Molecular Pathways Differentiate Hepatitis C Virus (HCV) Recurrence from Acute Cellular Rejection in HCV Liver Recipients
10
0
0
Full text
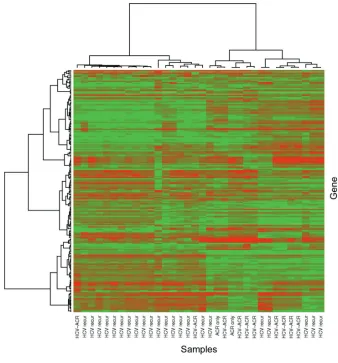
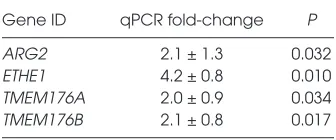
Figure



+2
Related documents