Dissertation. zur Erlangung des Doktorgrades (Dr. rer. nat.) der. Mathematisch-Naturwissenschaftlichen Fakultät. der
203
0
0
Full text
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
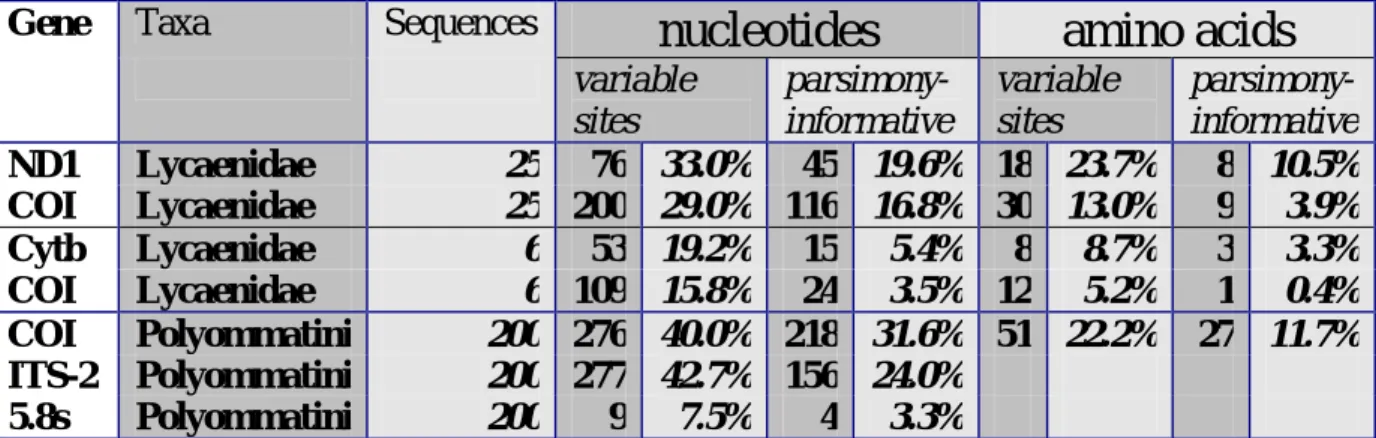
Figure



+7





Related documents
The FAO ABCDQ project (Agricultural Bulletin Board on Data Collection, Dissemination and Quality of Statistics) was established in 2000 to provide statistical metadata, on the
The results of this study showed that ethical culture and ethical climate (the informal ethical organizational context) have a much stronger effect on the frequency of observed
In the online supplements, we provide asymptotic theory for the case when objective functions are based on U-processes and verify regularity conditions for the maximum rank
Positive moment tension continuity in this connection was achieved by enlarging and relocating the dowel bars to the lower portion of the girder and extending them through
Meiosis: A form of cell division such that the daughter cells contain half the number of chromosomes as the parent cell.. –will be
In order to understand the influence of lateral membrane organization on the activity of the protein, the ATPase activity of LmrA reconstituted in to pure DMPC and a model
The Conservative Growth Option endeavors to maintain a 40% investment in corporate stocks and a 60% investment in bonds.. Stock ownership generally allows investors to grow their
