www.wjpr.net Vol 6, Issue 4, 2017. 41
BETA-SITOSTEROL STIMULATE PHAGOCYTOSIS IN HEPG2
CELL LINE TRANSFECTED WITH HEPATITIS B SURFACE
ANTIGEN
Ammar M. Lubeiny1-2*, Wuyun Li2, Xu Lin2 and Elshaikh A. Elobied3
1
Neelain Institute for Medical Research, Faculty of Medicine, Al-Neelain University,
Khartoum, Sudan.
2
Key Laboratory of Ministry of Education for Gastrointestinal Cancer, Fujian Medical
University Fuzhou, China.
3
Research and Grant Unit, Ahfad University for Women, Khartoum, Sudan.
ABSTRACT
Hepatocellular carcinoma (HCC) has a range of risk factors and
Hepatitis B Virus infection considers as one of these factors which
lead to develop HCC. This type of carcinoma is still classified as a
complex disease due to many risk factors and the patients with HCC
diagnosed always at advance stage. The surgery and transplantation
are still the most effective treatment while the response to
chemotherapy and radiotherapy has poor effect. Our aim is to study
the effect of β-Sitosterol, which is found in many plants on HBsAg
and on hepatocellular carcinoma. To address this issue, Huh7,
HepG2.2.15( harbouring four copies of HBV DNA ) and HepG2 cell
lines were used. The Cell viability assay, Enzyme – Linked
Immunosorbent Assay, and Gene Expression were used to evaluate the effect of
β-Sitosterol. The result of Gene Ontology shows it is effect on HepG2-HBsAg by stimulating
the phagocytosis we conclude β-Sitosterol has a positive regulation of phagocytosis to
HepG2 Transfected with HBsAg this process could be help full to eliminate the dead cells
caused by the apoptotic effect of β-Sitosterol on hepatocellular carcinoma cell infected with
hepatitis B virus.
KEYWORDS: Beta- sitosterol, phagocytosis, HCC, Hepatitis B virus.
Volume 6, Issue 4, 41-51. Research Article ISSN 2277– 7105
*Corresponding Author
Dr. Ammar M. Lubeiny
Neelain Institute for Medical
Research, Faculty of
Medicine, Al-Neelain
University, Khartoum,
Sudan.
Article Received on 22 Jan. 2017,
Revised on 12 Feb. 2017, Accepted on 05 March. 2017
INTRODUCTION
The poor prognosis of HCC due to late and advance stage and a wide range factors
classified it as complex disease. Moreover cannot responsive to standard chemotherapy and
resist to radiotherapy while in the most cases the surgical and liver transplantation remains
only the choice for treatment. There for the development of novel effective treatment is
prime important.[1] The recent studies using a combination of different technologies, like
genomics, proteomics, metabolomics, MicroRNA studies, and Bioinformatics, are
providing new understanding of the gene expression and protein profiles to develop novel
cancer therapeutics .[2] A 30% of the global populations are infected with HBV and they are
at risk of developing liver fibrosis, HCC, and other illness.[3, 4, and 5] In this study, we used
DNA microarray to evaluate the effect of Beta sitosterol on Hepatoma cell HepG2
transfected with HBsAg. The HBsurface (HBs) proteins, consist of large, middle and small
(SHBs) (LHBs, MHBs, SHBs) proteins and it are possibly distinguished by their different
domains and glycosylation status.[6] The carboxyl terminal domain containing SHBs is
present in all surface-proteins, preS1 N-terminal extension only in LHBs, preS2 in LHBs
and MHBs.[7]. These three forms of HBV surface proteins represent HBsAg.[8] and
synthesized in the endoplasmic reticulum and could lead to the situation that amounts of
protein produced exceed those required for virion assembly. Excess surface proteins
undergo multimerization resulting in their budding from the ER/Golgi compartment as both
non-infectious spherical and filamentous sub viral particles (SVP) or as versions.[9] The
SVPs typically outnumber the virions, could be components of circulating immune
complexes.[10], and may induce immune tolerance by a mechanism of “viral apoptotic -like
mimicry” [11].
The domains of HBV surface proteins are S, preS2 domain is 55 aa long, and
preS1. PreS1 contains 108, 118, or 119 amino acids (aa), depending on the genotype, and
S-domain (S) PreS extensions of LHBs and MHBs possess certain features that are very
important for HBV infectivity such as binding the nucleocapsid during virus envelopment [9]
HBsAg could be involved in the arrangement, of the immune responses, especially in
disturbances of the appropriate immune responses. [12, 13, 14 and 15]. The HCC carcinogenesis
caused by HBV has been studied and the relations with HBV were proof as a multiple
factors process that includes a direct mechanism involving viral protein, indirect
mechanisms through the chronic inflammation, and the integration of HBV DNA.[16] The
direct mechanism of the viral protein reported in the HBx gene.[17, 18] and PreS2 gene.[19]
based on a transgenic mouse model and force expression model of the cell lines act as
mitogen-activated protein kinase (MAPK), which is involved in cell proliferation.[20] Moreover,
PreS2 protein accumulates in the endoplasmic reticulum (ER) of hepatocytes, and DNA
injury is caused in the cell by ER stress [21, 22] we subjected the Hepatoma cell lines Huh7,
HepG2 and HepG2.2.215 to β-Sitosterol to evaluate it is an effect of this compound which
can be found in many plants it will be important to evaluate its function on HBV and HCC.
Materials
β-Sitosterol (C29H50O) (Sigma Aldrich analytical standard) prepared by dissolved
0.995mg β-Sitosterol in 10 ml Ethanol HepG2.2.15 cells (CRL-11997; ATCC) harboring
four copies of HBV-DNA. HepG2 from American Type Culture Collection (ATCC). Cell
counting kit 8 CCK8, DOjinDO laboratories, Japan. G418 Geneticin anti-biotic (Invitrogen)
Method
Cell culture HepG2.2.15, Huh 7and HepG2 Cell lines were cultured in Dulbecco's
Modified Eagle Medium (DMEM) (Invitrogen) with 10%FBS (Fetal Bovine Serum) from
Gibco® and G418 Geneticin (Invitrogen) at 380 μg/ml added to HepG2.2.15
Cell viability assay: Cell Counting Kit 8 (CCK8, DOjinDO laboratories, Japan) was used
for the Cell viability determination. 8x103 cells from Huh7 HepG2 and HepG2.2.15were
counted (Count star Biotech Automated Cell Counter Shanghai) and cultured in 100 µl
DMEM with 10% FBS in 96 wells plate for 24 hours, β-Sitosterol added in different
concentration from 0.0, 60, 120, 240 μM . Incubated for 24 hours to determine the cell
viability in selected time. One hour before measuring the viability of the cells, The DMEM
were changed with 100 µl of 1:10 CCK-8 solution and DMEM and incubated for one hour
at 37°C in 5% CO2 incubator. Then we measured the absorbance at 450 nm using a micro
plate reader (ELx800 Absorbance Reader). Care was taken not to introduce bubbles to the
wells during changing the medium, since they interfere with the O.D. reading.
Enzyme – Linked Immunosorbent Assay (ELISA).
Detection of Hepatitis B Surface Ag: We incubate the HepG2.2.15 cells in plate till a
90% confluence was reached .Then β-Sitosterol in ethanol at 60 μM concentration were
added and incubated for 24 hours at 37°C in 5% CO2 incubator. the reagents kit used from
Wantai Beijing China and we followed the company protocol. One hundred µl taken from
control were added. Then 50μl from diluents were added to all the wells, the plate were shook slowly, cover and incubated for one hour at 37°C in 5% CO2 incubator. 50μl of
conjugate added and incubated for 30 minutes at 37°C in 5% CO2 incubator. The reagent
was decanted and the plate was tapped to remove any reagent from the wells. Cells were
washed by adding wash solution four times; incubated after addition of wash solution for
two minutes each time. Then 50μl of Chromogen A and 50μl Chromogen B solution were
added incubated for 30 minutes at 37°C in 5% CO2 incubator. We stopped the reaction by
adding 50μl from the stop solution and read at 450nM. Then we measured the absorbance at
450 nm using a microplate reader (ELx800 AbsorbanceReader). Care was taken not to
introduce bubbles to the wells during changing the medium
Plasmid construction: Plasmids included HBs cDNA, were constructed and used as
templates for polymerase chain reaction (PCR) amplification. PCR-generated HBx genes
were inserted into the KpnI and XhoI sites of the pcDNA3.1/Hygro (+) plasmid (V870–20,
Invitrogen), the resultant vectors were called pcDNAhygro+- HBs the primers used for PCR
amplification is shown as followed:
S-hygro (+) cloneF: 5’ - CGGGGTACC GCC ACC ATG GAGAACATCGC ATCAGG -3’
S-hygro (+) cloneR: 5’ - CCGCTCGAG CTA CTTGTCGTCA TCGTCTTTGT
AGTCAATGTA TACCCAAAGA-3’
Transfection: HepG2cell line were cultured in DMEM supplemented with 10% FBS in 10
cm plate till a 90% confluence reached then the 2ml trypsin were added for 3mints and
removed 3ml of DMEM were added, HepG2 cells were counted and seeded in 6-wells plate
at the density of 5× 105 per well 24 hours before transfection. The cells would reach 90%
confluent and the medium changed to DMEM without FBS and incubated for one hour
before transfection. Plasmids were transected into HepG2 cells using lipofectamine 3000
(Invitrogen, Carlsbad, CA) prepared in two test tubes one as reference containing 2.87 µl
pcDNAhygro+- plasmid and Salmon sperm DNA 12.4 µl,P3000™ 4 µl, Opti-MEM 80.73
µl the total amount 100.00 µl and the second tube PcDNAhygro+- HBs 2.56 µl , Salmon
sperm DNA 0.0 µl, P3000™ 4µl, Opti-MEM 93.44 µl the total amount 100.00 µl , In
another two tubes we prepared lipofectamine as in table 1 five minutes before transfection
we added the tubes of lipofectamine with the tubes containing PcDNAhygro+- HBs and
pcDNAhygro+- plasmid respectively. And transfer it to HepG2 cells in 10 cm plate. 24
the medium with the addition of Hygromycin 800μg/ml till reached the clear clone, and then
we picked the clones into 24 wells plate after the cells propagated, the cells were passage
into 12, 6 wells, 6 cm, and 10cm plate and propagated.
Table – 1 two tubes prepare with lipofectamine 3000
Component Amount
Lipo3000 3µl
Opti-MEM 97µl
Total amount 100µl
Gene chip
After transfection the HepG2-HBsAg, were cultured in DMEM with10% FBS in 10 cm
plates till reached a 90%confulence .Then the 60 μM of β-sitosterol were added and
incubated at 37 °C, 5% CO2 for 24 hours then the DMEM removed and 1.5 ml of Trizol
(Invitrogen) added to each plate We used Affymetrix gene chip and Agilent gene chip.
Total RNA quality control, mark, chip hybridization, washing, staining and scanning, and
data extraction and analysis procedure as company protocol. Affymetrix micro array for
detection of mRNA expression profiles of total RNA samples as the starting, and
biotin-labeled in vitro amplification, markAmbion #1792 cRNA expansion tags Kit. Data
extraction and analysis software using the AGCC (Affymetrix® Gene Chip® Command
Console® Software) chip . Fluorescent Scanned image is saved as DAT files and analyzed
with KEGG PATHWAY, P ID, PANTHER, REACTOME, BIOCART, BIO CYCL
software.
RESULT
Cell viability: Cell viability: Cell viability by CCK8 result shows decreases in viable cells
related to concentration of β-Sitosterol in three cell lines In previous study we proofed that β-sitosterol has effect on HepG 2 and HepG2.2.15 cell lines by reducing viable cells .It is
0 0.2 0.4 0.6 0.8
0 60 120 240
Figure 1 shows the viable cells in Huh7 Cell line y axis represented the number of
viable cell, x axis represented the concentration of Sitosterol column 1- No
β-Sitosterol were added, column 2- 60 μM β-β-Sitosterol , column 3-120 μM β-β-Sitosterol,
column 4- 240 μM β-itosterol
Effect of β-Sitosterol on HBsAg
The optical reading (OD) was indicating that 60 μM of β-sitosterol has an effect on
HepG2.2.15by reducing the total amount of HBsAg. as shown in figure 2.
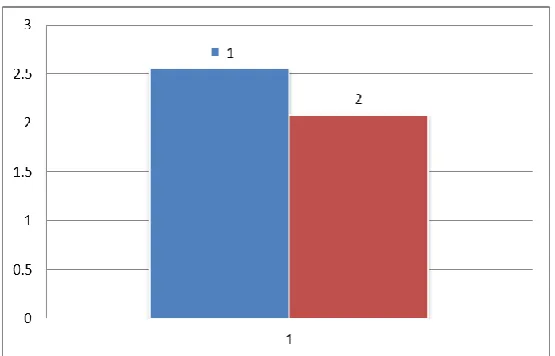
Figure- 2 ELISA test for HBsAg quantification 1 is a reference shows the amount of
HBsAg in HepG2.2.15 Column 2shows the amount of HBsAg in HepG2215with 60 μM
beta-sitosterol this figure shows the decreased in HBsAg amount in cells treated with
beta-sitosterol
Gene Expression
[image:6.595.143.465.76.197.2] [image:6.595.163.441.373.551.2]Figure 3 Electrophoresis for quantification of the protein in the samples for
microarray test Lane 6 HepG2 HBsAG, Lane10 HepG2- PcDNA-hygro treated with
β-sitosterol, M Lane Hela Cell Control RNA
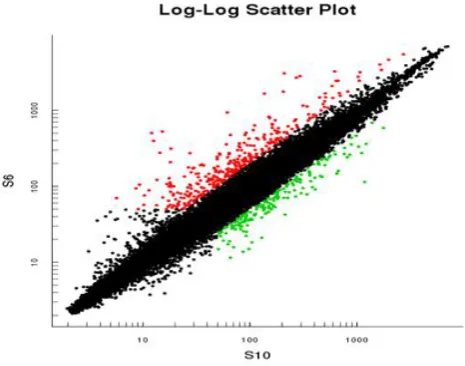
Figure 4 - Log-log scatter gives a visual comparison of gene level variation between
two cell lines. S6 refer to HepG2-HBsAg, the S10 is HepG2 cell line as reference both
the cell lines were treated with β-sitosterol red color represented up regulation of
genes, while green color down regulation and black equal expression in both reference
and sample
Gene Ontology
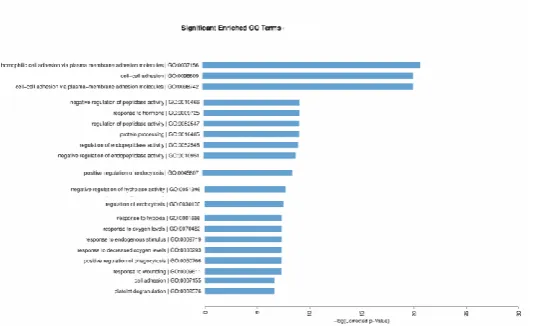
The result of Gene Ontology (GO) biological process increased the phagocytosis and
activation c3 and c5 pathways and increasing cell adhesion the –log (corrected p-value)
shows there are increased expression on several genes as shown in figure 7 and enrichment
[image:7.595.199.406.72.215.2] [image:7.595.172.405.307.490.2]Figure 5 Gene Ontology (GO) biological process increased in several genes involve in
biological process in HepG2-HBsAg
DISCUSSION
Phagocytosis is the way toward taking in microorganisms, parasites, dead cells, and foreign
debris by a cell [23].When β-sitosterol add to HepG2-HBsAg stimulate the phagocytosis by
positive regulation of phagocytosis/GO: 0050766 in HepG2cell line our previous study
showed β-sitosterol has an effect on HepG2-HBx by inducing the extrinsic apoptotic
signaling pathway more over ; it has senescence inducing effect to HepG2-HBSP, This
herbal compound (β-sitosterol) has anti-proliferative effect on Huh7cell line and its effect
on HepG2 , HepG.2.2.15shown in previous study. The extensive study by DNA microarray
and Gene Ontology studies under the biological process revealed that β-sitosterol can
stimulate the phagocytosis in HepG2-HBsAg .The important of the Phagocytosis of dying
cells is a major process that be the final stage of cell death in a tissue under basal
conditions, in a diseased tissue like cancer or after anticancer therapies.[24] The cluster of
differentiation (CD47) is a commonly expressed molecule on all cancers. Its function to
block phagocytosis is known, and blockade of its function leads to tumor cell phagocytosis
and elimination. CD47 is therefore a validated target for cancer therapies.[25] Studies from
the I.L.W. laboratory have shown that malignant cancers up-regulate the “don’t eat me”
signal CD47, presumably in their progression to allow escape from endogenous “eat me”
signals that were induced as part of apoptosis and programmed cell removal.[25, 26, 27and 28].
Treatment of Caski and HeLa cells with β-sitosterol significantly suppressed the expression
of genes involved in cell proliferation and oncogenic transformation, while augmenting the
expression of genes involved in apoptosis and tumor suppression. Ultrastuctural
anti-proliferative and anti-cancer activity of this natural product isolated from traditional
Chinese herbs.[29] We believe that β-sitosterol can stimulate the phagocytosis in
hepatocellular carcinoma infected with Hepatitis B Virus and by this process can eliminate
the defragmented and dead cells which occurs due to apoptotic effect of β-sitosterol on
hepatocellular carcinoma and HBV.
CONCLUSION
β-sitosterol has anti-proliferation effect on HCC cell lines Huh7, HepG2and HepG2.2.15
and stimulate the positive regulation of phagocytosis process in HepG2.2.15 by direct effect
on HBsAg. Therefore β-sitosterol has great effect on hepatitis B Virus replication and this is
the first study to evaluate the effect of β-sitosterol on HepG2-HBsAg as for our knowledge
and could open new areas of research on hepatitis B virus pathogenesis.
ACKNOWLEDGEMENT
This work done by the scholar position from the key laboratory of ministry of education for
gastrointestinal cancer Fujian medical university China.
REFERENCES
1. Aravalli, Rajagopal N., Clifford J. Steer, and Erik N. Cressman. "Molecular
mechanisms of hepatocellular carcinoma." Hepatology 48.6 (2008): 2047-2063. Print.
2. Chen, X. "Gene Expression Patterns in Human Liver Cancers." Molecular Biology of
the Cell 13.6 (2002): 1929-1939. Print.
3. Gabr S.A., Alghadir A.H. Prediction of fibrosis in hepatitis C patients: Assessment
using hydroxyproline and oxidative stress biomarkers. Virusdisease. 2014; 25: 91–100.
doi: 10.1007/s13337-013-0182-8.[PMC free article] [PubMed] [Cross Ref]
4. Trépo C., Chan H.L., Lok A. Hepatitis B virus infection. Lancet. 2014; 384: 2053–2063.
doi: 10.1016/S0140-6736(14)60220-8. [PubMed] [Cross Ref].
5. McMahon B.J. Natural history of chronic hepatitis B. Clin. Liver Dis. 2010; 14: 381–
396. doi: 10.1016/j.cld.2010.05.007. [PubMed] [Cross Ref].
6. Bremer, Corinna, and Dieter Glebe. "The Molecular Virology of Hepatitis B
Virus." Seminars in Liver Disease, vol. 33, no. 02, 2013; 103-112.
7. Glebe, Dieter. "Viral and cellular determinants involved in hepadnaviral entry." World
Journal of Gastroenterology, 2007; 22(13)1.
8. lich, Wolfram H. "Medical Virology of Hepatitis B: how it began and where we are
9. Bruss, Volker. "Hepatitis B virus morphogenesis." World Journal of Gastroenterology,
2007; 13(1): 65.
10. Vanlandschoot, Peter, and Geert Leroux-Roels. "Viral apoptotic mimicry: an immune
evasion strategy developed by the hepatitis B virus?" Trends in Immunology, 2003;
24(3): 144-147.
11. Sorrell, Michael F. "National Institutes of Health Consensus Development Conference
Statement: Management of Hepatitis B." Annals of Internal Medicine, 2009; 150(2):
104.
12. M. L. Op den Brouw, R. S. Binda, M. H. van Roosmalen et al., “Hepatitis B virus
surface antigen impairs myeloid dendritic cell function: a possible immune escape
mechanism of hepatitis B virus,” Immunology, 2009; 126(2): 280–289. X View at
Publisher · View at Google Scholar · View at Scopu.
13. Y. Chen, H. Wei, R. Sun, and Z. Tian, “Impaired function of hepatic natural killer cells from murine chronic HBsAg carriers,” International Immunopharmacology, 2005; 5:
13-14, 1839–1852, View at Publisher · View at Google Scholar · View at Scopus.
14. P. Vanlandschoot, F. van Houtte, A. Roobrouck, A. Farhoudi, and G. Leroux-Roels, “Hepatitis B virus surface antigen suppresses the activation of monocytes through
interaction with a serum protein and a monocyte-specific receptor,” Journal of General
Virology, 2002; 83(6): 1281–1289, View at Google Scholar · View at Scopus.
15. Carey, L. D'Antiga, S. Bansal et al., “Immune and viral profile from tolerance to
hepatitis B surface antigen clearance: a longitudinal study of vertically hepatitis B
virus-infected children on combined therapy,” Journal of Virology, 2011; 85(5): 2416–2428,
View at Publisher · View at Google Scholar · View at Scopus.
16. F. Imazeki, M. Omata, O. Yokosuka, and K. Okuda, “Integration of hepatitis B virus
DNA in hepatocellular carcinoma,” Cancer, 1986; 58(5): 1055–1060, View at Google
Scholar · View at Scopus.
17. K. Koike, T. Tsutsumi, H. Fujie, Y. Shintani, and K. Moriya, “Molecular mechanism of
viral hepatocarcinogenesis,” Oncology, vol. 62, supplement, 2002; 1: 29–37. View at
Publisher · View at Google Scholar · View at Scopus.
18. H. L. Y. Chan and J. J. Y. Sung, “Hepatocellular carcinoma and hepatitis B virus,”
Seminars in Liver Disease, vol. 26, no. 2, pp. 153–161, 2006. View at Publisher · View
at Google Scholar · View at Scopus.
transcriptional transactivator,” Nature, 1990; 343(6257): 457–461. View at Publisher ·
View at Google Scholar · View at Scopus.
20. J. H. Hung, I. J. Su, H. Y. Lei et al., “Endoplasmic reticulum stress stimulates the
expression of cyclooxygenase-2 through activation of NF-κB and pp38
mitogen-activated protein kinase,” Journal of Biological Chemistry, 2004; (279)45: 46384–
46392. View at Publisher · View at Google Scholar · View at Scopus.
21. H. C. Wang, H. C. Wu, C. F. Chen, N. Fausto, H. Y. Lei, and I. J. Su, “Different types
of ground glass hepatocytes in chronic hepatitis B virus infection contain specific pre-S
mutants that may induce endoplasmic reticulum stress,” American Journal of Pathology,
2003; 163(6): 2441–2449. View at Google Scholar · View at Scopus.
22. H. Kadowaki, H. Nishitoh, and H. Ichijo, “Survival and apoptosis signals in ER stress: the role of protein kinases,” Journal of Chemical Neuroanatomy, 2004; 28: 1-2, pp. 93–
100, View at Publisher · View at Google Scholar · View at Scopus.
23. Ernst JD, Stendahl O, ebrary Inc. Phagocytosis of bacteria and bacterial pathogenicity.
Cambridge: Cambridge University Press, 2006.
24. Garg, A. D., Romano, E., Rufo, N., & Agostinis, P. (2016). Immunogenic versus
tolerogenic phagocytosis during anticancer therapy: mechanisms and clinical
translation. Cell Death and Differentiation, 23(6): 938-951. doi:10.1038/cdd.2016.5.
25. Majeti R, et al. (2009) CD47 is an adverse prognostic factor and therapeutic antibody
target on human acute myeloid leukemia stem cells. Cell 138(2): 286–299.
CrossRefMedlineWeb of ScienceGoogle Scholar.
26. Willingham SB, et al. (2012) The CD47-signal regulatory protein alpha (SIRPa)
interaction is a therapeutic target for human solid tumors. Proc Natl Acad Sci USA
109(17): 6662–6667.
27. Chao MP, et al. (2011) Therapeutic antibody targeting of CD47 eliminates human acute
lymphoblastic leukemia. Cancer Res 71(4): 1374–1384. Abstract/FREE Full Text.
28. Chao MP, et al. (2011) Extranodal dissemination of non-Hodgkin lymphoma.
29. Cheng, D., Guo, Z., & Zhang, S. (2015). Effect of β-sitosterol on the expression of HPV
E6 and p53 in cervical carcinoma cells. Współczesna Onkologia, 1: 36-42.