Operant conditioning of aerial respiratory behaviour in Lymnaea stagnalis
9
0
0
Full text
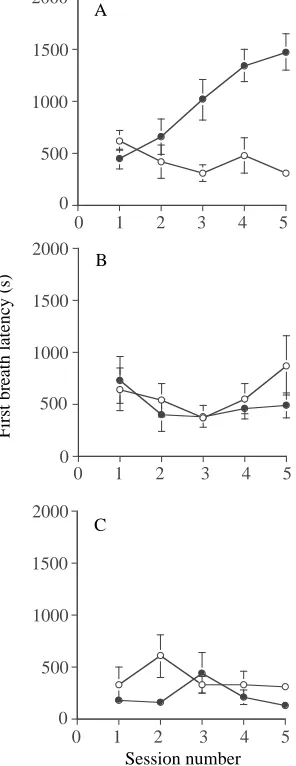
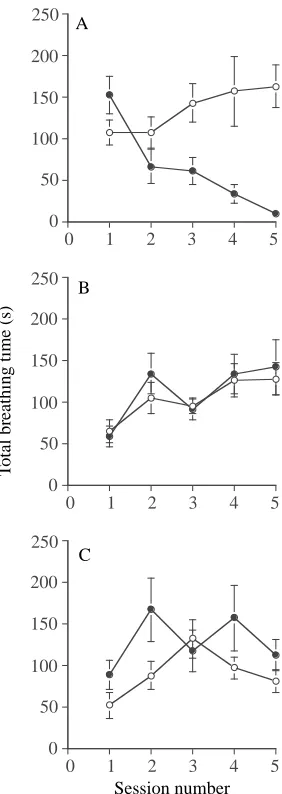
Figure

Related documents
