President’s Remarks
SRM began in 1948 with the objectives of enhancing the profes- sionalism of its members on the assumption that members’contri- butions to SRM made it strong and that they in turn received increased stature through improving credibility of the profession. However, when most range managers went to the public arena, it was more like “an unwilling friend at court.”
SRM has proposed the same objectives in the last few years and emphasized a new one, which asked for creation of public appreci- ation for range environments. This new objective requires that range professionals engage in public affairs with energy and imagination.
Another objective must now be considered, to develop public acceptance of the Range Management profession. Unless the pub- lic perceives a need for range managers, there will be no range profession. Range managers and SRM have worked long and hard for public benefits. I believe, as many of you do, that most range- lands are in better condition today than they were fifty years ago. But full public recognition of this success is lacking. Herein lies a major unfilled responsibility of SRM and the profession.
The willingness of individual members to engage in public advo- cacy and to permit SRM to represent them is gradually increasing. In recent years we have served as sources of expertise on controver- sial public policy issues. More and more the range scientists evalu- ate the potential uses and consequences of their findings. Range managers serve as advisors and consultants to elected policy mak- ers, as expert witnesses in the courts, and some are high level administrators. Some of us are social critics and advocates before and for community groups. These newly emerging professional roles present SRM with opportunities for public service that must not be lost.
Your officers for 1980 have seen fit to continue dealing with public policy issues and a start has been made toward re- examination of SRM’s total program. There is still much to do to promote sensitivity and understanding of issues that touch us.
My final report as president would be incomplete without a summary of SRM’s financial status. It was a traumatic few hours before and during the business meeting last year when we had to tell you that cash on hand would cover two months of operating expenses but it would be eight months before dues would be paid again. The donation campaign, which began a year ago, yielded over $27,000 from 500 members and started SRM’s comeback to financial solvency. A number of other opportunities were taken to increase income including higher sale prices of publications, dou- bling of our advertising rates, higher subscription prices, the unpopular dues increase, collection of fees and back rent owed us, closing of the books on the Old West grant, and an insurance settlement from a flood (broken pipe) that damaged floors, walls, doors, and publications in inventory. The Colorado and Wyoming Sections contributed the labor for repair after the flood.
Expenses were reduced for travel, by bulk purchase of office supplies, by postponing publication of a book, by reducing heat and light in the building, by not financing committee expenses as should normally be done, and by a host of other small actions. The
Executive Secretary donated considerable building upkeep on his own time.
The most visible changes of all have been improvements in office efficiency with new machines and procedures. A new typesetter was purchased at less monthly payment than annual costs of payments and frequent repairs on the old machine. The elimina- tion of down-time and about one-third faster operations has allowed a full-length book to be set while the publication schedule for JRM and Rangelands has been improved. A new copy machine decreased costs per page from 4.5 cents to 0.7 cents. That saving nearly covers installment payments on the new copier. The mem- bership records have been-automated including a new membership form, which has increased the capacity for retrieval of membershin information. The manually-operated 7,000 cardex card file was eliminated thereby eliminating a half-time job.
The accounting system has been completely revised and auto- mated into the computer, where accounts and membership are now inter-connected. The computer program gives printouts of costs and incomes for all SRM functions. Summary accounts of cash flow on a monthly basis allow detection of financial problems at a moment’s notice.
Budgets, balance sheets, and accounts summaries will appear in Rangelands for a permanent record for all to see. However, here are some highlights of SRM’s financial position at the moment. The 198 1 budgetshows estimated expenses at $275,186 and income at $3 10,598. &rr financial position on February 1 was as follows:
Renner fund $12,010
Building Trust fund 15,733
Life membership fund 26,128
Memorial fund 988
Savings 75,000
Cash on hand 21,641
$151,500 Of the total, $130,000 are in short term CD’s earning interest between 14 and 15.5%. A year ago SRM had $22,000 in its check- ing account and $11,000 in the Renner Fund. Thus, a step has been made to re-establish funds to meet building debts and to service life memberships.
SRM did not borrow from the bank to cover 1980 operations but we did borrow from the 1981 dues beginning in October. The goal in the 1981 budget is not to borrow from 1982 income. SRM’s financial situation has greatly improved. The membership, the committees, the sections-and the officers have made possible this financial turnaround and I personally thank everyone-in SRM for making this effort so successful. _
Finally, I say with all sincerety the SRM’s most valuable asset is the Denver office staff, consisting of Pat Smith and Pat Willems in the Publications Department, Candy Pacheco as Administrative Assistant, Carolyn Moore who is Records Supervisor, Mary Ann Nearing, Staff Secretary, and of course Floyd Kinsinger who makes all of us justify every cent in our expense accounts. These people in their dedication and high quality
put SRM on its feet than any others.
work have done more to
This address was given February I I, 1981, in Tulsa, Oklahoma, at the 34th Annual Meeting of the Society for Range Management.
JOURNAL OF RANGE MANAGEMENT 34(2), March 1981
Harold F. Heady, President, 1980
Temperature Profiles for Germination of Blue-
bunch and Beardless Wheatgrasses
J.A. YOUNG, R.E. ECKERT, JR., AND R.A. EVANS
Abstract
The germination of seeds of beardless and bluebunch wheat- grasses was investigated over a wide range of constant and alternat- ing temperatures. Seeds of ‘Whitmar’ beardless wheatgrass, a collection from Nevada, and numbered accessions of bluebunch wheatgrass were used. Seeds of ‘Whitmar’ beardless wheatgrass germinated at 87% of the 55 temperature regimes tested with a mean germination of 52%. Germination at 42% of the temperature regimes was optimum [defined as not significantly (P = 0.01) different from maximum], with a mean of 84%. Freshly harvested seeds of the Nevada source of bluebunch wheatgrass germinated at 78% of the temperature regimes with a mean of 40%. Comparable figures for fully ripened seeds 5 months after harvest were 84% with a mean of 62%. The germination response of l-month old blue- bunch wheatgrass seeds indicated that germination could occur at the high seedbed temperatures encountered in a late summer mois- ture event. The temperature-germination profiles for the num- bered accessions of bluebunch wheatgrass had the same general pattern as the Nevada source. Generally, seeds were highly germi- nable at a number of temperatures. Optimum germination of all the sources of seed occurred at 37 temperature regimes at least once and always occurred at 15 temperatures ranging from an alternat- ing 5/15’ C through a constant 25” C. This range of germination temperatures is much wider than that exhibited by squirreltail and Sandberg bluebunch. The bluebunch wheatgrass material has the inherent potential to germinate and to be highly germinable at a wide range of temperatures.
Bluebunch wheatgrass (Agropyron spicatum) is one of the most widespread and important native forage grasses on rangelands of the western United States (Hitchcock 1951). Bluebunch wheat- grass is the dominant or codominant perennial grass of sagebrush (Artemisia)/ bunchgrass communities in eastern Washington, Idaho, eastern Oregon, northern Nevada, and Utah (Stoddard 194 1, Daubenmire 1970).
Considering its wide distribution and the characteristics of other species of Agropyron, bluebunch wheatgrass is not a highly varia- ble species (Hitchcock et al. 1969). Generally, awned and awnless forms are recognized. The awnless phase has been generally recog- nized in the commercial seed trade and range literature as beardless wheatgrass (Agropyron inerme). Cytogentic evidence shows the two phases to be completely interfertile with normal pairing in meiosis (Cronquist et al. 1977). Understanding the germination ecology of bluebunch and beardless wheatgrasses is vitally impor- tant for cultivar selection, management of native stands, and artifi- cial revegetation of degraded stands. One aspect of the germination of these wheatgrass species is their response to temperature. Our purpose was to evaluate the germination of a broad spectrum of wheatgrass collections in response to constant and alternating incubation temperatures.
The authors are range scientists, U.S. Dep. Agr., Sci. and Educ. Admin., Agr. Res., Renewable Resource Center, 920 Valley Road. Univ. of Nevada, Reno, 89512.
This research is the result of cooperative investigations of the U.S. Dep. Agr., Sci. and Educ. Admin.-Agr. Res. and the Agr. Exp. Sta., Univ. of Nevada, Reno, Journal Series No. 444.
a4
Materials and Methods
We used seeds of beardless wheatgrass, cultivar ‘Whitmar’ obtained from the Soil Conservation Service (SCS) U.S. Depart- ment of Agriculture (Table 1). A total of 10 sources of bluebunch wheatgrass were tested, nine from the SCS, and one from Nevada. The material obtained from the SCS represents the accessions in their collections under evaluation for selection of a cultivar. Acces- sion P-285272 is considered a hybrid of beardless and bluebunch wheatgrass. The source from Nevada was collected from a native stand on the Goldbanks allotment south of Winnemucca, Hum- boldt County, Nevada.
The accessions obtained from the SCS differed in the location at which they were grown, Pullman, Washington, or Bridger, Mon- tana; and the year grown, 1968 to 1976 (Table 1). All SCS acces- sions were tested in 1978.
The Nevada material was grown in a space planted common garden at Reno, Nevada. The garden was given weed control and supplemental irrigation. It was established with seedlings trans- planted from the greenhouse in 1977. A limited number of seeds were collected from the plants in 1977 but seed production was excellent in 1978.
All germination experiments involved four replications of 25 seeds each and tests were repeated at least once. Seeds were placed on a single thickness of germination paper in petri dishes and kept moist with tap water. Tests were conducted in the dark. Seeds were incubated for 4 weeks with germination counts made weekly. Seeds from all sources were initially tested at constant incubation temperatures of 2,5, 10, 15,20,25, and 30” C. Based on the results of these initial experiments, we selected ‘Whitmar’, the Nevada material, P-6409, P-737, P-739, P-285272, and M-883 for addi- tional testing to develop complete germination profiles.
For development of germination profiles, seeds of the selected sources were incubated at constant temperatures of 0,2,5, 10, 15, 20, 25, 30, 35, and 40” C. Alternating temperature regimes con- sisted of 16 hours at each lower constant temperature and 8 hours at all possible higher temperatures in each 24-hour period. For example, 0” C was alternated with 2, 5, 10, 15, 20, 25, 30,35, and 40” C whereas 35” C was alternated with 40” C only. In all, 55 constant and alternating temperature regimes were tested.
In 1978 we tested the Nevada source seed 1 month after harvest and again 5 months after harvest to test for afterripening requirements.
After germination data were obtained for all 55 temperature regimes, the percentage of temperature regimes and mean germina- tion for those temperatures that produced some germination were obtained. We define optimum germination percentages as those not significantly different (P = 0.01) from the maximum. The number and mean of the optimums were calculated for each pro- file. The percent of temperature regimes having germination in the percentiles < 10, 10 to 25 (not including 25), 25 to 50,50 to 75,75 to 90 and >90% was calculated for each germination profile.
We compared the temperature-germination profiles of the selec- tions tested under a descriminate classification of seedbed temper-
Table 1. Species, accession, cultivar, or collection and origin of seeds of wheatgrasses used in germination experiments.’
Species or forma
Accession, cultivar, or collection Agropyron inerme
Agropyron spicatum Agropyron spicatum Agropyron spicatum Agropyron spicatum A&-obyron spicatum Agropyron spicatum X
A. inerme Agropyron spicatum Agropyron spicatum Agropyron spicatum Agropyron spicatum
‘Whitmar’ P-6409 P-737 P-739 P- 15623 P- 15634 P-285272 M-883 C-29 Nevada P- 15649
Origin
Union County, Oreg. Lewiston, Idaho Joel, Idaho Asotin, Wash. Granite County, Mont. Park County, Wyo.
Tag Germination germination profiles
Year grown Location grown (%) developed
76 Union, Oreg. 81 X
76 Pullman, Wash. 81 X
75 Pullman, Wash. 80 X
75 Pullman, Wash. 86 X
68 Bridger, Mont. 67
68 Bridger, Mont. 42
Whitman County, Wash. Wheatland County, Mont. Grand County, Colo. Humboldt County, Nev. Lincoln County, Mont.
71 Bridger, Mont. - X
74 Bridger, Mont. - X
70 Bridger, Mont. -
77-78 Reno, Nev. - X
75 Bridger, Mont. -
‘Year grown and tag germination taken from seed tag. Not all lots had germination indicated.
atures. This classification system included 19 moderate temperature regimes, 5/ 15, (5O C for 16 hr and 15’ C for 8 hr in each day) 5/20, 5/25, lo/ 10 through 10/30, IS/ 15 through 15/35, 20/20 through 20/35,25/ 25/ and 25/ 30” C; 1 I colder than moder- ate regimes, O/O through O/ 15,2/2 through 2/ 15,5/5and 5/ 100 C;
16 widely fluctuating regimes, O/20 through O/40, 2/20 through 2/40, 5/30 through 5/40, lo/35 and 10/40, and 15/400 C; and 9 warmer-than-moderate regimes 20/ 40, 25/ 35, 25/ 40, 30/ 30 through 30/40, 35/35, 35/40, and 40/400 C. This classification is based on the monitoring of field seedbeds in sagebrush grassland (Evans et al. 1970) and experience with the germination response of many grass species.
fluctuating temperature regimes of O/40, 2/40, and 5/40“ C, and the very warm incubation temperature of 40” C.
A frequency analysis was used to determine, for all sources of bluebunch wheatgrass, which temperature regimes supported ger- mination most frequently in the optimum range.
The inherent ability to germinate at low temperatures while soil moisture is still available is one of the most important germination characteristics influencing seedling establishment in the spring on sagebrush rangelands (Evans et al. 1970). Differences of a few days may mean the difference between successful establishment and seeding failure. ‘Whitmar’ seeds germinated after 4 weeks incuba- tion at O/20” C at a level significantly (P = 0.01) lower from the optimum. However, even this is a fairly high day temperature for field seedbeds in the early spring (Evans et al. 1970). A O/ 15” C temperature regime probably better reflects field conditions. At this regime ‘Whitmar’seed germination fell to 32% and at O/ lo” C it was only 3% (Table 3). Raising the cold period temperature to 2’ C did not significantly increase the germination with warm-period temperatures 15, 20, or 25’ C.
Results Screening of Seed from All Sources
Comparison of the germination at constant temperatures of bluebunch and beardless wheatgrass seeds from the various sour- ces revealed three lots, P-15634, C-29, and P-15623, that were significantly less germinable than the rest (Table 2). These three accessions were the oldest seeds tested and apparently had largely lost their germinability; two accessions had previously much higher germination percentages, indicated by their lot tags (Table
I). These three accessions were dropped from further testing.
With cold-period temperatures from 5 through 25” C and warm- period temperatures below 35” C, the germination was in the optimum range (Table 3). Germination at IS/ 25” C, the tempera- ture regime used for official determinations of the germination of this cultivar (Anon. 1978), was 95%, the maximum obtained. Bluebunch Wheatgrass
After-ripening
‘Whitmar’ Beardless Wheatgrass
The seeds of many species require a period of time after they fall from the maternal plant for embryo maturation before they will germinate. This time period is the afterripening requirement (Mayer and Poljakoff-Mayher 1963).
Some germination occurred at 87% of the temperature regimes When seeds of bluebunch wheatgrass from the Nevada 1978 tested with a mean germination of 52% (Table 3). Germination at source were tested 1 month after harvest, germination occurred at 42% of the 55 temperature regimes used was in the optimum range 78% of the 55 regimes tested, with a mean of 40% (Table 4). with a mean of optimums of 84%. Germination was completely Optimum germination occurred at 18% of the temperature regimes inhibited only at the very cold temperatures of 0” C, O/2 (00 C for with a mean of 83%. Two groups of temperature regimes supported 16 hr alternating with 2’ C for 8 hr) and O/So C, the extremely optimum germination; one was at 10/35O C, 20/200 C and all
Table 2. Percent germination at constant temperatures for bluebunch and beardless wheatgrass seed from 12 sources.
_ Germination (%)I
Temperature Nevada
(co) P-737 M-883 P-6409 ‘Whitmar’ P-l 5634 C-29 P-285272 P- 15649 P- I5623 P-739 1977 1978
2 54 a-c 87 a-d 30 e 8b 0 Id 8c 48 bc 0 12e Od 10 b
5 50 a-d 72 bd 50 de 88 a 2 Id 22 c 25 c 0 26 de 58 b 96 a
10 65 a 90 a-c 76 a-c 96 a I5 Id 92 a 80 a 8d 70 ac 71 ab 88 a
15 57 a-c 91 a-c 67 a-d 89 a 16 12 cd 83 ab 73 ab 0 93 a 84 a 89 a
20 52 ad 93 ab 72 a-d 85 a 15 30 a 90 a 81 a 0 78 a-c 62 b 85 a
25 43 ae 84 ad 70 a-d 82 a 19 7 cd 83 ab 77 ab 0 78 a-c 23 c 82 a 30 28 d-f 80 ad 47 de 21 b 11 4d 69 b 63 ab 10 cd 60 a-c 8 cd 19 b
Mean 50 b 85 a 59 b 67 ab 11 c 8c 64 ab 64 ab 3c 60 ab 44 b 67 ab
‘Means within sources compared vertically within sources, and overall means compared horizontally among sources. Means followed by the same letter are not significantly different at the 0.01 level of probability as determined by Duncan’s multiple range test. No letters indicates no significance.
Table 3. Germination prafile for seeds of ‘Whitmar’ beardless wheatgrass.’
Cool-period temperature
(” C) (16 hr) 0
0 0
2
5
10
15
20
25
30 35 40
~~ ~~ Germination (%)
Warm-period temperature o C 8-hr
2 5 10 15 20 25 30 35 40
0 0 3j 32 f-j 45 d-g 74 a-d 22 f-j 1 j 0
3j 3j 12 hi-j 45 d-g 71 ad 68 a-d 19 f-j 5 ij 0
90 ab 90 ab 89 ab 85 ab 95 a 89 ab 39 e-h 0
72 ad 89 ab 93 a 83 ab 86 ab 45 d-g 3i
87 ab 95 a 95 a 76 abc -47 c-f 14 hij
81 ab 95 a 87 ab 25 f-j 14 hij
71 a-d 68 ad 23 f-j 16 g-j
61 b-e 35 e-i 18 f-j 22 f-j 3j
0 Some germination = 87% X germination = 52%.
Optimums = 42% 2 of optimums = 84%.
Germination percentiles
< 10=25%, 10 to 25= 16%, 25 to 50= 15%, 50 to 75= 13%, 76 to 90= 18%, >90= 13%
1 Means followed by the same letter are not significantly different at the 0.01 level of probability as determined by Duncan’s multiple range test. For the reader’s convenience, means not significantly different from optimum are underlined.
temperatures alternating with 15” C as the cool-period tempera- ture and the second at O/ 15 and O/ 20” C.
It is not unusual for freshly harvested seeds to have optimum germination at temperature regimes with 2 or 0” C cold periods. The cool period portion of the regime serves as a cool-moist stratification environment that satisfies afterripening require- ments. The germination profile for the freshly harvested Nevada source seed is unusual in that germination is greatly restricted at temperature regimes with 2’ C cold periods compared to 0 or 5” C (Table 4). Upon observing this unusual pattern we immediately repeated the germination test with 2-month-old seed. We obtained generally higher germination, but the depression with the 2” C cold periods and optimums with the O/ 15 and O/ 20” C regimes con- tinued (data not shown).
Since bluebunch wheatgrass seeds germinate very well at some temperatures 1 month after harvest, germination with unusual moisture events during late August or early September is possible. A 1 S/ 400 C temperature regime is conceivable for this time period, and seeds could germinate if the moisture event was of sufficient duration and magnitude. Harris (1957) considered the establish- ment of spring germinated bluebunch wheatgrass to be condi- tioned by the occurrence of unusual moisture events during the usually dry summer periods characteristic of the far west. Fully Ripened Germination
Five months after harvest there was no additional increase in germinability. The 5-month post-harvest profile had some germi- nation at 84% of the temperature regimes tested, with a mean of
Table 4. Germination profile of bluebunch wheatgrass from Nevada source grown in a common garden at Reno in 1978. Germination test conducted 1 month after harvest.1
Cool-period Germination (%)
temperature
(“C) Warm-period temperature o C 8-hr
(16 hrj 0 2 5 10 15 20 25 30 35 40
0 0
2 5 10
15
20
25 30 35 40
0
0
3 up 57 c-g 70 a-e 74 a-e 55 d-h 11 k-p 1P 0
0 0 7 m-p 2 oP 1P 0 0 0
32 g-m 40 f-j 41 f-j 28 i-o 36 g-k 34 g-l 12 k-p 17 j-p 32 g-m 41 f-j 40 f-j 50 e-i 36 g-k 80 ad 66 b-f
82 abc 87 ab 87 ab 89 ab 86 ab 77 a-d _ 94 a 30 h-m 37 g-k 29 h-n 25 i-p
24 i-p 28 i-o 29 h-n 25 i-p 14 j-p 8 l-p 6 m-p
0 0
0
Some germination = 78% ‘; germination = 40%.
Optimums = 18% x’ of optimums = 83%.
Germination oercentiles
< 10=35%, 10 to 25= 1 l%, 25 to 50=29%, 50 to 75= 1 l%, 75 to 90= 13%, >90= 1%.
1 Means followed by the same letter are not significantly different at the 0.01 level of probability as determined by Duncan’s multiple range test. For the reader’s convenience, means not significantly different from maximum are underlined.
Table 5. Germination profile of seeds of bluebunch wheatgrass from the Nevada source, grown in a common garden. Germination test conducted 5 months after harvest.1
Cool-period Germination (%)
temperature
(“C) Warm-period temperature o C 8-hr
(16 hr) 0 2 5 10 15 20 25 30 35 40
0 0 0 0 4i 10 ghi 9-ghi 47 d-f 14 ghi 14 ghi 0
2 10 ghi 4i 34 e-h 98 a 95 a 87 abc 77 ac 0 0
5 96 a 95 a 94 ab 94 ab 97 a 58 c-e 77 ghi 0
10 88 ab 81 abc 89 ab 93 ab 87 ab 73 ad 65 bd
15 89 ab 94 ab 94 ab 94 ab 82 abc 72 a-d
20 85 abc 93 ab 92 ab 89 ab 93 ab
25 82 abc 85 abc 10 ghi 6 hi
30 19 ghi 22 f-i 5i
35 4i 0
40 0
Some germination = 84% x germination = 62%.
Optimums = 51% x of optimums = 89%.
Germination percentiles
<10=27%, 10 to 25=13%, 25 to 50=5%, 50 to 75=7%, 75 to 90=22%, >90=26%
‘Means followed by the same letter are not significantly different at the 0.01 level of probability as determined by Duncan’s multiple range test. For the reader’s convenience, means not significantly different from maximum are underlined.
62% (Table 5). Optimum germination occurred at 51% of the temperature regimes, with a mean of 89%.
The depression in germination with 2” C cold periods was no longer apparent. However, germination at O/ 10, O/ 15, and O/ 20” C was markedly lower for the fully matured seeds than for seeds tested l-month post-harvest.
sion P-6409 had greater than 50% germination at 73% of the temperature regimes tested although 90% germination was attained at only 6% of the regimes tested. However, there was no significant (P = 0.01) difference among the accessions in the per- centage of temperature regimes producing germination equal to or greater than 90%.
The seeds of the Nevada source of bluebunch wheatgrass must be classed as highly germinable, for 25% of the temperature regimes supported germination at or above 90% and 47% sup- ported germination at 75% or above (Table 5).
Optimum Germination
This germination profile is an indication of the potential of seeds of the species to germinate. Germination and seed production of many native grasses, including beardless wheatgrass, have been shown to increase under an optimum seed-production environ- ment (Miltimore et al. 1962). Seeds of bluebunch wheatgrass pro- duced from native stands under harsh environmental conditions probably would be much less germinable. Seeds of squirreltail
(Sitanion hystrix) from native stands have been shown to be more highly germinable, with optimum germination at a greater number of temperatures, in favorable seasons for growth and seed produc- tion (Young and Evans 1977).
We found 15 temperature regimes that always supported opti- mum germination for the accessions tested (Table 7). These temperatures were in a block of 5/ 15 through 5/30” C, lo/ 15 through 15/30” C, and 20/20 through 20/ 30 and 25/25’ C. Com- parable figures on the temperature regimes that support optimum germination for squirreltail and Sandberg bluegrass (Poa Sand- bergii) seeds collected from native stands reveal only three temper- ature regimes for each species that were always optimum (Evans et
Table 6. Selected characteristics of germination-temperature profiles for five numbered accessions of bluebunch wheatgrass seed tested under 55 temperature regimes.
Comparison of Numbered Accessions
Temperature profile Accessions
characteristics’ P-737 P-285272 P-6409 P-739 M-883 The numbered accessions represent the plant material from
which a cultivar of bluebunch wheatgrass may be selected for release for use on rangelands. Over a longer time, plant breeders may recombine this and other plant material through hybridiza- tion and selection to develop additional cultivars. The numbered accessions were originally collected over a wide geographic area of the western United States and provide a broad representation of the species.
Some germination
(%) 78 b 90 a 82 b 82 b 93 a Mean germination
(%) 35 c 49 bc 70 a 64 ab 59 ab Optimum
germination (%) 38 b 38 b 56 a 51 a 51 a Mean of optimums
(%) 55 b 66 ab 81 a 81 a 82 a
Accession M-883, originally collected in Wheatland County, Montana, had optimum germination in four categories: some germination, mean germination, optimums, and mean of opti- mums (Table 6). Both accessions P-6409 and P-739 were optimum in all categories except for the some germination category. The hybrid accession P-285272 had some germination at 90% of the 55 temperature regimes tested.
Percentiles <lO%
10 to 25% 25 to 50% 50 to 75% 75 to 90% >90%
33 a W 10 CY 4dZ 25bW 16cXY 13abY 11 bY 20 a Y 3cY 7 bc Z 25 b X 32 a X 4dZ 11 cx 15 c XY 29bWX40aW 27bX 24 b W 22 b X
ocz 7cY 40 a W 27 b W 29 b W
oz OZ 6Z 10 x 11 YZ
Germination Percentiles
Comparing germination by percentiles revealed accession P- 6409 had less than 10% germination at only 4% of the temperature regimes tested (Table 6). In comparison, accession P-737 had less than 10% germination at 33% of the temperature regimes. Acces-
1 Means of temperature-profile characteristics and percentiles are compared horizontally among accessions. Means followed by the same letter (II through d) are not significantly different at the 0.01 level of probability as determined by Dunan’s multiple range test. No letters indicates no significance. Means of percentiles are compared vertically within accessions and percentile means followed by the same letter (w through z) are not significantly different at the 0.01 level of probability.
Table 7. Frequency ($) that optimum germination occurred at a given temperature regime in the germination profiles of five numbered accessions and the Nevada collection of bluebunch wheatgrass.’
Cool-period temperature e C)
(16 hr) 0 0 2 5 10 15 20 25
2 10
17 68 85
Freqtiency of optimum germination (%) Warm-period temperature o C 8-hr
15 20 25
85 85 85
100 100 100
100 85 100
100 100 100
100 100
100
30 35 40
68 17
100 51
100 68
100 68 51
100 68 51
85 17 34
30 17 34 17
35 40
‘For emphasis, the 100% frequencies are underlined.
al. 1977; Young and Evans 1977). These temperatures were lo/ 15, lo/ 20, and 15115’ C for squirreltail and 5/ 20, lo/ 20, and 1 S/ 200 C for Sandberg bluegrass.
For the bluebunch wheatgrass complex, 37 temperature regimes supported optimum germination at least once (Table 7). In com- parison, only 26 temperature regimes for squirreltail (Young and Evans 1977) and 17 for Sandberg bluegrass supported optimum germination at least once (Evans et al. 1977). However, the wheat- grass seeds were harvested from plants grown in gardens and the squirreltail and Sandberg bluegrass seeds were collected from native stands.
Constant vs. Alternating Temperatures
Germination in the field is almost always under alternating temperature regimes, but it is very convenient to do germination tests at one constant temperature. In the field the amount of diurnal fluctuation in seedbed temperature can be modified by the depth of planting or through the use of furrows (Evans and Young 1972). The germination of the seeds of some species is markedly enhanced by alternating temperatures. Mayer and Poljakoff- Mayher ( 1963) and Plummer ( 1943) reported that bluebunch wheatgrass seeds germinated better under alternating than under constant temperatures.
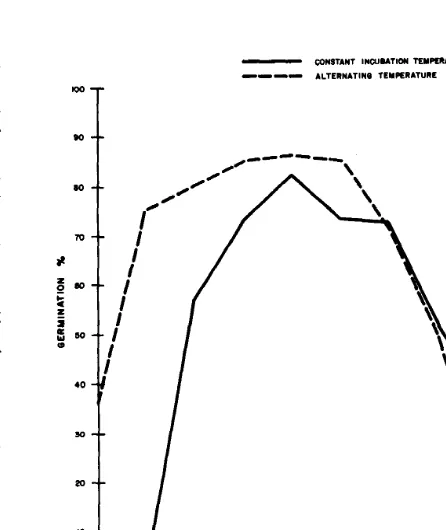
We found no readily apparent advantage to alternating versus constant temperature. For a more detailed comparison, we plotted the mean germination of all sources tested at constant temperature and the mean germination at alternating temperature with the base temperature with the highest mean germination (Fig. 1). In this comparison the alternating temperature regimes produced greater germination at low temperature, 0 through lo” C; roughly equal germination at moderate temperatures, 15 and 30" C; and lower germination at 35” C.
We compared the germination at constant temperatures with alternating temperature regimes that had the same number of
0 2 s IO II LO es w %
INCUBATION TEMPERATURE
Fig. 1. Comparison of germination at constant incubation temperatures
and at the alternating regimes with the base temperature that had the highest mean germination. Means of jive numbered accessions, the Nevada collection of bluebunch wheatgrass, and ‘Whitmar’ beardless wheatgrass.
Table 8. Mean germination and percentages of temperature regimes with some germination at moderate and extreme (colder, widely fluctuating, and warmer) temperature regimes.’
Mean germination (o/O) at indicated regime Regimes with some germination (%)
Widely Widely
Wheatgrass Moderate Colder fluctuating Warmer Moderate Colder fluctuating Warmer
‘Whitmar’ beardless 80 bc 25 jk 31 i 21 km 100 a 73 ef 81 c 89 b
Nevada bluebunch 90 a 32 i 40 h 18 mn 100 a 73 ef 75 de 78 cd
M-883 bluebunch 85 ab 23 km 48 fg 43 gh 100 a 64 h 100 a 100 a
P-737 bluebunch 52 ef 13 n 18 mn 18 mn 100 a 46 j 81 c 67 gh
P-6409 bluebunch 83 bc 30 ij 45 gh 56 e 100 a 75 de 69 fg 89 b
P-739 bluebunch 78 c 33 i 39 h 52 ef 100 a 46 j 81 c 89 b
P-285272 bluebunch 66 d 15 n 42 h 44 gh 100 a 55 i 100 a 100 a
1 Means followed by the same letter are not significantly different at the 0.0 1 level of significance as determined by Duncan’s multiple range test. Mean germination and percent with some germination compared separately.
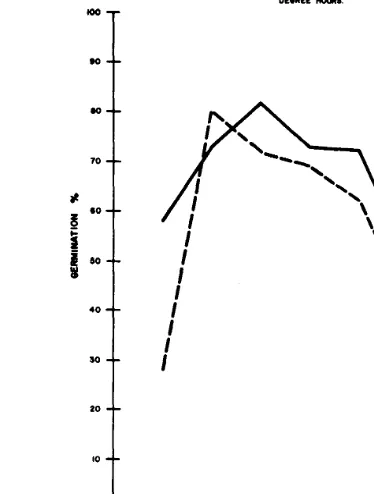
degree hours above 00 C in equivalent time periods (Fig. 2). In this comparison, there appears to be an advantage to constant temper- ature incubation at 5” C, probably because the equivalent alternat- ing regimes alternate with 0“ C.
Comparison with Seedbed Temperatures
At what we classify as moderate seedbed temperatures the Nev- ada source of bluebunch wheatgrass had the highest germination (Table 8), but was not significantly (P = 0.01) higher in germina- tion than accession M-883. The Nevada collection and accessions P-6409 and P-739 of bluebunch wheatgrass had the highest germi- nation at colder than moderate seedbed temperatures.
The range between highest and lowest mean germination drops dramatically with the colder than moderate temperature regimes, 38 versus 20 percent (Table 8). Accession P-285272 of bluebunch wheatgrass had the next to lowest mean germination at moderate and colder than moderate temperatures, but was among the high- est germinators at widely fluctuating and warmer temperatures. There appears to be a correlation between germination at warmer than moderate temperatures and widely fluctuating temperatures. Such a relationship would indicate that the limiting factor in the widely fluctuating temperature regimes may be the warmer temperatures.
All of the sources tested generally had some germination at a high percentage of the temperature regimes tested with the excep- tion of the colder than moderate classification. However, acces- sions M-883 and P-285272 were exceptional in germinating at all of the widely fluctuating and warmer regimes. No matter how limited, germination at extreme temperatures may be of practical importance. Wheatgrasses are normally seeded on sagebrush ran- gelands at the rate of 8 seeds per dm of row, and a fully stocked stand is one established plant per 3 dm of row. Theoretically, a fully stocked stand could be obtained with 4% germination, Obviously many more factors than germination enter into successful seedlings establishment, but a slight amount of germination at extra moder- ate temperatures might produce a successful stand.
Significance of Study
Many factors other than the germination response to tempera- ture enter into the selection of a cultivar of bluebunch wheatgrass for use on rangelands. This study shows that most of the bluebunch wheatgrass accessions evaluated have the inherent potential for germination over a wide range of temperatures.
Literature Cited
Anon. 1978. Rules for testing seeds. Assoc. of Official Seed Analysts. Vol. 3 (3):36.
Cronquist, Arthur, A.H. Holmgren, N.H. Holmgren, J.L. Reveal, and P.K. Holmgren. 1977. Intermountain Flora, Vol. 6. The Monocotyled- ons. Columbia Univ. Press, New York.
Daubenmire, R. 1970. Steppe vegetation of Washington. Washington Agr, Exp. Sta. Tech. Bull. 62. 131 p.
Evans, R.A., H.R. Holbo, R.E. Eckert, Jr., and J.A. Young. 1970. Func- tional environment of downy brome communities in relation to weed control and revegetation. Weed Sci. 18: 154-162.
70 E 4 00
P
1
SO.O
20
IO
-m-B
ooMsl4llT **uInon TE-lum? 4Ll2ml4n~ lmCa4nJM wlm 2ouI”4L2ln 02042E noun2
Fig. 2. Comparison of germination at constant incubation temperatures and at alternating temperatures regimes with equivalent amounts of degree hours above 00 C. Means offive numbered accessions, the Nevada collection of bluebunch wheatgrass, and ‘ Whitmar’ beardless wheatgrass.
Evans, R.A., and J.A. Young. 1972. Microsite requirements for establish- ment of annual rangeland weeds. Weed Sci. 20:350-356.
Evans, R.A., J.A. Young, and B.A. Roundy. 1977. Seedbed requirements for germination of Sandberg bluegrass. Agron. Jour. 60:8 17-820. Harris, G.A. 1967. Some competitive relationships between Agropyron
spicatum and Bromus tectorum. Ecol. Monogr. 37:89-l 11.
Hitchcock, A.S. 1951. Manual of the grasses of the United States. 2nd edition revised by Agnes Chase. Misc. Pub. No. 200. U.S. Government Printing Office. Washington, D.C.
Hitchcock, G.L., A. Cronquist, M. Ownby, and J.W. Thompson. 1969. Vascular plants of the Pacific Northwest. Part I. Vascular cryptogams, gymnosperms, and monocotyledons. Univ. of Washington Press, Seattle.
Mayer, A.M., and A. Poljakoff-Mayher. 1963. The germination of seeds. Pergamon Press, Oxford. 236 p.
Miltimore, J.E., J.L. Mason, and C.B.W. Rogers. 1962. Increase in seed production from nitrogen fertilization of native beardless wheatgrass. Can. J. Plant Sci. 42:359-364.
Plummer, A.P. 1943. The germination and early seedling development of twelve range grasses. J. Agron. 35:19-34.
Stoddard, L.A. 1941. The palouse grassland association in northern Utah. Ecology 22: 15% 163.
Young, J.A., and R.A. Evans. 1977. Squirreltail seed germination. J. Range Manage. 30:33-36.
A Versatile Flail-Type Forage Plot Harvester
.I. STUBBENDIECK AND CHARLES R. FENSTER
Abstract
A flail-type plot harvester was developed using eommercinllJ manufactured components and P specitically designed and con. strutted forage collection box. This forage plot harvester has been used successfully on experimental rangeland and seeded forage plots. The equipment is more versatile and less expensive than commercially manufactured plot harvesters.
Forage research is dependent on the successful quantification of vegetative response. Several different characteristics have been evaluated to measure vegetative response. However, the most uni- versally accepted criterion is yield. Weight of berbage produced per unit area (yield) is one of the most important characteristics of range plants, and it is probably the best single measure of growth (Hanson 1950). Yield is also one of the most labor intensive and time consuming vegetative characteristics to quantify, particularly when the historical technique of hand clipping is employed. The recognized importance of yield data to forage research has emphasized the need for more efficient, economical methods of experimental plot harvestmg.
Brown (1954) and Mannetje (1978) have discussed the many techniques and types of plot harvesting equipment which include band and electric clippers, sickle and rotary mowers, and flail choppers. Each harvesting technique has its advantages and disad- vantages. Hand and electric clippers are relatively inexpensive, but plot size may be limited due to labor requirements. Numerous researchers have used rotary mowers, but many found that tall plant material was difficult to harvest (Howell 1956; Fortmann 1956; McGinnies 1959). Several self propelled flail-type forage harvesters have been developed and tested (Kemp and Kalbfleisch
1957; Hubbard and Willis 1962; Thompson and Heinrichs 1963 Buker 1967; Swallow ,967; Allen et al. 1968; Collins et al. 1969) Flail-type plot harvesters were more labor efficient, but they were expensive and could be used for only one purpose, because the mower was permanently attached to the tractor, allowing no versa- tility in tractor use. This paper describes a rather simple, efficient, relatively economical, and versatile forage plot harvester that has been used successfully on experimental rangeland and seeded for- age plots.
Harvester Design and Operation
The harvester is a combination of three main components: (I) a garden tractor, (2) a commercially manufactured front-mounted flail mower, and (3) a specifically designed and custom constructed forage collection box. Garden tractors in a 12 to 16 b.p. range with both standard and hydrostatic transmissions were evaluated. The larger tractor (John Deere 316)’ with the hydrostatic transmission has been the most satisfactory combination. The hydrostatictrans- mission eliminates the problem of clutch slippage at reduced speed. The Haban 5 121 flail mower has a cutting width of 91 cm. It is belt driven by an electric p.t.a. at 1,800 to 2,200 rpm. Normal revolu- AUthorr ax aSI0Ci.a protcssor Of Agronomy (Range Management,, ““wernity of Nebraska. Llnco,” 6RSRL and profesiar
of
agronomy.
U”iYerSityof
Nebraska Pan- handle Station, Scotlrbluff 69361.tion direction of the rotor was reversed (factory option). This causes the harvested forage to be moved forward and lifted toward the top of the mower. The mower must be equipped with a remova- ble top cover (factory option). The top cover is removed and a piece of sheet metal (9 X 96 cm) and a 900 bend is attached to the lower lip of the opening. This sheet metal deflector (Fig. IA) forces the harvested forage into the collection box.
The collection box is 132 cm wide and is constructed of angle iron and sheet metal. Its lid (Fig, IB) latches in a closed position and hinges back over the hood of the tractor in the open position. The ends of the collection box, as well as two areas in the lid, have inserts of hardware cloth (Fig, IQ which allows air to escape causing an even distribution of forage in the collection box.
Cutting height is set by raising or lowering the gauge wbeel(Fig. ID) by adding or removing spacers (Fig. IE) on the wheel bracket shaft. The flail roller (Fig. IF) levels the mower. The level may be altered by changing the flail roller adjustment (Fig. IG).
This harvester has been used on irrigated and dryland grasses, legumes, and grass-legume mixtures, as well as on rangeland. Cutting speed was approximately I mjsec. Cutting heights have varied from 2.5 to 15.0 cm. At the lower cutting heights, a trace of soil has been detected in the samples. At the higher cutting heights, a small portion of the plant material is deposited on the soil surface.
The collection box will hold up to I5 kg of harvested forage. This capacity makes it possible to harvest forage plots of sufficient sire, even those that are irrigated. Forage is removed from the collection box and weighed separately. Representative subsamples are rela- tively simple to obtain because harvested plant material is coarsely ground and mixed. Subsamples are dried to determine moisture ccmtent and may be ground and analyzed for various quality partimeters.
Advantages
factured flail-type harvesters. One advantage is lower cost. The cost of our harvester in 1978 was approximately $3,100: (1) 16 h.p. hydrostatic tractor, $2,350; (2) flail mower, $600; and (3) custom constructed collection box, $150. This compared to an approxi- mate cost of $7,000 for a commercial flail-type harvester.
The harvester is easily transported. It can be hauled in a ‘/2 T pickup. We constructed a tilt-bed trailer, but a snowmobile trailer would work equally well. Another advantage is versatility. The collection box can be easily removed and the mower top cover replaced in a few minutes. The machine may then be used for cutting plot alleys and other routine mowing. The entire flail mower assembly may also be removed from the garden tractor in approximately 15 minutes. The garden tractor may then be used as the power source for numerous small implements.
Literature Cited
Alien, R.J., T.W. Casseimann, and F.H. Thomas. 1968. An improved forage harvester for experimental plots. Agron. J. 60: 584-585. Brown, Dorothy. 1954. Methods of surveying and measuring vegetation.
Bull. 42. Commonwealth Agr. Bur. Hurley, Berkshire, England. 223 p.
Buker, R.J. 1967. Forage plot harvester. Agron. J. 59:203-204. Collins, K.L., C.L. Rhykerd, and C.H. Noiier. 1969. A self-propelled
experimental plot forage harvester. Agron. J. 61:338-339.
Fortmann, H.R. 1956. Harvester for experimental forage plots. Agron. J. 48:241-242.
Hanson, Herbert C. 1950. Ecology of the grassland II. Bot. Rev. 16:283- 360.
Howell, H.B. 1956. A new experimental plot harvester. Agron. J. 48:240- 241.
Hubbard, W.A., and T.G. Willis. 1962. Note on a self-propelled flail-type forage plot harvester. Canadian J. Plant Sci. 42:739-741.
Kemp, J.G., and William Kaibfleisch. 1957. A crop harvester for forage plots. Canadian J. Plant Sci. 37:418-422.
Mannetje, L. ‘t. 1978. Measurement of grassland vegetation and animal production. Bull. 52. Commonwealth Agri. Bur. Hurley, Berkshire, England. 260 p.
McGinnies, W.J. 1959. A rotary lawn mower for sampling range herbage. J. Range Manage. 12:203-204.
Swallow, Clarence. 1967. Self-propelled plot forage harvester. Agron. J. 59:609-610.
Response of
Muhlenbergia porteri
Scribn. to
Season of Defoliation
RICHARD F. MILLER AND GARY B. DONART
Abstract
Removing 65% of the leaf area of bush muhly in three consecu- tive years during the growing season reduced plant vigor regardless
if effective management is to be implemented. of season of clipping. Late or continuous season defoliation had
The purpose of this study was to evaluate the effect of season of the greatest impact on food reserves, production, crown diameter
defoliation on (1) total nonstructural carbohydrates, (2) number of elongated internodes, (3) production and (4) crown diameter. and number of stem internodes. Defoliation during the vegetative
stage had the least effect of the clipping treatments.
Bush muhly (Muhlenbergia porteri) is a perennial grass charac-
Study Area
terized by numerous wiry, much-branched culms, growing on dry The study area was located on the New Mexico State University mesas and rocky slopes throughout the Southwest (Gould 1951). College Ranch, 32 km north of Las Cruces in Dona Ana County, Historically forming extensive stands in parts of its range, bush New Mexico. The arid climate is characterized by low precipita- muhly now is generally restricted to growing under the protection tion, low humidity, high temperatures, high evaporation potential, of shrubs and sub-shrubs. This grass species has been reported as and periodic strong winds. Average annual rainfall is 22 cm. being highly palatable (Gardner 195 1). However, Welsh and Beck Approximately 55% of the annual precipitation falls during the (1976) observed that the palatability of the species was only aver- growing season from July through September (Agr. Res. Serv. age and that it was sensitive to grazing because of its branching and 1975). Most summer precipitation comes from high intensity con-
nodal disarticulation characteristics. vectional thunderstorms (Paulsen and Ares 1962). Annual rainfall
Bush muhly can become established in badly deteriorated sites is highly variable with drought commonly occurring throughout occupied primarily by creosotebush (Lurrea tridentata) where their the desert grasslands. Drought (75% or less of average rainfall) geographic ranges overlap. As bush muhly gains in stature, it can occurs 20 to 40% of the time (Campbell and Crafts 1939). Potential outcompete creosotebush, leading to a decrease in shrub vigor and, annual evaporation rate averages 229 cm (Herbel et al. 1974). The in numerous instances, mortality (Welsh and Beck 1976). The frost-free period lasts approximately 200 days with maximum air ability of bush muhly to revegetate these areas naturally makes it a temperatures reaching 36” C in June.
highly desirable plant to maintain or increase in abundance. We The growing season generally ranges between 90 and 100 days need more information on the growth characteristics of this species depending primarily on rainfall distribution. Mean air tempera-
tures range from 16’ C to 33” C during this period with approxi- Authors are assistant professor, Rangeland Resources Program, Oregon State
University, Corvallis; and professor, Department of Animal and Range Sciences, New
mately 4 cm of rain falling per month.
Mexico State University, Las Cruces, respectively. Miller was formerly graduate Precipitation during the growing season was 207% of the mean research assistant, Department of Animal and Range Sciences, New Mexico State in 1974, 126% of the mean in 1975 and 72% of the mean in 1976 University, Las Cruces. (Fig. 1). Production during the 1974 growing season was lush.
This was submitted as Journal Article 747, New Mexico Agricultural Experiment
Station, Las Cruces, New Mexico, 88003. It reports on work supported in part by However, in 1975 over half of the rainfall came at the end of the Bureau of Land Management Contract YA-512-CT6-205. , growing season, losing much of its effectiveness.
Manuscript received November 11, 1979. The study site was located within a 5.6-ha cattle exclosure at an
l Monthly precipitation
l 59-Year average precipitatjon
__-_-___---_-_-__---.
JA SO J JASO J J AS0
1974 1975 1976
Fig. 1. Monthly precipitation during the growing seasons of 1974. 1975, 2976 and a 59-year average.
elevation of 1,400 m. The site has been protected from cattle grazing since 1965. Prior to 1960 very little bush muhly was present (Welsh 1972). In 1960 bush muhly began to establish rapidly at the base of individual creosotebush plants. Soils were gravelly sandy loam varying from zero to 92-t cm in depth. Soil depth was restricted by a fractured calcium carbonate hardpan. Common subordinate plants are fluffgrass (Erioneuron puchellum), loco- weed (Astragalus sp.), broom snakeweed (Xanthocephalum saro- thrae), honey mesquite (Prosopis jultjlora), and tarbush (Flourensia cernua).
Methods
The effects of six different clipping treatments were evaluated during 1974, 1975, and 1976. The six treatments involved clipping at different phenological stages during the three-year period. The specific treatments were (1) vegetative, plants clipped only in the vegetative stage once each year during the three-year period; (2) VFS, plants clipped once in the vegetative stage in 1974, once during flowering in 1975, and once during leaf senescence in 1976; (3) VS V, plants clipped once in the vegetative state in 1974, once in leaf senescence in 1975, and once in the vegetative stage in 1976; (4) maturity, plants clipped once in leaf senescence and once during quiescence in 1974, 1975, and 1976; (5) continuous, plants clipped once in each of the vegetative, flowering, leaf senescence, and quiescence stages during each of the three years; and (6) control, nondefoliated plants. The four phenological stages designating the period of defoliation were the third to fourth leaf stage for vegeta- tive, dough stage for flowering, after seed ripening for leaf senes- cence, and chlorophyll absent from about 90% of the leaf tissue for quiescence. Defoliation for all treatments was accomplished by clipping approximately 65% on a weight basis of the current annual growth.
Phenological data were collected on 24 bush muhly plants per treatment during each defoliation period throughout the study. Phenological measurements were recorded on the same plants during the three-year period.
Six plants were excavated in each treatment during each of the four phenological stages seven days after defoliation, placed on ice in the field and brought into the laboratory, where they were dried at 68’ C for 48 hours. Total nonstructural carbohydrates were extracted from 0.5 g root sample in 0.8 N H&04 (Smith et al.
1964). All samples were refluxed on a crude fiber apparatus. An extension of Forsee’s photocolorimetric ferricyanide method was used for determining TNC levels (Morrell 1941).
Elongated internodes were counted on ten randomly selected culms per plant in each treatment prior to defoliation in 1975 and 1976. Eight observations were made per treatment each year.
Clipped material was collected and bagged for each plant during defoliation for dry matter yield determinations. Crown diameters were measured at the end of each growing season for all six treatments on bush muhly.
Analysis of variance, Tukey’s w-procedure and the pooled-t test were used to analyze data at the 95% confidence level (Steel and Torrie 1960).
Results and Discussion
Bush muhly initiated primary leaf growth during mid to late July in all three growing seasons, dependent upon rainfall (Table 1). In late May and early June of 1975 and 1976, limited moisture initiated leaf growth. However, bush muhly returned to a quiescent state in mid-June due to an absence of available soil water. Flower- ing occurred 34 and 29 days earlier in 1974 than in 1975 and 1976, respectively. This was due to limited rainfall during August in 1975 and 1976 (Fig. 1). Senescence was delayed in 1974 due to available moisture in October and November. In 1976 cold temperatures in September terminated seasonal growth. Soil moisture and air temperature determined the time of quiescence.
Table 1. Dates of phenological stages and range of their occurrence in 1974, 1975, and 1976 for bush muhly.
Phenology 1974
Vegetative 7116 Flowering 8/ 14 Senescence 1 l/20 Quiescence 12/ 15
Dates 1975
7128 9/ 17 10122 11/6
1976 Range (Days)
7120 12
9112 34
9129 52
lo/ 12 63
Three years of clipping bush muhly did not significantly influ- ence TNC levels until the close of the 1976 growing season. Defo- liating bush muhly continuously through the growing season or during maturity significantly reduced TNC levels below that found in nondefoliated plants or plants only clipped during the vegetative stage for three years (Table 2). Prior to the end of the third year, concentration of TNC did not respond to defoliation. VFS and VSV treatments had less effect on TNC levels than did the other four treatments. Although the VFS and VSV treatments included defoliation during maturity, earlier defoliation periods two years in three appeared to nullify this late clipping effect.
Bush muhly elongates the internodes on all culms very early during the vegetative stage. This morphological characteristic allows easy accessibility of apical meristems and axillary buds to grazing. The number of elongated internodes produced per culm was very sensitive to the previous year’s clipping treatment (Table 3).
During vegetative growth in 1975 bush muhly plants produced significantly more internodes per culm on nondefoliated plants and plants defoliated during the vegetative stage in 1974 (including the vegetative, VFS and VSV treatments) than plants defoliated continuously or during maturity in 1976. There was no significant difference in internode numbers per culm in 1975 between plants
Table 2. TNC concentration (%) of bush muhly roots durine auiescence 1976 from plants subjected to different clippinp treatments. Phenological stage of clipping
1974 Control Vegetative Vegetative Vegetative
1975 Control Vegetative Flowering Senescence
1976 Control Vegetative Senescence Vegetative
% TNC 18.5”’ 18.7” 1 5.5”b 16.7”b
‘Data followed by a different letter are significantly different at the 95% level, by Tukeys w-procedure.
Maturity Continuous Maturity Continuous Maturity Continuous
14.5b 14.2b
Table 3. Mean number of elongated stem internodes on bush muhly prior to defoliation in 1975 and 1976 during the vegetative stage.
Number of internodes during vegetative stage
Defoliation treatment 1975 1976
Control 1974 and 1975 ll.la’ 11.8”
Vegetative 1974 and
1975 9.0a 9.1b
Vegetative 1974
Flowering 1975 9.9” 8.4b
Vegetative 1974
Senescence 1975 10.6” 5.4”
Maturity 1974 and 1975 5.1b 4.9”
Continuous 1974 and
1975 4.7b 4. lC
‘Data in columns followed by a different letter are significantly different at the 95% level, by Tukey’s w-procedure.
defoliated in the vegetative stage in 1976 and non-defoliation plants.
Following two years of defoliation all clipped plants showed a decrease in the number of elongated internodes per culm in 1976 regardless of season of defoliation. However, plants defoliated after the flowering stage showed a significantly greater detrimental effect than plants defoliated prior to or during flowering. The data indicate that removing 65% of the current year’s growth in two consecutive years during the vegetative stage is detrimental to the number of elongated internodes produced per culm. However, if defoliation occurs after flowering for only one growing season, the number of elongated internodes produced per culm will be only 50% of that produced by non-defoliated plants.
Removing 65% of the leaf area during the vegetative stage accounted for approximately 30% of the total annual growth in
1974 (Table 4). This was about 50% less than the amount of forage obtained from the mature or continuous treatments.
In 1975, the second year of clipping, production sharply decreased from that produced in 1974. Although it was impossible to compare the clipping effect across years due to climatic varia- tion, production of four other species studied in the area did not decrease as drastically (Miller 1976). Plants defoliated early in the
growing season in 1974 produced significantly more herbage than those clipped continuously or at maturity througout the vegetative flowering and senescence stages in 1975 (Table 4). Clipping during maturity was as detrimental to forage production as continuous defoliation.
In 1976, the third year of defoliation, production appeared to continue to decline. Again climatic variation must be taken into account. Bush muhly clipped in the vegetative stage for two consec- utive years produced significantly more forage during this pheno- logical stage than plants subjected to the continuous or VSV treatments during 1976 (Table 4). This response was similar to internode response.
Crown diameter was significantly larger in non-defoliated plants than in defoliated plants during the 1975 growing season (Table 5). Following the 1976 growing season crown diameters in plants continuously defoliated for a three-year period, continued to decrease significantly as compared to non-defoliated plants. The control plants showed no change in size while continuously defol- iated plants decreased in diameter by 4 cm.
Conclusions
The decrease in TNC levels, internode numbers, yield, and crown diameter in 1976 indicated continuous or late defoliation of bush muhly had a greater detrimental impact than any of the remaining clipping treatments. Further, the average number of internodes and plant material harvested during the vegetative stage in 1976 indicated that the seed ripening stage may have been the plants’ most vulnerable stage to harvesting. Clipping in the vegeta- tive stage in three consecutive years had the least detrimental effect of the five clipping treatments. Yield and TNC levels were optimal with defoliation practices during vegetative growth. However, removing 65% of the leaf area on bush muhly during any of the four growth stages reduced plant vigor as observed by yield and number of internodes present. Only the continuous and maturity defolia- tions resulted in a significant decrease in TNC levels. The vegeta- tive treatment may have had less impact on the plants since only 30% of the total season’s growth was harvested. The decrease in production from 1974 through 1976 on the five clipping treatments appeared to be caused by more than climate alone. The reduction in both crown diameters and elongated internodes due to clipping suggested defoliation to be part of the cause for the sharp decrease
Table 4. Mean dry weight of forage harvested per bush muhly plant from five defoliation treatments during 1974,1975 and 1976. Data are accumulative within years across phenology.
Phenological stage at time of clipping
1974 1975 1976
Treatment Ve&_ Flow Sen Qui Veg Flow Sen Qui Veg Flow Sen Qui
Vegetative 34.5& 11.9” 4.5”
VFS 34.8” 12.3” 2.3”
vsv 46.8” 20.7” 0.9b
Maturity 60.4” 72.9” 5.7b 7.1” 0.5b 0.5b
Continuous 36.1” 45.3 55.1” 60.3” l.lb 2.3b 7.2b 9.4” 0.6b 0.7 2.0” 2.0”
IAbbreviations are: Veg = Vegetative, Flow = Flowering, Sen = Senescence, Qui = Quiescence. These treatments reflect the period of defoliation for 1974, 1975 and 1976 respectively.
ZData in columns followed by a different letter are significantly different at the 95% level by Tukey’s w-procedure.
Table 5. Mean annual change in crown diameter (cm) per bush muhly plant during the 1975 and 1976 growing seasons.
Phenological stage at time of clipping
1974 Control Vegetative Vegetative Vegetative
1975 Control Vegetative Flowering Senescence
1976 Control Vegetative Senescence Vegetative
1975 6.4” 2.0b1 l.ob -0.5b
1976 o.oa -0.5ab -2.3ab - 1 .6ab
IData in rows followed by different letters are significantly different at the 95% level by Tukey’s w-procedure.
Maturity Continuous Maturity Continuous Maturity Continuous
-0.7b -2.0b
-0.7”b -4.0b