The Ameliorative Effects of
CE (A Chicken Extract) on
Learning and Memory Function
of Restraint-Stressed Mice
Copyright © 2016 INTRODUCTION
Numerous studies have revealed that psychological stress has a strong side effects on organism health and alters susceptibility to various diseases1,2,
including central nerve system (CNS), immune system, cardiovascular system and metabolic syndrome and so on. Generally, stressors injure organs and systems through the activation of the hypothalamic–pituitary–adreno-cortical (HPA) axis3 and the sympathetic nervous system (SNS)4. It has been
widely reported that stress triggers responses in HPA axis and triggering the releasing of glucocorticoids (GC), which activates its receptor (GR) and induces obvious effects on CNS, such as in the hippocampus, which is the main regions for cognitive functions formation. It has been reported that stress and GC can reduce hippocampal dendritic numbers and complexity and increase neuron apoptosis in hippocampal dentate gyrus5–7. Moreover,
there have been a number of studies suggesting that stress activates several central neurotransmitter systems and results in changes of neurotransmitters and their metabolites in brain or plasma. It is now well-established that these multiple changes of neurotransmitter systems in stress response are ulti-mately involved in various functional changes of brain, especially learning and memory function.
On the other hand, it is well-known that various food affected physi-ological function8. Chicken Essence (CE), which is an essence of chicken
extract and composed of water soluble substances extracted from chicken by double boiling, is a traditional health food among people in China and in Southeast Asia and is nowadays recognised as a popular health supplement. Recent studies have shown its various physiological roles, such as anti-anxiety, anti-stress, alleviating mental fatigue9,10, improving anemia and immune
function11–13. It has been reported that CE produced by Cerebos Pacific Ltd.
contains some beneficial ingredients such as carnosine, anserine, various amino acids, peptides and proteins14, as shown in Table 1.
In our laboratory, previous studies have demonstrated the protective effects of EC on stress and metabolic disorder of restraint-stressed mice through
Wen-Shan Huang1, Yi-Fang Li1,
Huan Chen1, Ting-Mei Wang1, Hiroshi Kurihara1,2, Rong-Rong He1,2*
1 Anti-Stress and Health Research center, College of Pharmacy, Jinan University, Guangzhou, China
2 Institute of Traditional Chinese Medicine and Natural Products, Jinan University, Guangzhou, China
Address reprint requests to *Rong-Rong He, Anti-Stress and Health Research center, College of Pharmacy, Jinan University, Guangzhou, China E-mail: [email protected]
Article citation: Huang WS, Li YF, Chen H, Wang TM, Kurihara H, He RR. The ameliorative effects of CE (a chicken extract) on learning and memory function of restraint-stressed mice. J Pharm Biomed Sci 2016;06(04): 259–263.
Available at www.jpbms.info
Statement of originality of work: The manuscript has been read and approved by all the authors, the requirements for authorship have been met, and that each author believes that the manuscript represents honest and original work.
Sources of funding: None.
Competing interest / Conflict of interest:
The author(s) have no competing interests for financial support, publication of this research, patents, and royalties through this collaborative research. All authors were equally involved in discussed research work. There is no financial conflict with the subject matter discussed in the manuscript.
Disclaimer: Any views expressed in this paper are those of the authors and do not reflect the official policy or position of the Department of Defense.
NLM Title J Pharm Biomed Sci CODEN JPBSCT
2230-7885 ISSN No
ABSTRACT
In this study, we studied the effects of Chicken Essence (CE) on learning and memory function of restraint-stressed mice. Male Kunming mice of 7-week-old were randomly divided into five groups as follows: normal control, restraint stress control, low dosage of CE (12 ml/kg/d, CE-L) and high dosage of CE (24 ml/kg/d, CE-H). The normal control group and restrain stress control group received water only. On the 14th day of admin-istration, all mice were conducted to step through training, and physically restrained in a 50 ml restrained tube with holes for 18 h except for normal control mice. All mice were conducted to step through testing, 1 day after restraint stress. All animals were anesthetized, their brains and blood were obtained. We determined the neuron protective transmitters, dopamine hydrochloride and norepinephrine level in brain and plasma via ESA–HPLC. The results showed that the administration of CE could improve the impaired function of learning and memory. Furthermore, it also recovered the changed levels of neurotransmitters in brain or plasma in restraint-stressed mice.
KEYWORDS learning and memory; restraint stress; stress hormone
histaminergicneurons15. Thus, in this study, we
investi-gated the effects of CE on learning and memory function in restraint-stressed mice and its related mechanism.
MATERIALS AND METHODS
Reagents
Norepinephrine (NE) and dopamine hydrochloride (DA) were purchased from Sigma (St. Louis, MO. USA. Diethyl ether was purchased from Guangdong Chemical Reagent Factory (Guangzhou, China). Chromatographic grade methanol and acetonitriles were purchased from Fisher (Iowa, USA).
Animals and regime
Male Kunming species mice of 7-week-old were acquired from the Center of Laboratory Animal Science Research of Southern Medical University (Guangdong, China). The animals were kept in a specific pathogen-free animal room at 23 ± 1°C and humidity conditions (50–70%) with a 12 h light–dark cycle (lights on from 6:00 to 18:00) under dim white light (about 15 Lux). Animals were allowed to acclimatise to the environment for 1 week before the experiment. Mice were randomly divided into normal control, restraint stress, low dose of CE (12 ml/ kg/d) and high dose of CE (24 ml/kg/d) group. The CE were concentrated to make sure the intakes of sample were 0.1 ml/10 g body weight. The normal control mice received saline water only, while restraint-stressed mice received gelatin, 7.2% gelatin in 0.3% caramel solution with the same caloric as CE. The care and treatment of the animals were conducted in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the United States National Institutes of Health (NIH publication no. 85–23, revised 1985),
and the experiment was conducted in accordance with animal ethics standards. The detailed experiment proce-dure is shown in Fig. 1.
Step-through passive avoidance task
Step-through passive avoidance task is performed with the JLBehv-STM4 video analysis apparatus (JLSOFTTECH Co. Ltd), which consisted of an illuminated and a dark compartment (each 20.3 × 15.9 × 21.3 cm) adjoin-ing each other through a small gate with a grid floor of 3.175 mm stainless steel rods set 8 mm apart. On the 14th day, mice were conducted to step through training
and then physically restrained in a 50 ml polypropylene centrifuge tube with holes for 18 h. After restraint stress, all mice were recovered for 24 h and conducted to step through testing. The interval between the placement of mouse in illuminated compartment and its first entry into dark compartment was recorded as error latency, which reflects memory retention. The entering events per testing trial were also recorded as the error counts. Then blood and brain tissues of mice were obtained under diethyl ether-anesthetised conditions.
Determination of NE in plasma by ESA–HPLC
Blood samples were rapidly taken from heart in diethyl ether-anesthetised mice. For plasma analysis, each tube contained 2% sodium heparin. The tubes were centrifuged at 5,000 rpm for 5 min. The supernatant was pipetted off and all samples were stored at −20°C until assay or mea-surements were conducted by ESA–HPLC. In short, the mobile phase contained methanol solution (100 ml/L) with citric acid monohydrate (71 mmol/L), trisodium citrate (69 mmol/L), EDTAg·2Na (0.123 mmol/L) and sodium octanesulphonate (0.83 mmol/L). After ultra-sonication, the mobile phase was filtered with 0.22 μm micropore film and disposed with ultrasonic degasifica-tion. Control for the temperature of the column was at 33°C, and the flow rate at 0.8 ml/min. Electrochemical detection was designed using two-way electrode poten-tials (passage I: −150 mV; passage IV: +450 mV).Measurement of neurotransmitters NE and DA
in the brain tissue
NE and DA were determined by ESA–HPLC. After the blood of mice was drawn, their brains were quickly removed and dissected into midbrain and brain cortex. These tissues were weighed and added with PBS solution
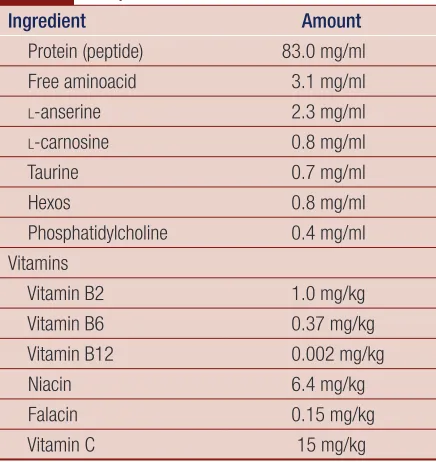
Table 1 Composition of Chicken Essence.
Ingredient Amount
Protein (peptide) 83.0 mg/ml
Free aminoacid 3.1 mg/ml
l-anserine 2.3 mg/ml
l-carnosine 0.8 mg/ml
Taurine 0.7 mg/ml
Hexos 0.8 mg/ml
Phosphatidylcholine 0.4 mg/ml Vitamins
Vitamin B2 1.0 mg/kg
Vitamin B6 0.37 mg/kg
Vitamin B12 0.002 mg/kg
Niacin 6.4 mg/kg
Falacin 0.15 mg/kg
Vitamin C 15 mg/kg
Sacrificed and sampled blood and brain tissue samples Restraint for 18 h
Administration of CE or water from 1 day to 14 days
1 day 14 days 15 days
were significantly decreased in restraint-stressed mice (20.46 ± 3.78 vs 34.36 ± 4.15), whereas with oral administration of different dose of CE could markedly recover NE level nearly to normal level (28.11 ± 3.17 and 30.83 ± 2.43, respectively), as shown in Fig. 3.
The plasma NE level was determined by ESA–HPLC, restrain stress lowered NE level in plasma and pre-CE administration could significantly improve that decre-ment of plasma NE in restrained stress mice. The data were shown as mean ± SD (n = 6) and the significance of difference from normal control group (normal) at **P < 0.01; from restraint stress group at ##P < 0.01.
Effect of CE on NE levels in the brain of
restraint-stressed mice
NE also belongs to neurotransmitters which play an important role for regulating nerve system functions. The measurements of NE in different brain regions were conducted by ESA–HPLC. Figure 4 shows that NE levels in the brain regions including cerebral cortex, hypo-thalamus and hippocampus were significantly decreased in restraint-stressed mice, and the administration of CE in advance showed a marked improvement, compared with stressed mice, even almost too normal level in these three brain regions of stressed mice.
NE levels in cerebral cortex, hypothalamus and hippocampus regions were conducted by ESA–HPLA, respectively. Restrained stress caused a significant decre-ment of NE levels in these three brain regions, while oral administrated different dosage of CE could effectively restored the low NE levels in restraint-stressed mice. The data were shown as mean ± SD (n = 6) and the signifi-cance of difference from normal control group (normal) at **P < 0.01; from restraint stress group (stress) at #P <
0.05 and ##P < 0.01.
Effect of CE on DA level in brain of
restraint-stressed mice
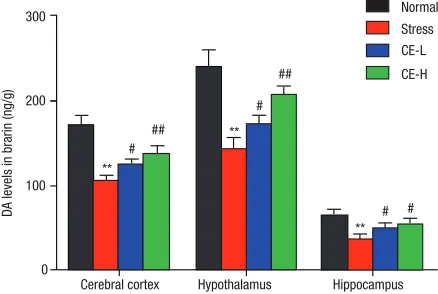
As another important neurotransmitter, DA is also involves in stress response and cooperates with other hormones and neurotransmitters to regulate neuroendocrine responses to stress. As shown in Fig. 5, when compared containing 3% perchloric acid whose final
concentra-tion was 1 g/ml. The materials were homogenised with a Physcotron homogeniser at the maximum setting level for 30 s under ice-cold condition. Then, the homogenate was centrifuged at 15,000 rpm for 20 min at 4°C. The supernatant was filtered through a 0.45 μm membrane filter and used as assay samples. And the following meth-ods for determining NE and DA levels were referred to that protocol for plasmic NE, as in section ‘Determination of NE in plasma by ESA–HPLC’.
Statistical analysis
The experimental data were given as mean ± SD. One-way analysis of variance (ANOVA) was applied to anal-yse the differences in the data of biochemical parameters among the different group followed by Dunnett’s test for pairwise multiple comparisons. The differences were considered as statistically significant at P < 0.05.
RESULTS
Effects of CE on learning and memory in
restraint-stressed mice
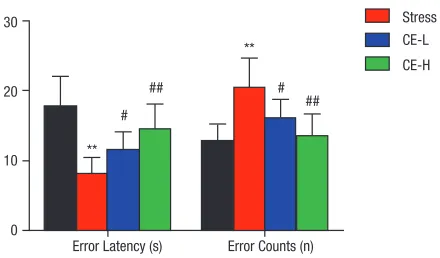
To investigate the effects of CE on learning and memory in restraint-stressed mice, CE at 12 and 24 ml/kg/d (CE-L and CE-H) were administrated to restraint-stressed mice by lavage in advance. As shown in Fig. 2, when mice were treated with restraint stress and recovered for 24 h, error latency was reduced and error counts were increased sig-nificantly, compared with normal control mice (8.20 ± 2.27 vs 17.86 ± 4.26 s, 20.50 ± 4.18 vs 12.88 ± 2.37 s). While, the administration of different dosage of CE (12 and 24 ml/kg/d) significantly recovered the error latency and counts, and caused a dose- dependent improvement of error latency and error counts in mice loaded with restraint stress. The results indicated the protective effects of CE on learning and memory behaviours in restraint-stressed mice.
Kunming mice were fixed in a restraint cage for 18 h after oral administration of different dosages of CE (12 and 24 ml/kg/d) or water for 14 days, then all the animals were conducted to perform step-through passive avoidance task with the JLBehv-STM4 video analysis apparatus on the next day of restraint stress, recorded animals, error latency and error counts. The data were shown as mean ± SD of values obtained from 10 mice in each group and the significance of difference was from normal control group at **P < 0.01; from restraint stress group at #P < 0.05 and ##P < 0.01.
Effect of CE on NE levels in plasma of
restraint-stressed mice
Concerning sympathetic-adrenmedulla system to stress response, NE level in plasma were measured by HPLC with an electrochemical detection; when compared with normal control mice, NE levels in the plasma
Fig. 2 Effect of CE on learning and memory in restraint- stressed mice.
30
20
** #
# ##
## **
Stress Normal
CE-L CE-H
10
0
with normal control mice, DA levels in the brain regions including cerebral cortex, hypothalamus and hippocam-pus were significantly decreased in restraint-stressed mice. The administration of CE caused a dose-dependent elevation of NE levels in the brain, especially in the cere-bral cortex and hypothalamus regions.
DA levels in cerebral cortex, hypothalamus and hippocampus regions were conducted by ESA–HPLA respectively. Restrained stress could significantly reduce DA levels in the cerebral cortex, hypothalamus and
hip-pocampus, while oral administrated different dosage of CE in advance could effectively restored the decrement of NE levels in restraint-stressed mice. The data were shown as mean ± SD (n = 6) and the significance of difference from normal control group (normal) at **P < 0.01; from restraint stress group (stress) at #P < 0.05
and ##P < 0.01.
DISCUSSIONS
In this study, we employed step-through passive avoid-ance task to investigate the effects of CE on learning and memory function of restraint-stressed mice. The results showed that, before stress, there was no difference at the latency or error counts between any two groups. When mice were treated with restraint stress, the latency was significantly reduced and the error counts were remark-ably increased. The results indicated that learning and memory of mice might be impaired by stress. Moreover, the administration of different dosage of CE could recover the latency and error counts to normal level, indicating the effects of CE on improving learning and memory function in restraint-stressed mice.
The brain is the master of behavioural and physio-logical process in stress response. The previous studies suggested the changes of central neurotransmitter sys-tems in brain, may account for the abnormal learning and memory function in restraint-stressed mice18,19. On
one hand, Selye pointed out the SNS plays an import-ant role and NE release from the SNS in the stress response20. As shown in our results, NE levels were
significantly decreased in cortex, hypothalamus, hip-pocampus and plasma, when mice exposed to acute restraint stress. The reduced NE levels may be related to the decreased activity of tyrosine hydroxylase during acute stress, which is a rate-limiting enzyme in the biosynthesis of NE. Moreover, it may be also due to the increase in its utilisation of NE, such as the increased activity of monoamine oxidase (MAO)21, whether this
alteration is involved in decreased biosynthesis or increased utilisation still needs to be further studied. After the administration of the positive drug or CE, NE levels were significantly increased in brain regions or plasma, indicating the effect of CE on NE system may be responsible for improving the learning and memory function impaired by restraint stress. The dopaminer-gic system is also very sensitive to stress and dopamine that impacts the emotion of organism plays a key role in learning and memory in the brain. As shown in our results, DA level was significantly decreased in the brain regions, including cortex, hypothalamus and hip-pocampus. There may be some relationships between restraint stress and DA metabolism, and the exact under-lying mechanism remains to be further verified. When mice were oral administered with positive drug or CE, DA level was remarkably increased in the brain, it indi-cated that the effects of CE on learning and memory may be also partly through its impact on DA system. Fig. 3 Effect of CE on plasma NE levels in mice loaded with
restraint stress. 50
40
30
20
10
** ## ##
0
Normal Stress CE-L CE-H
NE levels in plasma (ng/ml)
Fig. 4 Effect of CE on NE levels in the brain of restraint-stressed mice.
400
** # ##
**
** # ##
# ## 300
200
100
0
Cerebral cortex Hypothalamus
NE levels in brain (ng/g)
Hippocampus Stress Normal
CE-L CE-H
Fig. 5 Effect of CE on DA levels in brain of restraint-stressed mice. **
#
## **
** # # #
## 300
200
100
0
Cerebral cortex Hypothalamus
DA levels in brarin (ng/g)
Hippocampus Stress Normal
In conclusion, CE has ameliorative effects on learn-ing and memory function through modulatlearn-ing the neu-rotransmitters levels in restraint-stressed mice, while the modulated mechanisms still remained inconclusive. Otherwise, concerning CE contains so many kinds of nutrients, which play an important role in maintain-ing the homeostasis of organism through its anti-stress effects9–13, thus, the underlying mechanism may be also
related to its beneficial ingredients.
REFERENCES
1. Ader R. Psychoneuroimmunology. Curr Dir Psychol Sci. 2001; 10(3):94–98.
2. Cohen S, Herbert TB. Health psychology: psychological factors and physical disease from the perspective of human psycho-neuroimmunology. Ann Rev Psychol. 1996;47:113–42. 3. Haddad JJ, Saade NE, Safieh-Garabedian B. Cytokines and neuro-
immune–endocrine interactions: a role for the hypothalamic– pituitary–adrenal revolving axis. J Neuroimmunol. 2002;133(1–2): 1–19.
4. Madden KS. Catecholamines, sympathetic innervation, and immu-nity. Brain Behav Immun. 2003;17:S5–S10.
5. Conrad CD, McLaughlin KJ, Harman JS, Foltz C, Wieczorek L, Lightner E, et al. Chronic glucocorticoids increase hippocampal vulnerability to neurotoxicity under conditions that produce CA3 dendritic retraction but fail to impair spatial recognition memory. J Neurosci. 2007;27(31):8278–8285.
6. Kleen JK, Sitomer MT, Killeen PR, Conrad CD. Chronic stress impairs spatial memory and motivation for reward without dis-rupting motor ability and motivation to explore. Behav Neurosci. 2006;120(4):842–851.
7. MacPherson A, Dinkel K, Sapolsky R. Glucocorticoids worsen excitotoxin-induced expression of pro-inflammatory cytokines in hippocampal cultures. Exp Neurol. 2005;194(2):376–383. 8. Matsumura Y, Okui T, Ono H, Kiso Y, Tanaka T. Antihypertensive
effects of chicken extract against deoxycorticosterone acetate- salt-induced hypertension in rats. Biol Pharm Bull. 2001;24(10): 1181–1184.
9. Nagai H, Harada M, Nakagawa M, Tanaka T, Gunadi B, Setiabudi ML, et al. Effects of chicken extract on the recovery from fatigue caused by mental workload. Appl Human Sci. 1996; 15(6):281–6.
10. Xu C, Sim M. Effect of oral feeding of essence of chicken on the level of 5-hydroxyindoie acetic acid in the cerebrospinal fluid of the rat. Int J Food Sci Nutr. 1997;48(2):113–117.
11. Geissler C, Boroumand-Naini M, Harada M, Iino T, Hirai K, Suwa Y, et al. Chicken extract stimulates haemoglobin restoration in iron deficient rats. Int J Food Sci Nutr. 1996;47(4):351–360. 12. Man YC, Yee CW, Shing WK, Lai TP, Ching WK, Kei KK. The
enhanc-ing effects of a chicken-meat extract on serum Ig concentra-tions in normal and scalded animals. Br J Nutr. 2005;94(01): 51–55.
13. Candlish J. A traditional blood remedy as a modulator of the respiratory burst of the human neutrophil: an in vitro study. Int J Food Sci Nutr. 1998;49(1):55–63.
14. Geissler C, Boroumand-Naini M, Tomassen C. Large acute ther-mic response to chicken essence in humans. Nutr Rep Int USA. 1989;39:547–556.
15. Kurihara H, Yao XS, Nagai H, Tsuruoka N, Shibata H, Kiso Y, et al. The protective effect of BRAND’S Essence of Chicken (BEC) on energy metabolic disorder in mice loaded with restraint stress. J Health Sci. 2006;52(1):17–23.
16. Kulkarni MP, Juvekar AR. Effect of Alstonia scholaris (Linn.) R. Br. on stress and cognition in mice. Indian J Exp Biol. 2009;47(1):47–52. 17. Wong YN, Chien BM, D’Mello AP. Analysis of corticosterone
in rat plasma by high-performance liquid chromatography. J Chromatogr B Biomed Appl. 1994;661(2):211–218.
18. Cabib S, Puglisi-Allegra S. Opposite responses of mesolimbic dopamine system to controllable and uncontrollable aversive experiences. J Neurosci. 1994;14(5):3333–3340.
19. Rossetti ZL, Lai M, Hmaidan Y, Gessa GL. Depletion of meso-limbic dopamine during behavioral despair: partial reversal by chronic imipramine. Eur J Pharmacol. 1993;242(3):313–315. 20. Selye H. Stress without Distress. Toronto: McClelland and Steward
Ltd.; 1974. p. 62.