21. T. D. Pennington, C. Reynel, A. Daza,Illustrated Guide to the Trees of Peru, D. Hunt, Ed. (Sherborne, Dorset, UK, 2004). 22. J. H. Torres-Romero, inFlora de Colombia., P. Pinto,
G. Lozano, Eds. (Universidad Nacional de Colombia, Bogotá, Colombia, 1988), chap. 5.
23. We thank M. Hall (Instituto Geofísico, Quito, Ecuador), A. Correa-Metrio and M. Bush (Florida Institute of Technology), and Erazo residents. We also thank the
reviewers for their valuable comments. This work was supported by funding from The Open University, Royal Botanical Garden Edinburgh, Santander, British Ecological Society, and University Utrecht.
Supporting Online Material
www.sciencemag.org/cgi/content/full/331/6020/1055/DC1 Materials and Methods
SOM Text Figs. S1 to S9 Tables S1 and S2 References
20 September 2010; accepted 14 January 2011 10.1126/science.1197947
A Terminal Pleistocene Child
Cremation and Residential Structure
from Eastern Beringia
Ben A. Potter,1*Joel D. Irish,1Joshua D. Reuther,2Carol Gelvin-Reymiller,1Vance T. Holliday2
The dearth of human remains and residential sites has constrained inquiry into Beringian lifeways at the transition of the late Pleistocene–early Holocene. We report on human skeletal remains and a residential structure from central Alaska dated to ~11,500 calendar years ago. The remains are from a ~3-year-old child who was cremated in a pit within a semisubterranean house. The burial-cremation and house have exceptional integrity and preservation and exhibit similarities and differences to both Siberian Upper Paleolithic and North American Paleoindian features.
E
vidence for the human colonization and early habitation of the Americas is pri-marily represented by stone tools and short-term camps and special task areas (e.g., work-shops, kill sites) (1, 2). Residential structures and human burials from the American late Pleis-tocene and early Holocene are rare; none are known for the North American Subarctic and Arc-tic. Here, we describe the discovery of human and cultural remains within a semisubterranean struc-ture at the Upward Sun River Site (USRS) in central Alaska (3). The human remains are those of a child who was cremated and buried, providing cultural information regarding early Beringians. USRS is located on a loess-mantled sand dune near the northern scarp of a terrace above the active Tanana River floodplain (Fig. 1). Exca-vations indicate that ~260 cm of loess overlies > 640 cm of aeolian sand (Fig. 2). Several thin paleosols (Ab horizons) 50 to 160 cm below the surface indicate brief breaks in loess sedimenta-tion. There are four cultural components each separated by 20 to 90 cm of sterile sediment; the upper three (Components 2 to 4) are associated with these paleosols (Fig. 2). The stratigraphy of these paleosols indicates little to no cryoturba-tion or other mixing processes in the occupacryoturba-tion layers, though there is evidence of limited cryo-turbation in the upper B horizons (B through Bwb2), above the burial (4) (Fig. 2). Geoarchae-ological details are provided in the supporting online material (fig. S1 and table S1). The burial and house are associated with Component 3, the third oldest at the site.The burial and house were identified dur-ing a 2010 excavation designed to explore the earliest late Pleistocene component. The remains were within a pit-hearth east of our main ex-cavation (Fig. 1). After consultation with gov-ernment and Native representatives, we excavated the entire feature and connected it with the Main Block through an 8-m trench, for a contiguous excavated area of 42 m2(Figs. 2 and 3).
The burial pit–hearth (Feature 5, Figs. 2 and 3) is an oval depression ~ 45 cm deep, measur-ing 130 cm by 100 cm at the top and 80 cm by 60 cm at the bottom. It is well defined by a thick
lens of charcoal and oxidized sediment 11 cm thick at the base and thinning on the sides. The pit edges flare out to form a flat, roughly cir-cular surface 280 cm across, delineated by a thin (~5- to 10-mm thick) layer of charcoal and gray-stained loess. Lithic artifacts and faunal remains are localized within this stain and exhibit clear arcs of debris to indicate the presence of a house wall and floor. The cross-section of this larger feature shows that it was a semisubterranean fea-ture dug about 27 cm below the contemporary ground surface (Fig. 2). Six apparent postmolds were observed as localized reddened stains, most with charcoal fragments and all with sharp edges in plan view and cross-section; four surround the pit-hearth. Overall minimum dimensions, arcs of debris among artifacts and fauna, and clear strat-igraphic integrity of the feature support the in-terpretation of a semisubterranean house (fig. S2). The pit-hearth consists of 15 to 37 cm of fill overlying a base composed of a 3-cm-thick oxi-dized layer with burned human and few faunal remains, a 1- to 3-cm-thick charcoal-rich layer, a 1-cm-thick ashy layer containing numerous burned fish and small-mammal bones, and a 3- to 4-cm-thick oxidized layer with abundant charcoal with some burned faunal remains. The human remains were above the fauna, in-dicating that the pit was not specifically created
1
Department of Anthropology, University of Alaska Fairbanks, Fairbanks, AK 99775, USA. 2School of Anthropology and Department of Geosciences, University of Arizona, Tucson, AZ 85721, USA.
*To whom correspondence should be addressed. E-mail: [email protected]
Fig. 1.Location of ancient North American human remains. (Inset) Upward Sun River Site map. Glacial limits for the earliest Beringian occupations (~14,100 cal yr B.P.) and time period of USRS occupation (~11,500 cal yr B.P.) are derived from (7). kya, thousand years ago.
25 FEBRUARY 2011 VOL 331 SCIENCE www.sciencemag.org
1058
for the burial, which represents the last use of the feature.
The human remains were concentrated toward the west/central part of the pit (Fig. 3); they comprise many variably burned bones that, al-though fragmented (most <2 cm across), were still in their approximately original position. Charred wood fragments (some 15 cm long) were interspersed with the human and faunal remains throughout the oxidized layers of the pit, representing the fuel source(s) of the cre-mation and earlier burning episodes. Two of these larger fragments arePopulus balsamifera (poplar). The spatial orientation of the human remains, artifacts, and contexts indicate that the pit was backfilled soon after interment, thus en-capsulating the find and facilitating preservation. Twenty14C samples from within the pit and overlying and underlying strata provide a secure chronology for this feature, and all four compo-nents (table S2). Two charcoal samples from the base of the pit (Beta-280585, 280586) were con-temporaneous with a date on the top of the pit fill (Beta-280584); all three average 9990T30 years before the present (yr B.P.) [11,620 to 11,280 calendar (cal) yr B.P.] (5). Two outdoor hearths are located in the same stratigraphic layer (~14 to 17 cm below Ab3) in the Main Block several meters to the west (Fig. 1); these yield similar ages. Three additional 14C samples from strata above the burial pit provide upper limiting ages, including two contemporaneous dates on Ab3 (Paleosol 2) averaging 8870T30 yr B.P. (10,170 to 9790 cal yr B.P.). In sum, the14C dating and stratigraphy support secure contexts for the pit and human remains. They are estimated at ~11,500 cal yr
B.P., at the end of the Younger Dryas chronozone, after the opening of the Ice Free Corridor and during a period when a land connection between northeast Asia and Alaska was still present or had only recently been inundated (6,7).
The burned human bones vary in color from black (Munsell 2.5Y 2.5/1), through gray/brown (2.5Y 6/1-2), to white (calcined) (2.5Y 8/1). Tem-peratures associated with these colors are 300° to 360°C, 400° to 525°C, and 645° to 800°C or above, respectively (8,9). Burning duration could have ranged from 1 to 3 hours (9,10). This het-erogeneity in combustion reveals that the skeletal elements were not extensively stirred in the fire. The posterior occipital is black, with a sharp line of demarcation to gray/brown (fig. S3). Parietal and remnant frontal fragments transition from gray/brown to white. Some long bone fragments are black, though most postcrania are lighter in color. Fragile facial and postcranial bones are largely absent. In agreement with the in situ map-ping, the individual was likely supine with the body inclined somewhat toward the right, on or near the hearth floor. The position of the body likely shielded the back of the head and part of the body from the hottest temperatures, and/or the black color resulted from direct contact with the soil, which can prevent bone carbonization (9). The body was angled relative to the long axis of the dune and nearby Tanana River, with the head oriented toward the southwest (~245°). The low-er limbs wlow-ere probably flexed, given the proxim-ity of the ribs near the edge of the pit and lack of preserved lower limb elements. Beyond the ana-tomically approximate in situ distribution of skel-etal elements, transverse fracturing and extensive
warping of all fragments suggest that soft tissue was present at the time of burning.
Less than 20% of the skeleton survived, in-cluding much of the posterior cranium, fragments of most deciduous and unerupted permanent teeth (i.e., unidentifiable enamel pieces, along with deciduous roots and incomplete permanent crowns protected by alveolar bone), and some postcrania. No indications of anomaly, pathol-ogy, or trauma are evident. The postcranial ele-ments primarily comprise ribs, some sections of upper limb cortical bone and, surprisingly, many fragile elements from both hands, positioned to the right of the axial elements. Most vertebral centra and arches, the scapulae, clavicles, innominates, and almost all bones of the legs and feet are absent. Some refitting is possible, but the postcrania are too incomplete to allow osteometric measure-ments for aging. However, the state of dental eruption (11) is consistent with an age of around 3T1 years (figs. S4 and S5). Variation in in-dividual tooth formation supports this estimate. Liversidge and Molleson’s (12) method for de-ciduous teeth yields a minimum of 1.98T0.31 years and a maximum of 3.48T0.69 years; most estimates are between 2.38T0.35 to 2.87T0.53 years. Similar findings were obtained with an alternative approach (13, 14) for deciduous and permanent teeth; on the basis of charts for males, the minimum age is roughly 1.9 yearsT 8 months and the maximum is 4 yearsT6 months; other ages lie between 2 and 3 years—with most near the higher end of this range. On the basis of these estimates, the USRS human remains rep-resent the second youngest individual of this pe-riod recovered in the Americas, after Anzick (15).
Fig. 2.Stratigraphic profiles and radiocarbon date locations for Upward Sun River. (Inset) Stratigraphic profile locations on the excavation grid. For radiocarbon dates, s denotes split samples. Illustrated strata comprise the uppermost lithostratigraphic unit. IV, upper silt loess. See SOM Text for sediment and soil descriptions.
www.sciencemag.org SCIENCE VOL 331 25 FEBRUARY 2011 1059
Although samples of deciduous or permanent teeth from populations are necessary to indicate biological affinity, some general information can be gleaned on an individual level. Slight shoveling and double shoveling in the unerupted permanent upper incisors and canines, among other diagnostic traits (16), are suggestive of a Sinodont pattern characteristic of Northeast Asians and Native Americans (17).
Skeletal sex determination of children is prob-lematic; in the present case it is impossible be-cause the most sexually dimorphic elements, i.e., mandible and innominates (18,19), are missing.
All identifiable faunal remains in this cul-tural layer were found within the pit, mostly below the human cremation. The assemblage is dominated by salmonid fish and small mammals (Table 1). The presence of ground squirrels with unfused epiphyses and salmon signify a mid-summer occupation. Multiple avian taxa were represented, primarily Tetraoninae (ptarmigan). Initial analyses indicate that complete carcasses were brought to the site and processed. Different elements from food animals are represented, and most fragments are <2 cm in size, likely due to differential preservation and fragmentation or burning (20,21). More complete element rep-resentation of microtines suggests that whole animals (or in the case of ground squirrels, the remains of cooked animals) were deposited in the pit/hearth. Differential burn temperatures,
lack of articulation, and high fragmentation sug-gest that some remains were not associated with the burial but accumulated during separate cook-ing or hearth-cleancook-ing events.
More than 350 lithic artifacts were found directly associated with the house feature, most (86%) concentrated near the eastern edge (Fig. 3). The debitage consists of generally small (79% are <1 cm) tertiary flakes, related to late-stage tool maintenance rather than tool production. Four used pieces are associated with the house and pit: a unifacially retouched flake, two flakes with light edge modification, and a biface.
Ad-ditional bifaces were in the Main Block (Fig. 4). Technological characteristics and local concen-trations of the artifacts suggest activity areas where tools were used and discarded. No evi-dence of grave goods (e.g., ornaments) was found, though two small ochre fragments were present within the pit-hearth near the human and may represent ritual activities at the time of burial.
Cultural chronologies for central Alaska are debated, but currently two broad interpretations for the late Pleistocene–early Holocene are as follows: (i) a single broad technological tradi-tion with variatradi-tion based on habitat use,
sea-A
B
Fig. 3.Plan view of house floor and burial. (Inset A) Spatial position of human remains. (Inset B) Stratigraphic profile of the house floor.
Table 1.Identified fauna from burial pit–hearth and house floor.
Taxon (common name) NISP % NISP % Burned % Complete
Pisces 286 37.9
Oncorhynchussp. (salmonid) 286 37.9 99 3
Mammalia 442 58.5
Arvicolinae (vole and microtine) 213 28.2 100 10
Urocitellus parryii(ground squirrel) 174 23.0 99 8
Lepus americanus(snowshoe hare) 50 6.6 100 8
Marmotasp. (marmot) 4 0.5 100 0
Sorexsp. (shrew) 1 0.1 100 0
Aves 27 3.6
Tetraoninae (ptarmigan/grouse) 16 2.1 100 18 Paridae (passerine) 10 1.3 100 10 Avian, unknown, possible Picidae 1 0.1 100 0
Total NISP 755 100.0
25 FEBRUARY 2011 VOL 331 SCIENCE www.sciencemag.org
1060
sonality, and mobility patterns (22–24); and (ii) multiple technological traditions represent-ing different populations (2,25,26). The USRS lithics, site structure, faunal data, and differences (e.g., lack of microblade technology, broader range of faunal resources, presence of residential fea-tures) relative to coterminous sites in the region (27, 28) support the former interpretation. Al-though the sample is small, the bipointed bifaces are similar to those of other Denali Complex specimens from Carlo Creek (29), Dry Creek Component 2 (30), and Houdini Creek (31) and unlike bifaces associated with the Nenana, Mesa, and Sluiceway Complexes (25,26,31,32). USRS Component 3 appears to be most closely asso-ciated with the regionally ubiquitous Denali Complex (or Paleoarctic tradition) (33).
Only one other ancient burial site is known for Beringia: Ushki Lake 1, in Kamchatka, Rus-sia (34–37) (Fig. 1). Ushki Lake 1, Level 7 (Ushki L7) (~13,000 cal yr B.P.) contained an adult burial associated with bone beads in a rock-lined ochre-filled pit separated from the house struc-tures. Ushki Lake 1, Level 6 (Ushki L6) (~12,000 cal yr B.P.) is roughly contemporaneous with USRS Component 3 and contains two unburned burials of children within two separate houses (35,36). One child burial contained ochre, a pen-dant, a mat of lemming incisors, and numerous microblades and wedge-shaped cores (the second burial is undescribed) (35). Thus, the USRS burial context is more like Ushki L6 than L7. This rep-licates technological linkages between continents: Diuktai Culture of Ushki L6 is comparable with the Denali Complex, which dominates the record from 12,000 to 6000 cal yr B.P. in interior east-ern Beringia (24,38), whereas the Ushki Cul-ture of Ushki L7, associated with stemmed points and lacking microblades, arguably has no direct counterpart in North America [(39), but see (34)]. North American human remains of this antiq-uity are uncommon (Fig. 1). Many were found in nonburial contexts, including the only other early eastern Beringian find, at On-Your-Knees
Cave (40); the latter is about 1000 calendar years younger than the USRS child (41). The USRS burial shares few similarities with Paleoindian burials from North America, including those within pits at Arch Lake, Buhl, Gordon Creek, Horn Shelter 2, Spirit Cave, Mostin, Whitewater Draw, and Wilson-Leonard II (2, 42, 43). No Paleoindian remains were found within houses, and cremations are known only from Marmes (44) and Spirit Cave (45). Associated artifacts interpreted to be grave goods vary by site, but ochre is relatively common. Child burials are rare in early contexts, though Anzick (Clovis age) con-tained a child with numerous ochre-scon-tained ar-tifacts (15). Thus, USRS appears more similar to burials in northeast Asia than in central North America, although samples are small.
The USRS semisubterranean house shares some similarities with the only other known Beringian site with houses: Ushki L6 (n= 12 houses). Floors at that site were excavated up to 50 cm (35) and ranged from 9 to 44 m2in area (36), sufficient for small groups of individuals. Both USRS and Ushki L6 contain child burials in pits within house floors, though the latter were not cremations. Differences include central stone-lined hearths (not pit-hearths) and entrance tunnels at Ushki L6. Six houses exhibit small pits within their floors, eight have postmolds around the periphery of the house floor, and four have them around the hearth (36). The USRS postmold distributions appear similar to those in Ushki L6 houses. Cache pits were found with-in Ushki L6 houses, with-includwith-ing one with dimen-sions similar to those of the USRS pit.
The fish- and small game–dominated assem-blage and few formal tools at USRS can be compared with the few other Beringian sites con-taining preserved fauna, particularly Broken Mammoth Cultural Zones 3 and 4 where birds and small mammals combined constitute 62% and 89% of the total number of identified spec-imens (NISP), respectively (46). Fish were found in small quantities at Broken Mammoth
Cul-tural Zone 3 (46). The pattern of broad-spectrum fauna with bifacial and expedient tools at these components can be contrasted with the micro-blade- and composite point–dominated assem-blage with multiple wapiti and bison at Gerstle River Component 3, a Denali Complex fall oc-cupation (27). The USRS evidence suggests that multiple subsistence strategies operated within a single archaeological tradition, the Denali Com-plex: (i) economic specialization, geared to cap-turing large ungulates in the context of logistically organized hunting groups and short-term camps; and (ii) broad-spectrum foraging in the context of local foraging parties near residential base camps. Similarities in location, house form, burial practices, and possibly fauna between Ushki L6 Diuktai Culture [also with large and small mam-mals, birds, and fish (34)] and USRS Component 3 suggest similar settlement and habitat use, i.e., residential occupations near large rivers accessing a broad spectrum of resources. However, notable differences between Ushki L6 and USRS com-plicate comparisons; at the former, semisubter-ranean houses were interpreted as winter dwellings interspersed with more ephemeral houses inter-preted to be summer dwellings, indicating year-round habitation (36).
A reasonable sequence of events at USRS can be inferred from these data. A small social group, including adult females and young chil-dren, foraged from their residential base camp in mid-summer, acquiring locally available fish, birds, and small mammals. The pit was dug with-in the house and functioned as a cookwith-ing hearth, cooking debris disposal area, and/or cache pit. The child died and was placed within the pit, with little evidence of disturbance after cremation. The pit was backfilled soon after burning, and the relative lack of artifacts atop the pit fill suggests immediate abandonment of the house (Fig. 3).
References and Notes
1. J. F. Hoffecker, S. A. Elias,Human Ecology of Beringia
(Columbia Univ. Press, New York, 2007).
2. E. J. Dixon,Bones, Boats, and Bison: Archeology and the First Colonization of Western North America(Univ. of New Mexico Press, Albuquerque, NM, 1999). 3. The site name is a translation of a nearby Athabaskan
placename,Xaasaa Na’. The site was previously named “Little Delta Dune.”The individual has been named
Xaasaa Cheege Ts’eniin(or Upward Sun River Mouth Child) by the local Native community.
4. B. A. Potter, J. D. Reuther, P. M. Bowers, C. Gelvin-Reymiller,
Curr. Res. Pleistocene25, 132 (2008). 5. All14C dates are calibrated with Intcal09 (47). 6. S. A. Elias, S. K. Short, C. H. Nelson, H. H. Birks,Nature
382, 60 (1996).
7. A. S. Dyke, A. Moore, L. Robertson,“Deglaciation of North America”(Geological Survey of Canada Open File, 1574, 2003). 8. P. M. Manye Correia, inForensic Taphonomy: The
Postmortem Fate of Human Remains, W. D. Haglund, M. H. Sorg, Eds. (CRC Press, Boca Raton, FL, 1997), pp. 275–293.
9. P. L. Walker, K. W. P. Miller, R. Richmond, inThe Analysis of Burned Human Remains, C. W. Schmidt, S. Symes, Eds. (Academic Press, London, 2008), pp. 129–136. 10. P. Shipman, G. Foster, M. Schoeninger,J. Archaeol. Sci.
11, 307 (1984).
11. D. H. Ubelaker,Human Skeletal Remains: Excavation, Analysis, Interpretation(Taraxacum, Washington, DC, ed. 2, 1989).
Fig. 4.Lithic tools from Component 3: bifaces (A,
E to H), modified flakes (BandC), unifacially re-touched flake (D). (B) to (D), and (F) were found in direct association with the house floor and buri-al pit; (A) was found ~13 cm below the house floor; and (E), (G), and (H) were found near hearth features in the Main Block.
www.sciencemag.org SCIENCE VOL 331 25 FEBRUARY 2011 1061
12. H. M. Liversidge, T. Molleson,Am. J. Phys. Anthropol.
123, 172 (2004).
13. C. F. A. Moorrees, E. A. Fanning, E. E. Hunt Jr.,Am. J. Phys. Anthropol.21, 205 (1963).
14. C. F. A. Moorrees, E. A. Fanning, E. E. Hunt Jr.,
J. Dent. Res.42, 1490 (1963).
15. S. Jones, R. Bonnichsen,Curr. Res. Pleistocene11, 42 (1994).
16. C. G. Turner II, C. R. Nichol, G. R. Scott, inAdvances in Dental Anthropology, M. S. Kelley, C. S. Larsen, Eds. (Wiley-Liss, New York, 1991), pp. 13–32. 17. C. G. Turner II,Am. J. Phys. Anthropol.82, 295
(1990).
18. H. Schutkowski,Int. J. Anthropol.2, 347 (1987). 19. S. R. Loth, M. Henneberg,Am. J. Phys. Anthropol.115,
179 (2001).
20. R. A. Nicholson,J. Archaeol. Sci. 20, 411 (1996). 21. J. L. Bennett,J. Archaeol. Sci.26, 1 (1999). 22. F. H. West, inAmerican Beginnings: The Prehistory and
Palaeoecology of Beringia, F. H. West, Ed. (Univ. of Chicago Press, Chicago, 1996), pp. 537–559. 23. C. E. Holmes,Arctic Anthropol.38, 154 (2001). 24. B. A. Potter,Radiocarbon50, 181 (2008).
25. J. F. Hoffecker, W. R. Powers, T. E. Goebel,Science259, 46 (1993).
26. T. E. Goebel, W. R. Powers, N. H. Bigelow, inClovis Origins and Adaptations, R. Bonnichsen, K. Turnmire, Eds. (Center for the Study of the First Americans, Corvallis, OR, 1991), pp. 49–79.
27. B. A. Potter,Environ. Archaeol.12, 3 (2007). 28. B. A. Potter, P. M. Bowers, J. D. Reuther, O. K. Mason,
Alaska J. Anthropol.5, 1 (2007).
29. P. M. Bowers,“The Carlo Creek site: Geology and archaeology of an Early Holocene site in the central Alaska Range” (Cooperative Park Studies Unit, Univ. of Alaska Fairbanks, 1980). 30. W. R. Powers, R. D. Guthrie, J. F. Hoffecker, Eds.,“Dry
Creek: Archeology and paleoecology of a Late Pleistocene Alaskan hunting camp”(U.S. National Park Service, Washington, DC, 1983).
31. M. R. Bever,Arctic Anthropol.38, 98 (2001). 32. J. T. Rasic, thesis, Washington State University, Pullman (2008). 33. F. Hadleigh-West,Am. Antiq.32, 360 (1967). 34. N. N. Dikov,Drevnie kul'tury severo-vostochnoi Azii[Early
Cultures of Northeastern Asia] (Nauka, Moscow, 1979, transl. by National Park Service, Anchorage, 2004). 35. S. B. Slobodin, inArchaeology of Northeast Asia on the
Pathway to Bering Strait, D. E. Dumond, R. E. Bland, Eds. (Univ. of Oregon, Anthropology Papers 65, 2006), pp. 9–23. 36. T. E. Goebel, S. B. Slobodin, inIce Age people of North
America: Environments, Origins, and Adaptations, R. Bonnichsen, K. L. Turnmire, Eds. (Oregon State Univ. Press, Corvallis, 1999), pp. 104–155.
37. T. E. Goebel, M. R. Waters, M. Dikova,Science301, 501 (2003). 38. J. P. Cook, thesis, Univ. of Wisconsin (1969). 39. S. B. Slobodin,Arctic Anthropol.38, 31 (2001). 40. E. J. Dixonet al.,Geoarchaeology12, 689 (1997). 41. E. J. Dixon, inProjectile Point Sequences in Northwestern
North America, R. L. Carlson, M. P. R. Magne, Eds.
(Simon Fraser Univ. Archaeology Press, Burnaby, Canada, 2009), pp. 11–18.
42. J. F. Powell,The First Americans: Race, Evolution, and the Origin of Native Americans(Cambridge Univ. Press, Cambridge, 2005).
43. D. W. Owsley, M. A. Jodry, T. W. Stafford,Arch Lake Woman(Texas A&M Press, College Station, TX, 2010). 44. G. Krantz,Northwest Anthropol. Res. Notes13, 159 (1979). 45. S. M. Wheeler,Nev. Hist. Soc. Q.40, 15 (1997). 46. D. R. Yesner,Quat. Sci. Rev.20, 315 (2001). 47. P. J. Reimeret al.,Radiocarbon51, 1111–1150 (2009). 48. This project was funded by the NSF (grants 0813819
and ARC-1057448). We thank J. Polston, First Chief of Healy Lake Tribal Council, and J. Isaacs, president and CEO of Tanana Chiefs Conference, for their support. We also thank the Northern Land Use Research, Inc., for field support; O. Davis and C. Alix (wood identification); and K. Blood and R. Bowman (laboratory assistance). We thank D. Meltzer, J. Hoffecker, and an anonymous reviewer for their comments.
Supporting Online Material
www.sciencemag.org/cgi/content/full/331/6020/1058/DC1 SOM Text
Figs. S1 to S5 Table S1 and S2 References
13 December 2010; accepted 26 January 2011 10.1126/science.1201581
The Developmental Role of Agouti
in Color Pattern Evolution
Marie Manceau,1,2Vera S. Domingues,1,2Ricardo Mallarino,1Hopi E. Hoekstra1,2*
Animal color patterns can affect fitness in the wild; however, little is known about the mechanisms that control their formation and subsequent evolution. We took advantage of two locally camouflaged populations ofPeromyscusmice to show that the negative regulator of adult pigmentation, Agouti, also plays a key developmental role in color pattern evolution. Genetic and functional analyses showed that ventral-specific embryonic expression ofAgoutiestablishes a prepattern by delaying the terminal differentiation of ventral melanocytes. Moreover, a skin-specific increase in both the level and spatial domain ofAgoutiexpression prevents melanocyte maturation in a regionalized manner, resulting in a novel and adaptive color pattern. Thus, natural selection favors late-acting, tissue-specific changes in embryonicAgoutiexpression to produce large changes in adult color pattern.
V
ariation in pigment type (i.e., color) and distribution (i.e., color pattern) can have a profound impact on the fitness ofor-ganisms in the wild (1). In vertebrates, several genes involved in pigment type switching (2,3) and those necessary for proper pigment
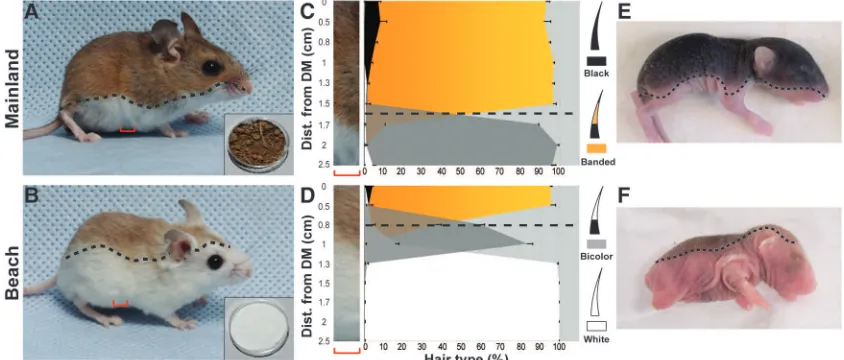
pattern-ing in mice (4,5) and fish (4,6,7) have been de-scribed; however, such work has focused on laboratory mutants rather than natural variation. Therefore, the molecular factors responsible for color pattern formation and evolution (i.e., the genes and developmental processes targeted by selec-tion) remain poorly understood in wild vertebrates. We took advantage of the striking color pat-tern variation in natural populations of deer mice (genusPeromyscus). Mainland mice (P. polionotus subgriseus) inhabit oldfields with dark soil and have the most common color pattern observed in vertebrates: a dark dorsum and light ventrum
1Department of Organismic and Evolutionary Biology,
Harvard University, Cambridge, MA 02138, USA.2Museum of Comparative Zoology, Harvard University, Cambridge, MA 02138, USA.
*To whom correspondence should be addressed. E-mail: [email protected]
Fig. 1.(AandB) Mainland and beach mice differ in coat color pattern, which provides camouflage in their respec-tive habitats (inset shows lo-cal soil sample). (CandD) Position of the boundary between the dorsal region, comprising banded and black hairs, and ventral region, com-prising bicolored or white hairs, in mainland and beach mice (black dashed lines). DM, dorsal midline; n= 5 for each subspecies. Error bars indicate SEM. (EandF) The position of the dorsoventral boundary is established be-fore birth (1-day-old pups).
25 FEBRUARY 2011 VOL 331 SCIENCE www.sciencemag.org
1062
www.sciencemag.org/cgi/content/full/331/6020/1058/DC1
Supporting Online Material for
A Terminal Pleistocene Child Cremation and Residential Structure from
Eastern Beringia
Ben A. Potter,
*Joel D. Irish, Joshua D. Reuther, Carol Gelvin-Reymiller, Vance T.
Holliday
*To whom correspondence should be addressed. E-mail: [email protected]
Published 25 February 2011,
Science
331
, 1058 (2011)
DOI: 10.1126/science.1201581
This PDF file includes:
1
Supporting Online Material:
Figs. S1 to S5
Tables S1 to S2
Site Formation and Disturbance
General characteristics of the sediments and soils at USRS are summarized in
Table S1 and Figs. S1-S2 (
S1
). Four lithostratigraphic units defined at the site consist of:
Unit I, >640 cm thick (260-900+ cm BS) fine-to-medium laminated sand; Unit II, 20 cm
thick (244-260 cm BS) massive silt; Unit III, 2-4 cm thick (240-244 cm BS) medium
sand; and Unit IV, 240 cm thick (0-240 cm BS) massive silt. Alluvial medium-to-coarse
sands and gravels were observed in lower interdunal areas at a depth of ~200 cm BS.
These descriptions confirm our observation that the dune ridge rests on an alluvial terrace
of the Tanana River.
Observed soils at the site are confined to the upper silt deposits (Units IV) and are
indicative of forest- and shrub tundra-derived soils (Typic Cryochrept and Cryorthents,
respectively;
S4
) typically formed in subarctic regions on areas of well-drained substrates
with sedimentation over 1 m. A series of forest soils are present in the upper 60-70 cm of
the sediment column. A modern Subarctic Brown Forest soil is represented by the
greatest amount of soil horizonation (OA-B-BC horizons) at the site with the upper 60 cm
of sediments. In some areas this soil is welded with at least two older buried incipient
forest soils (Bwb1 and Bwb 2 horizons). They are well expressed in the pit and house
excavation area between 25-50 cm BS and likely represent the establishment of the
spruce forest in the region during the Holocene around 9000 cal BP (
S5-7
). Horizon
boundaries within these upper forest soils (above the cultural components) range from
clear to abrupt in distinctness with topography of the boundaries consistently wavy to
slightly irregular; these features typically expresses seasonally frozen active layer
processes (cryoturbation) that affect forest soil horizons within subarctic regions.
The stratigraphy of paleosols below 80 cm BS in the Main Block and 50 cm BS in
the house and pit feature area indicate little or no cryoturbation or other homogenization
mechanism (e.g., pedoturbation, bioturbation) suggesting these features were not
substantially affected by post-depositional disturbance (Fig. S1). A discontinuous Ab
horizon (Ab3), designated Paleosol 2 (P2), is situated ~80 cm BS in the Main Block and
~50 cm BS in the house and pit feature area, overlying the top of the fill and house floor
by 5-15 cm. P2 developed by 10,170-9790 cal BP coinciding with a period in the pollen
record at nearby Birch Lake that displays a rise in
Populus
(
S6-7
), and may relate to the
establishment of a deciduous forest and beginning of forest soil development at USRS.
P2 boundaries are clear to abrupt in distinctness and wavy to slightly irregular in
topography likely due to minor cryoturbation and pedoturbation.
2
BP; the soils likely date to the older end of this range based on stratigraphy and presence
of younger soil-derived exogeneous carbon (i.e., humic acids) from upper soils, probably
Ab3 (see discussion of radiocarbon data in these supplemental data). This pedocomplex
underlies Component 3 in the main excavation area by 10 cm. The house floor overlies
the pedocomplex by approximately 40 cm, but the pit feature may have truncated part of
this pedocomplex. These soils are level across the site with smooth and very abrupt to
abrupt horizon boundaries that indicate little to no disturbance in this part of the lower
soil column. Spatially limited faunalturbation (krotovinas) and microfaulting were
observed in the excavations 4 m to the west of the house and pit feature and between 225
and 282 cm BS in depth; this location is >1 m below the lowest Ab4 soil (P1a) and the
house and pit features (
S9
). In sum, in the house and pit feature area, the effects of
turbative agents appear limited to the upper 50 cm of forest soils, i.e., above Ab3, well
above these features.
Site Chronology
USRS stratigraphy is linked by a series of 20
14C dates, consistent across the site
(Table S2). Component integrity is high, demonstrated by thin vertical distributions of
flat-lying lithics and faunal remains and ~20-90 cm of sterile sediments between
components. Both cultural feature and stratigraphic dates are available for Components 2
and 3, cultural feature dates for Component 1, and stratigraphic dates for Component 4
(
S7
).
Component 1 is situated 230 cm below surface and within Unit 2 (Lower Loess),
about 1 meter below Component 2 within Unit 2. Cultural materials consist of lithics and
faunal remains associated with a hearth feature, Feature 2. Two hearth charcoal fragments
were identified as willow (
Salix
sp.); one was a twig split and sent to two labs, both
yielding contemporaneous ages. A third, cross-check sample of willow was also
contemporaneous. All three dates were contemporaneous (t = 4.37,
χ
2= 5.99, df = 2, with
α
= 0.05) and average 11,320±30 BP (13230-13120 cal BP) (
S10
).
3
of Ab3 and Ab4 and the large suite of concordant dates, the hearth charcoal assays are
considered to date Component 2.
Component 3 is situated ~100 cm BS in the Main Block and ~80 cm BS in the
cremation area, and may be associated with a weak paleosol in the former area. All
Component 3 remains are in the same stratigraphic position, 14-20 cm below Paleosol 2
(Ab3). Cultural material consists of lithics and faunal remains associated with the house
(elements include Features 4, 7-8, 11-12) and pit (Feature 5) described in the main text
(Fig. S2) and three features interpreted to be outdoor hearths (Features 1, 3, and 9). Given
that there are no significant differences (see below) in tree and short-lived shrub species
(
Populus balsamifera
and
Salix
sp.), we consider old wood effect to be minimal. Two of
these hearth features (1 and 3) are directly linked through a lithic tool refit. Seven
radiocarbon dates have been run on three features, three on the cremation pit, and two
each on hearth Features 1 and 3, and several dates are contemporaneous. All three
cremation pit dates, two on charcoal at the base of the pit and one on charcoal at the top
of the fill, are contemporaneous (t = 1.12,
χ
2= 5.99, df = 2), yielding an average of
9990±30 BP (11620-11280 cal BP). This nearly overlaps with the Feature 1 crosscheck
dates, which are contemporaneous with each other (t = 0.80,
χ
2= 3.84, df = 1), and
average 9690±40 BP (11220-10870 cal BP). Feature 3 crosscheck dates are not
contemporaneous with each other (t = 0.80,
χ
2= 3.84, df = 1), but one of these two dates
(Beta-280914) overlaps with the cremation feature. Thus, of these seven dates, we
consider one (Beta-286265) as anomalous.
Component 4, consisting of a few lithic flakes, is situated ~80 cm below surface
in the Main Block in direct association with Paleosol 2 (Ab3), and about 20 cm above
Component 3. Both samples (from trees and shrubs,
Populus tremuloides
and
Salix
sp.)
from Paleosol 2 are contemporaneous (t = 0.22,
χ
2= 3.84, df = 1), and average 8870±30
BP (10170-9790 cal BP), providing effective upper limiting ages for Component 3.
The bottom of Bwb2 has a single assay, dating to 7600±40 BP (8510-8340 cal
BP), providing a lower limiting date on the formation of the upper Bwb horizons, which
are welded together and exhibit evidence of cryoturbation in the Main Block, but are
more distinct in the cremation area (see Fig. 2 of the main article).
In sum, the concordant suite of
14C dates, the mix of short-lived shrubs and tree
species yielding similar ages in various contexts suggesting minimal old wood effect, and
the clear stratigraphy that can be followed over many meters provide secure
chronological controls on the components at USRS.
Human Remains
4
Overall dental eruption (
S14
) is most consistent with an age of 3 years ± 1 year.
Individual tooth formation is supportive of this estimate. The method of Liversidge and
Molleson (
S15
) yields a minimum age of 1.98 ± 0.31 years based on full closure of the
lower left deciduous central incisor’s root apex (i.e., their grade of H2); open root apices
of both upper deciduous second molars indicates a maximum age of 3.48 ± 0.69 years
(between G-H1). As noted in the text, age estimates from other deciduous tooth roots
using this method are between 2.38 ± 0.35 to 2.87 ± 0.53 years. Moorrees et al. (
S16-17
)
derived tooth formation standards for both deciduous and permanent teeth. Their charts
for male children suggest the minimum age is roughly 1.9 years ± 8 months for the lower
right deciduous first molar, whose anterior root is not fully closed (between their grade of
A1/2-C). However, their maximum age, based on crown formation in the lower
permanent first premolar crowns (see Fig. S4) is around 4+/-1 years (between C1/2-3/4);
other ages using this method lie between 2 to 3 years – with most near the higher end of
the range.
With regard to the child’s possible biological affinity, several dental
morphological traits are suggestive (though see caveats in the text concerning assessment
of affinity in individuals). Slight shoveling, approximately ASUDAS Grade 2 (Fig. S5),
and double shoveling are evident in the unerupted permanent upper incisors and canines,
respectively. These and other diagnostic traits (
S18
), including what would likely have
been a large (ASUDAS 2+) deflecting wrinkle and anterior fovea, on the undeveloped
permanent lower left first molar crown (Fig. S4), are consistent with the Sinodont pattern
of Northeast Asians and Native Americans (
S19
).
References and Notes
S1. Soil descriptions follow national conventions established by
S2
, modified by
S3
S2. USDA Soil Survey Division Staff. “Soil survey manual.” (U.S. Department of
Agriculture Handbook 18, Soil Conservation Service, Washington D. C., 1993).
S3. Holliday, V. T.
Soils in archaeological research
. (Oxford Univ. Press, Oxford,
2004).
S4. Rieger,
S.
The Genesis and classification of cold soils
. (Academic Press, New York,
1983).
S5. Bigelow, N. H., thesis, University of Alaska Fairbanks (1997).
S6. Ager, T. A., L. B. Brubaker, In
Records of Late Quaternary Northern American
sediments
, B. Vaughn, Jr., F. Holloway, Eds. (American Association of
Stratigraphic Palynologists, Dallas, 1985), pp. 353-384.
S7. Bigelow, N. H., W. R. Powers, Climate, vegetation, and archaeology 14,000-9000
cal yr B.P. in Central Alaska.
Arctic Anthropology 38
:171-195.
S8. Dilley, T. E., thesis, University of Arizona, Tuscon (1998)
S9. Potter, B. A., J. D. Reuther, P. M. Bowers, C. Gelvin-Reymiller. Little Delta Dune
site: a Late Pleistocene multi-component site in Central Alaska.
Current Research
in the Pleistocene
25:132-135 (2008).
5
S11. Stuiver, M., Reimer, P. J., and Reimer, R. W. 2005. CALIB 5.0. [WWW program
and documentation].
S12. Reimer P. J., et al., IntCal09 and Marine09 radiocarbon age calibration curves, 0–
50,000 years cal BP.
Radiocarbon
51
:1111–50 (2009).
S13. Ward, G. K., S. R. Wilson, Procedures for comparing and combining radiocarbon
age determinations: A critique,
Archaeometry
20
:19-31 (1978).
S14. Ubelaker D. H.,
Human skeletal remains: excavation, analysis, interpretation
, 2nd
Ed. (Taraxacum, Washington, D.C., 1989).
S15. Liversidge, H. M., T. Molleson, Variation in crown and root formation and eruption
of human deciduous teeth.
American Journal of Physical Anthropology
123
:172-180 (2004).
S16. Moorrees C. F. A, E. A. Fanning, E. E. Hunt, Formation and resorption of three
deciduous teeth in children.
American Journal of Physical Anthropology
21
:205-213 (1963).
S17. Moorrees C. F. A, E. A. Fanning, E. E. Hunt, Age variation of formation stages for
ten Permanent teeth.
Journal of Dental Research
42
:1490-1502 (1963).
S18. Turner, C. G. II, C. R. Nichol, G. R. Scott, In
Advances in dental anthropology
, M.
S. Kelley, C. S. Larsen, Eds. (Wiley-Liss, New York, 1991), pp. 13-32.
S19. Turner C. G. II, Major features of sundadonty and sinodonty, including suggestions
about East Asian microevolution, population history, and Late Pleistocene
relationships with Australian Aboriginals.
American Journal of Physical
6
Table S1. General Description of Stratigraphy at Upward Sun River.
Lithostratigraphic
Unit
Pedostratigraphy Description
IV Upper Silt
(Loess) (0-240
cmBS) mostly
massive yellowish
brown (10YR 5/4)
to pale brown
(10YR 6/3) silt;
two fine sand
laminations occur
around 160-180
cmBS
[Component 3]
O
2-18 cm thick; modern vegetative growth and slightly to
moderately decomposed organics; abrupt and wavy boundary
A
6-26 cm thick; dark brown (7.5YR 3/2) to yellowish brown
(10YR 5/4) loam; abrupt and wavy boundary
B
26-37 cm thick; dark yellowish brown (10YR 4/6) to
yellowish brown (10YR 5/4) silt loam; welded onto and
turbated with Bwb1 and Bwb2 at the eastern and western most
edges of the house feature area and within most of the Main
Block; clear and wavy boundary
BC
4-10 cm thick; yellowish brown (10YR 5/6) silt to silt loam;
slightly welded onto and turbated with Bwb2 and Ab1 (P2) at
western end of the house feature excavation area; abrupt to
clear and smooth to wavy boundary
Bwb1
7-22 cm thick; brown (7.5YR 4/3 to 4/4) silt loam; abrupt to
clear and smooth to wavy boundary
Pedocomplex
Bwb2 (Bwb2a
and Bwb2b)
5-13 cm thick; brown (7.5YR 4/3 to 4/4) silt loam; slightly
welded onto P2 at far eastern end of house feature excavation
area; Bwb2 bifurcates above the pit feature in the house into
two possibly two distinct horizons (Bwb2a and Bwb2b);
abrupt to clear and smooth to slightly wavy boundary.
Ab3 (Paleosol P2)
0.5-5 cm thick; dark yellowish brown (10YR 4/4)
discontinuous Ab horizon; appears as a Bwb at its thickest
point at the western edge of the structure excavation area;
contains Component 4
; lowest limit of field observed minor
cryo- and pedoturbation at the site; very abrupt to abrupt and
smooth to slightly wavy boundaries;
Ab4
(Pedocomplex P1)
0.5-1 cm in thickness; at least six dark yellowish brown
(10YR 4/4) discontinuous Ab horizons; P1a and P1b soil
couplet are the best expressed of these six Ab2 horizons; P1a
and P1b are separated by 5 cm of pale brown (10YR 6/3)
unaltered silt;
contains Component 2
; very abrupt and
smooth boundaries;
III Aeolian sand (240-244 cm BS)
2-4 cm thick; grayish brown (10YR 5/2) massive
fine-to-medium sand; no lamination present; very abrupt and smooth
boundary;
II Lower Silt (Loess) (244-260 cm BS)
16-20 cm thick; yellowish brown (10YR 5/4) to pale brown
(10YR 6/3) massive silt;
contains
Component 1
; very abrupt
to abrupt and smooth boundary
7
Table S2. Radiocarbon assay data (ordered by strata and block; Block H refers to the
cremation area)
Lab Material Context (block and depth) δ13 C (‰)
Conv. age (BP)
Calibrated age (IntCal09) (2 σ cal BP) Beta-280581 charcoal (Juniperus sp.) Bottom of B horizon, H,
47 cm BS
-23.9 7600±40 8508-8343
Dates associated with Component 4
Beta-232392 charcoal (Salix sp.) Paleosol 2, Main, 78 cm BS
-23.3 8880±40 10,179-9789
Beta-280582 charcoal (Populus sp., probably P. tremuloides)
Paleosol 2, H, 57 cm BS -26.3 8850±50 10,168-9736
Beta-219656 charcoal From test pit, 76 cm S -24.8 9360±70 10,756-10,299
Dates Associated with Component 3
Beta-220218 hearth charcoal (Salix sp.) Feature 1 hearth, Main, 100 cm BS
-24.8 9650±60 11,200-10,775
Beta-280913 hearth charcoal (Populus balsamifera)
Feature 1 hearth, Main, 100 cm BS
-25.2 9720±50 11,242-10,830
Beta-286265 hearth charcoal (Populus balsamifera)
Feature 3 hearth, Main, 100 cm BS
-25.4 9030±40 10,248-10,163
Beta-280587 hearth charcoal (Populus balsamifera)
Feature 3 hearth, Main, 100 cm BS
-24.1 10130±50 12,012-11,408
Beta-280584 Pit charcoal
(Populus/Salix, probably
Populus sp.)
Feature 4 top of backfill of cremation pit, H, 80 cm BS
NA 9920±80 11,705-11,201
Beta-280585 Pit charcoal (Populus balsamifera)
Feature 5 cremation pit, H, 85 cm BS
-24.0 9990±50 11,705-11,261
Beta-280586 Pit charcoal (Populus balsamifera)
Feature 5 cremation pit, H, 85 cm BS
-23.4 10020±50 11,753-11,261
Dates associated with Component 2
Beta-286266 charcoal (Populus sp.) Paleosol 1b, Main, 135 cm BS
-22.6 9220±40 10,500-10,257
Beta-280914 hearth charcoal (Amelanchier sp.)
Feature 6 hearth, Main, 135 cm BS
-25.0 10090±50 11,966-11,397
Beta-286267 hearth charcoal (possibly
Populus sp.)
Feature 6 hearth, Main, 135 cm BS
-24.8 10180±50 12,063-11,630
Beta-286268 charcoal (indeterminate) (split 2)
Paleosol 1b, H, 115 cm BS
-25.1 9310±40 10,653-10,303
Beta-288416 charcoal (indeterminate) (split 2)
Paleosol 1b, H, 115 cm BS
NA 9720±50 11,242-10,830
Beta-288418 alkali soluble portion of Beta-288416
Paleosol 1b, H, 115 cm BS
NA 9290±50 10,647-10,285
Beta-232393 charcoal (Salix sp.) Paleosol 1b, Main, 135 cm BS
-22.6 9670±40 11,204-10,796
Dates associated with Component 1
AA-76863 hearth charcoal (Salix sp.) (split 1)
Feature 2 hearth, Main, 230 cm BS
-26.8 11250±60 13,297-12,954
Beta-232394 hearth charcoal (Salix sp.) (split 1)
Feature 2 hearth, Main, 230 cm BS
-26.9 11300±40 13,294-13,105
Beta-233316 hearth charcoal (Salix sp.) Feature 2 hearth, Main, 230 cm S
8
Figure Captions
Fig. S1. Generalized stratigraphic profiles for the Main Block and Cremation Area (Block
H). Note: Paleosols P1a, P1b, and P2 are nearly continuous between the Main Block and
cremation area.
Fig. S2. Photograph of the burial pit and north wall stratigraphy during excavation. At
this point, excavations have gone through the house floor to expose the pit/hearth in cross
section and plan views. A single thin charcoal-stained surface extended from the edges of
the pit/hearth to the north and west walls, marked ‘house floor’ and ‘house edge’.
Fig. S3. Posterior occipital fragment exhibiting differential burning indicative of body
position in the cremation fire (see text for details).
Fig. S4. Unerupted permanent lower lateral incisors, canines, first premolars, and left first
molar. Differential burning is evident by variation in color.