a-R (below). The 2.3-kbLFYpromoter (11) was fused toaMIRBRM, and the
pLFY::aMIRBRM fragment was subcloned into vector pBAR and transformed into L.erandsyd-2/+plants. Transgenic plants were selected on soil by spraying with the 0.5% herbicide BASTA (Crescent Chemical).
For pSYD::GFP-SYD, genomicSYD fragment from 2,372 bp upstream of the translational start codon to 133 bp downstream of the stop codon was subcloned into pENTR3C (Invitrogen) be-tween the SalI and EcoRV sites to obtain pENTR3C-gSYD. The junction between SYD promoter andfirst exon (agATGA; start codon is underlined) was mutated to agATCT by a mutagenesis kit (Invitrogen) to make a BglII site, and a GFP fragment was inserted. The new junctions were sequenced to confirm in-frame of GFP and SYD. The resultingGFP-SYDfragment was shuttled into the binary vector pGWB1 by LR clonase (Invitrogen).
ForpBRM::BRM-GFP, a genomicBRMfragment from 1,124 bp upstream of the translation start codon to 231 bp downstream of the stop codon was cloned into pENTR/D-TOPO (In-vitrogen). The stop codon was PCR-mutagenized to a SacI site to insert the GFP fragment. The junctions were sequenced to confirm in-frame fusion. The resultingBRM-GFPfragment was shuttled into the binary vector pMLBART.
For bimolecularfluorescent complementation (BiFC), the N-terminal domain of BRM (amino acids 1–976) and SYD (amino acids 1–657) plus the nuclear localization signal from VirD (12) were cloned into pCL113 to obtain p35S::cYFP-BRMN and
p35S::cYFP-SYDN. The mCherry coding region was cloned into pENTR/D-TOPO and shuttled into pEarley102 to obtain
p35S::2xmcherry(13, 14).
To express GST-LFY and GST-SEP3 inEscherichia coli, the full-lengthLFYandSEP3cDNA was inserted into EcoRI/NotI and BamHI/NotI sites of pGEX-5×-1 (GE Healthcare), respectively.
Gene Expression Analyses.Quantitative RT-PCR (qRT-PCR) was
yeast strain. Transformed cells were grown in −Trp −Leu/SD liquid media overnight, adjusted for equal cell density, serially diluted (1∼10−5), and spotted on selection media (−Trp−Leu −His/SD media) with or without various concentrations (0.1, 0.5, 1, and 2 mM) of 3-amino-1, 2, 4-triazole (3-AT; Sigma).
BiFC was performed by transforming proteins fused to the N-terminal half of YFP (nYFP) or to the C-N-terminal half of YFP (cYFP) into onion epidermal cells using the BioRad PDS-1000/ He system as per the manufacturer’s instructions. We tested both nYFP and cYFP fusions for individual bait proteins and ob-tained identical results. The negative control consisted of an unrelated nuclear localized protein described in ref. 14. The presumptive BRM complex component SWI3C (16, 17) was used as a positive control. p35S::2xmCherry was cotransformed as a transformation control. Fluorescent images of BiFC were taken using an Olympus MVX100 epifluorescence microscope.
GST pull-down assays were performed as previously described (18).
For coimmunoprecipitation experiments, nuclear extracts were prepared as previously described (19) from 1.5 g ofap1-1 cal-1 p35S::AP1-GR pSYD::GFP-SYD inflorescences 4 d after dexa-methasone induction. Two independent pSYD::GFP-SYD lines were used for the analysis. Nuclei were isolated in 10 mL of extraction buffer [100 mM 3-(N-morpholino) propanesulfonic acid (MOPS) (pH 7.6), 10 mM MgCl2, 0.25 M sucrose, 1%
dextran T-40, 0.25% Ficoll 400, 40 mM β-mercaptoethanol, 5 mM EDTA, and plant protease inhibitor mixture]. Nuclei were pelleted and resuspended in 500μL of disruption buffer [50 mM MOPS (pH 6.5), 1.5 mM MgCl2, 40 mMβ-mercaptoethanol, and
plant protease inhibitor mixture] followed by sonication. After the debris was removed by centrifugation, 10 μg of nuclear protein in 1 mL disruption buffer was precleared by incubating with 50μL of Dynabead protein G magnetic beads (Invitrogen) for 1 h at 4 °C. GFP-SYD was immunoprecipitated with 2μg (10
μL) of mouse monoclonal anti-GFP antibody (Invitrogen; A11120) overnight at 4 °C to precipitate GFP-SYD. Ten mi-croliters of 1× PBS (pH 7.0) was added to the no-antibody control. The immunocomplex was captured by 50μL of Dyna-bead protein G magnetic Dyna-beads for 2 h at 4 °C, and the Dyna-beads were washed with 1 mL of disruption buffer three times at 4 °C for 5 min. The immunocomplex was eluted with 30μL of 1×SDS loading buffer by incubating at 95 °C for 10 min, and all of the elutate was loaded onto 10% SDS-PAGE gel for Western blot analyses. Ten microliters of the nuclear extract from line A was run as input control. Rabbit polyclonal anti-LFY (20) (1:5,000 diluted) and anti-rabbit HRP conjugate (Amersham; NA934V; 1:5,000 diluted) were used as primary and secondary antibodies, respectively. Coimmunoprecipitated LFY protein was detected by Western blotting performed as described in ref. 19.
genetic background analyzed. Primers used in real-time PCR are listed below.
Primers Used.The primers used are described below.
qRT-PCR primers. EIF4-5′, AAACTCAATGAAGTACTTGAGGGACA; EIF4-3′, TCTCAAAACCATAAGCATAAATACCC; AGcDNA-5′, CAAAACTCCAACAGGCAATTG; AGcDNA-3′, CATTTTCAGCTATCTTTGCAC; AP3cDNA-5′, GCCCTAACACCACAACGAAGG; AP3cDNA-3′, CTCACCTAGCCTCTGCTTGATC; LFYRT-F, ACGCCGTCATTTGCTACTCT; LFYRT-R, ACGCCGTCATTTGCTACTCT. aMIR-BRM primers. 167a-F, CACCACTAGTCACTTTCGACCCTTAAACTCTC-CAT; 167a-R, CCGAGCTCCAAAGCTCCTCTTTCCTAGCTTCA; 464-F1, TTGGAGTTGCTGTTTGGTAAAATTAGCTTTCT-TTATCCTTTG; 464-R1, TTTACCAAACAGCAACTCCAATCAGATGCCGG-TGCACCATAG; 464-F2, TTTACCAAACACCAACTCCAACCCGTTGACTG-TCGCACCCTT; 464-R2, TTGGAGTTGGTGTTTGGTAAAATCGAGACTG-ATCTCTTAACC; AMIRBRM-F2, GGCATCTGATTGGAGTTGCT; AMIRBRM-R2, AACGGGTTGGAGTTGGTGTT. ChIP primers. pAG-F, TGAGTGATTGCCCAACTTGA; pAG-R, TGGTGGGTAGTTCTTGTGTGG; AGi1-F, GGGAAACAAATTGGGGAGAG; AGi1-R, CAACAATGGAGGATGGATGA; AGi2-F, CGTTGTGATGTTACTCGGACA; AGi2-R, CAACAACCCATTAACACATTGG; pAP3-F, CAAACGCCGTTAAGTTTGTC; pAP3-R, AGGGGGACCAAAGCTAAAAA; TA3-F2, CTGCGTGGAAGTCTGTCAAA; TA3-R2, CTATGCCACAGGGCAGTTTT; At4g18970-F, ACCGAATCGAATCCAAACAC; At4g18970-R, CACGGTAACAATGCGTTGAC; At3g54350-F, TCCGGGAAATAAGCATTTGA; At3g54350-R, CGTTTGAGAATGGAACGACA. Cloning primers. BRMins-F, CAGCAGCAGCACAGAAACTC; BRMins-R, CGACCTTGCTTGGTCTTCTC. 1. Weigel D, Alvarez J, Smyth DR, Yanofsky MF, Meyerowitz EM (1992) LEAFY controls
floral meristem identity in Arabidopsis.Cell69:843–859.
2. Kwon CS, et al. (2006) A role for chromatin remodeling in regulation of CUC gene expression in the Arabidopsis cotyledon boundary.Development133:3223–3230. 3. Wagner D, Meyerowitz EM (2002) SPLAYED, a novel SWI/SNF ATPase homolog,
controls reproductive development in Arabidopsis.Curr Biol12:85–94.
4. Liu C, Xi W, Shen L, Tan C, Yu H (2009) Regulation offloral patterning byflowering time genes.Dev Cell16:711–722.
5. Mandel MA, Gustafson-Brown C, Savidge B, Yanofsky MF (1992) Molecular char-acterization of the Arabidopsisfloral homeotic gene APETALA1.Nature360:273–277. 6. Goodrich J, et al. (1997) A Polycomb-group gene regulates homeotic gene expression
in Arabidopsis.Nature386:44–51.
7. Wu X, et al. (2003) Modes of intercellular transcription factor movement in the Arabidopsis apex.Development130:3735–3745.
8. Wellmer F, Alves-Ferreira M, Dubois A, Riechmann JL, Meyerowitz EM (2006) Genome-wide analysis of gene expression during early Arabidopsis flower development.PLoS Genet2:e117.
9. Sieburth LE, Meyerowitz EM (1997) Molecular dissection of the AGAMOUS control region shows that cis elements for spatial regulation are located intragenically.Plant Cell9:355–365.
10. Jack T, Fox GL, Meyerowitz EM (1994) Arabidopsis homeotic gene APETALA3 ectopic expression: Transcriptional and posttranscriptional regulation determinefloral organ identity.Cell76:703–716.
11. Blázquez MA, Soowal LN, Lee I, Weigel D (1997) LEAFY expression andflower initiation in Arabidopsis.Development124:3835–3844.
12. Relic B, Andjelkovic M, Rossi L, Nagamine Y, Hohn B (1998) Interaction of the DNA modifying proteins VirD1 and VirD2 of Agrobacterium tumefaciens: Analysis by subcellular localization in mammalian cells.Proc Natl Acad Sci USA95:9105–9110. 13. Earley KW, et al. (2006) Gateway-compatible vectors for plant functional genomics
and proteomics.Plant J45:616–629.
14. Ma Y, Slewinski TL, Baker RF, Braun DM (2009) Tie-dyed1 encodes a novel, phloem-expressed transmembrane protein that functions in carbohydrate partitioning.Plant Physiol149:181–194.
15. Winter CM, et al. (2011) LEAFY target genes revealfloral regulatory logic, cis motifs, and a link to biotic stimulus response.Dev Cell20:430–443.
16. Farrona S, Hurtado L, Bowman JL, Reyes JC (2004) The Arabidopsis thaliana SNF2 homolog AtBRM controls shoot development andflowering.Development131: 4965–4975.
17. Archacki R, et al. (2009) Genetic analysis of functional redundancy of BRM ATPase and ATSWI3C subunits of Arabidopsis SWI/SNF chromatin remodelling complexes.Planta 229:1281–1292.
18. Pastore JJ, et al. (2011) LATE MERISTEM IDENTITY2 acts together with LEAFY to activate APETALA1.Development138:3189–3198.
19. Su Y, et al. (2006) The N-terminal ATPase AT-hook-containing region of the Arabidopsis chromatin-remodeling protein SPLAYED is sufficient for biological activity.Plant J46:685–699.
20. Wagner D, Sablowski RW, Meyerowitz EM (1999) Transcriptional activation of APETALA1 by LEAFY.Science285:582–584.
21. Kwon CS, Chen C, Wagner D (2005) WUSCHEL is a primary target for transcriptional regulation by SPLAYED in dynamic control of stem cell fate in Arabidopsis.Genes Dev 19:992–1003.
22. Hurtado L, Farrona S, Reyes JC (2006) The putative SWI/SNF complex subunit BRAHMA activatesflower homeotic genes in Arabidopsis thaliana.Plant Mol Biol62:291–304. 23. Johnson L, Cao X, Jacobsen S (2002) Interplay between two epigenetic marks. DNA
methylation and histone H3 lysine 9 methylation.Curr Biol12:1360–1367. 24. Liu C, Thong Z, Yu H (2009) Coming into bloom: The specification offloral meristems.
Development136:3379–3391.
25. Krizek BA, Fletcher JC (2005) Molecular mechanisms offlower development: An armchair guide.Nat Rev Genet6:688–698.
26. Gross-Hardt R, Lenhard M, Laux T (2002) WUSCHEL signaling functions in interregional communication during Arabidopsis ovule development.Genes Dev16: 1129–1138.
27. Schmid M, et al. (2005) A gene expression map of Arabidopsis thaliana development. Nat Genet37:501–506.
boxed area) insyd-2 lfy-5andbrm-101 lfy-5compared withlfy-5inflorescences. (C)pWUS::GUSreporter (6) expression. No defect inWUSexpression was observed in youngflower primordia ofsyd-2 lfy-5andbrm-101 lfy-5relative tolfy-5. (D)WUSis strongly expressed in stamens and much reduced in all mutant
flowers that lack stamens. Expression data were obtained (7), and the fold change relative to stage 12flowers in the wild type is shown. The combined data suggest that the reduction inWUSis a consequence and not the cause of theAP3andAGreduction insyd-2 lfy-5andbrm-101 lfy-5.
Fig. S2. syd-2andbrm-101mutantflowers show weakfloral homeotic defects. (A) Wild-type (L.er),syd-2, andbrm-101flowers. An arrowhead (Right) in-dicates a petal partially transformed into a sepal. Outer whorl organs were removed to show inner organs in L.erandbrm-101. (Scale bars: 0.5 mm.) (B) Expression levels ofAG,PI, andAP3in wild-type (wt),syd-2 (syd), andbrm-101(brm) inflorescences. Expression of each gene in the wild type was set to 1. The error bars represent the SEM.
1. Sieburth LE, Meyerowitz EM (1997) Molecular dissection of the AGAMOUS control region shows that cis elements for spatial regulation are located intragenically.Plant Cell9:355–365. 2. Jack T, Fox GL, Meyerowitz EM (1994)Arabidopsishomeotic gene APETALA3 ectopic expression: Transcriptional and posttranscriptional regulation determinefloral organ identity.Cell
76:703–716.
3. Liu C, Xi W, Shen L, Tan C, Yu H (2009) Regulation offloral patterning byflowering time genes.Dev Cell16:711–722. 4. Liu C, Thong Z, Yu H (2009) Coming into bloom: The specification offloral meristems.Development136:3379–3391. 5. Krizek BA, Fletcher JC (2005) Molecular mechanisms offlower development: An armchair guide.Nat Rev Genet6:688–698.
6. Gross-Hardt R, Lenhard M, Laux T (2002) WUSCHEL signaling functions in interregional communication duringArabidopsisovule development.Genes Dev16:1129–1138. 7. Schmid M, et al. (2005) A gene expression map ofArabidopsis thalianadevelopment.Nat Genet37:501–506.
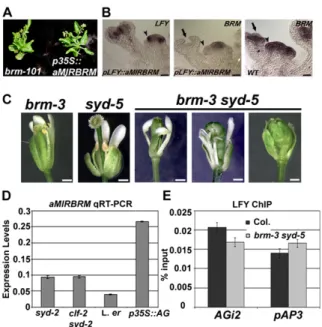
Fig. S3. Conditionalsyd brmdouble mutants. (A)p35S::aMIRBRMphenocopiesbrm-101null mutant. (B) Expression ofpLFY::aMIRBRMin wild-type plants did not alterLFYexpression (Left), but did specifically reduceBRMexpression infloral primordia compared with shoot meristem (compareCenterandRight panels). Stage 1 and 3floral primordia are indicated by arrowheads and arrows, respectively. (Scale bar: 30μm.) (C) Phenotypes of single- and double-mutant
flowers. Genotypes are indicated above. Double mutants betweensyd-2and thebrm-3intermediate allele show different degrees offloral homeotic defects. (Scale bars: 0.5 mm.) (D) Expression levels ofpLFY::aMIRBRMin the genotypes indicated. LFY binding toAP3andAGregulatory regions. Anti-LFY ChIP PCR in wild-type andsyd-2 pLFY::aMIRBRMinflorescences normalized overLFYlevels in the two genotypes. (E) LFY binding toAP3andAGregulatory regions. Anti-LFY ChIP PCR in wild-type (Col.) andbrm-3 syd-5inflorescences normalized overLFYlevels in the two genotypes.
Fig. S4. SYD and BRM bind LFY via the helicase/SANT-associated (HSA) and the HSA adjacent domain. (A) The N-terminal regions of SYD or BRM (SYDN or BRMN) can interact with LFY in yeast two-hybrid assays. (Left) Growth on media lacking tryptophan and leucine (−WL; not selecting for interaction). (Right) Growth on media that selects for interaction by further omitting histidine and adding 0.1 mM 3-AT for SYD or 0.5 mM 3-AT for BRM). BD, binding domain; AD, activation domain. (B) GST-LFY pull-down assays with different regions of SYDN, indicated by the amino acid numbers above. (Top) Prey protein levels. (Middle) GST pull-down. (Bottom) GST-LFY levels. The molecular mass is indicated on the right in kilodaltons. (C) Fine-mapping of the LFY-interacting regions in SYDN and BRMN by GST pull-down assays as shown inB. The QLQ and HSA domains are indicated above, and the HSA adjacent motif is represented by dashed boxes between these domains. Numbers indicate amino acid positions. Strong, weak, and no interactions are indicated by“++”,“+,”and“−,”respectively. (D) Amino acid alignments of the HSA domain (solid line) and the HSA adjacent motif (dotted line) of different angiosperm SWI2/SNF2 ATPases. The SYD orthologs are CHR3 (Arabidopsis thaliana), CHR910 and CHR925 (Populus trichocarpo), CHR6912 (Vitis vinifera), CHR720 (Oryza sativassp.japonica), and CHR2227 [Oryza sativa(indica cultivar group)]. The BRM orthologs are CHR2 (A. thaliana), CHR902 and CHR958 (P. trichocarpo), CHR6934 (V. vinifera), CHR20901 and CHR20918 (Ricinus communis), CHR1302 (Medicago truncatula), CHR707 (O. sativassp.japonica), CHR2214 [O. sativa(indica cultivar group)], and CHR131 (Zea mays).
Fig. S5. SEP3expression levels. In situ hybridization performed on early stage 3flower primordia of the wild type (L.er),lfy-6, andap1-1 lfy-6. Note the reduction ofSEP3expression in the center of theflower primordia inlfy-6relative to the wild type and inap1-1 lfy-6relative tolfy-6. (Scale bars: 30μm.)
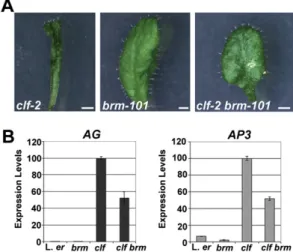
Fig. S6. BRM acts as a trithorax group protein. (AandB) Mutations inBRMcan overcome the defect caused by loss of polycomb repression. (A) Loss of BRM activity in thebrm-101null mutant restores leaf identity inclf-2mutants. Rosette leaves from 23-d-old plants are shown. (Scale bars: 0.5 cm.) (B) qRT-PCR ofAG andAP3expression in 13-day-old wild-type (L.er),clf-2,brm-10,1, andclf-2 brm-101seedlings. The error bar represents the SEM.
Fig. S7. Histone modification changes inclf-2 syd-2relative toclf-2. H3K27me3 (A) and H3K4me3 (B) ChIP data from Fig. 6Dbut with the wild-type andclf-2 single-mutant data omitted for better visualization of the difference in H3K27 and H3K4 methylation status ofAGandAP3regulatory regions inclf-2 syd-2relative toclf-2.