Acoustic communication in terrestrial and aquatic vertebrates
12
0
0
Full text
Figure
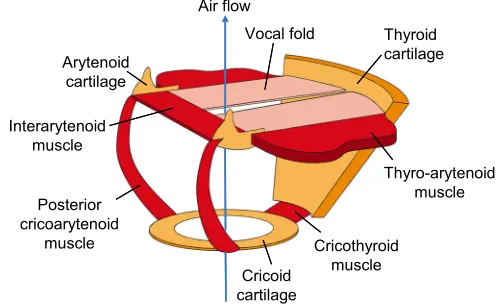
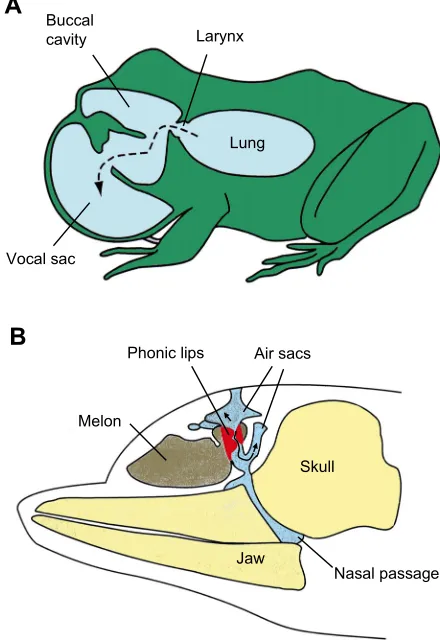


+2
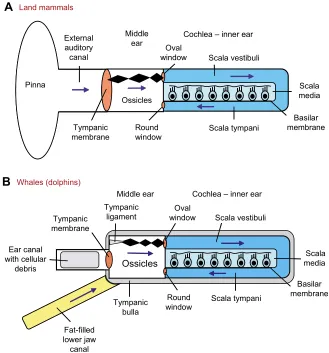
Related documents