With 7 text-figures Printed in Great Britain
PHYSIOLOGY OF INSECT RHYTHMS
III. THE TEMPORAL ORGANIZATION OF THE ENDOCRINE EVENTS UNDERLYING PUPATION OF THE TOBACCO HORNWORM
BY JAMES W. TRUMAN* AND LYNN M. RIDDIFORD*
The Biological Laboratories, Harvard University, Cambridge, Massachusetts 02138 U.S.A.
(Received 14 August 1973)
The moulting process of insects is governed by the interaction of three hormones: prothoracicotropic hormone (PTTH), juvenile hormone, and ecdysone (see Doane, 1973; Wyatt, 1972; Wigglesworth, 1971 for recent reviews). To initiate the moult the brain releases PTTH which causes the secretion of ecdysone from the prothoracic glands. Ecdysone in turn triggers the detachment of the epidermis from the old cuticle and the subsequent secretion of the new cuticle. Whether this new cuticle is larval, pupal, or adult depends on the titre of juvenile hormone at the time of ecdysone secretion.
In larvae of the tobacco hornworm, Manduca sexta, the secretion of PTTH by the brain is regulated by a photosensitive circadian clock (Truman, 1972). The interaction of this clock with the environmental photoperiod serves to restrict hormone release to a specific portion of the light-dark cycle. Consequently, if a larva becomes competent to release hormone after the close of one of these temporal' gates', it must then wait for the opening of the gate on the following day.
The gating of PTTH release facilitated a quantitative study of the temporal re-lationship of the activity of the endocrine organs which were involved in the moult to the fifth larval stage (Truman, 1972). The present paper considers the timing of the endocrine events which govern pupation in the tobacco hornworm.
MATERIALS AND METHODS A. Experimental animals
The M. sexta stock was derived from animals obtained from Dr R. A. Bell, A.R.S., U.S.D.A., Fargo, N. D. Larvae were fed with the Bell modification of the Yamamoto (1969) hornworm diet. Each animal was reared at 25 °C in an individual container as described by Truman (1972).
B. Photoperiod conditions
Larvae were reared in constant-temperature rooms under a 12L: 12D photoperiod cycle. In this paper lights-off has been arbitrarily designated at midnight (24.00). According to convention (Pittendrigh, 1965), times are given as arbitrary "zeitgeber" times (A.Z.T.).
372 JAMES W. TRUMAN AND LYNN M. RIDDIFORD
24 12
Fig. i. The timing of the entry into the wandering stage of a group olManduca larvae which were reared under a i z L : 12D photoperiod at 25 °C.
RESULTS
A. The timing of overt morphological events during the late fifth instar andprepupal stages
During the first part of the fifth instar, Manduca larvae feed continuously. But by the 4th or 5th day after the ecdysis an abrupt change occurs in their appearance and behaviour. Within a span of a few hours, feeding ceases, the gut is cleared, the epidermis along the dorsal midline becomes somewhat transparent thereby exposing the heart, and a pink pigment appears on the dorsum of the abdomen along either side of the heart. Larvae which have undergone this transition are referred to as 'wandering larvae'.
In a 12L: 12D regimen at 25 °C, hornworm larvae undergo ecdysis to the fifth instar during the 14 h period from 22.00 to 12.00 A.Z.T. (Truman, 1972). Collection of fifth-stage larvae each morning thereby yields groups of developmentally synchronous larvae which have a mean ecdysis time of 03.00 A.Z.T. Within such a group approxi-mately one-half of the individuals become wandering larvae on the fourth night after ecdysis. The remainder of the caterpillars then wait through the following day and make the transition to the wandering stage during the fifth night.
373
V 1/
1
DayO
I
Purging of the gut, etc.
Day 1 ' Ocellar retraction
Day 2 Day 3 Day 4
\
Pupal ecdysis Feeding larva Wandering larva Prepupa
Fig. 2. The timing of the stages between the cessation of feeding by the larva and^the pupal ecdysis. The arrows give the median time of occurrence of the respective transitional events.
0
<-12.00 16.00 20.00 Time of ligation (AZT)
24.00
Fig. 3. The response of neck-ligatured fifth-instar larvae which were feeding as a function of the time of ligation; • — • indicates those which subsequently showed an exposed heart; A — • indicates those which became pink. Each point represents approximately 30 animals. At 25 °C the events occurring after the beginning of the wandering stage have a relatively rigid temporal sequence (Fig. 2). The wandering stage lasts for almost 2 days until the retraction of the ocellar pigment marks the entry into the prepupal phase. Approximately 3 days thereafter the pupa then undergoes ecdysis. In this paper the age of animals will be given relative to the time of entry into the wandering stage on day o.
B. The endocrine control over the entry into the wandering stage
1. The effect of ligation between the head and thorax
In the puss moth, Cerura vinula, the last-stage larva turns pink after it ceases feeding. This pink coloration was shown to be due to the ecdysone-induced deposition of ommochrome pigments (Biickmann, 1959). It therefore seemed likely that in Manduca the gated appearance of the pink coloration (as well as other features of a wandering larva) was similarly due to the secretion of ecdysone and, accordingly, to the gated release of PTTH.
374 JAMES W. T R U M A N AND L Y N N M . RIDDIFORD
of a given day, one can be assured that it will make the transition during the following night. In order to examine the possible role of PTTH in causing this change, groups of feeding larvae which had attained 7 g by lights-on were subsequently neck-ligatured at various times of day. The condition of the animals on the following morning was then determined. The ligature seemed to interfere with the purging of the gut; con-sequently, this manifestation was not considered. The effects of ligation on the exposure of the heart and on the appearance of the pink pigment are seen in Fig. 3. These two characters responded differently to ligation in that the body required longer contact with the head in order to become pink than it did to expose the heart. When larvae were ligatured at 17.00, one-half showed a prominent heart when examined on the following morning, but only 10% showed any sign of pink coloration. When ligatures were applied 3-5 h later at 20-30, then 50% of the larvae subsequently became pink. Thus, the critical period for the head's involvement in the exposure of the heart occurs about 3-5 h earlier than for the production of the pink pigment. Moreover, the function of the head is completed approximately 7-10 h before the morphological characters actually appear.
2. Effect of double ligations
From the above-mentioned work on Cerura vinula we assumed that the entry into the wandering stage in Manduca sexta was due to ecdysone which was released from the prothoracic glands. In order to determine the relationship between the brain and these glands, each of a series of 400 feeding larvae of 7 g or larger was ligatured around the neck and between the second and third abdominal segments. The next morning both the thoracic and abdominal compartments were checked for exposure of the heart and for pink pigmentation.
In the majority of animals there was no difference between the two compartments; when the thorax showed only an exposed heart, the abdomen also showed only an exposed heart, etc. In a few instances the thorax was more affected than was the abdomen. However, it is important that there were also individuals in which the re-spective responses of the thorax and abdomen were reversed; e.g., the thorax had an exposed heart whereas the abdomen showed both pink pigmentation and the exposed heart. The significance of these results will be discussed below.
C. Timing of endocrine events necessary for pupation
375
100 r80
60
& .0 40
20
— 02.30
— 16.30 ~ 18.30 — 14.30 — 12.30
[image:5.451.82.375.47.252.2]Days after the onset of the wandering larva stage
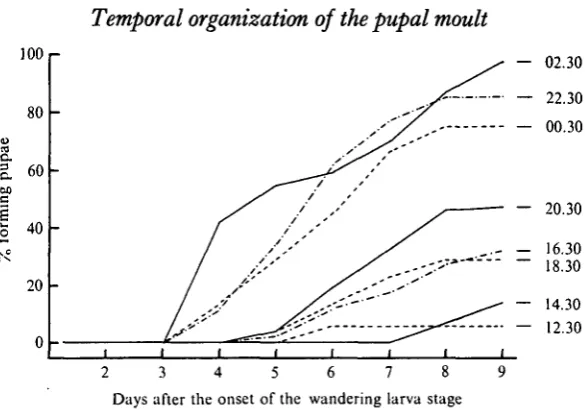
Fig. 4. The time-course for pupation for groups of Manduca larvae which were ligatured at various times during day i and day 2 (00.30 and 02.30) after the onset of the wandering stage. The time of ligation is indicated to the right of each curve. Each curve is based on approximately 30 larvae.
1. Effects of neck-ligations
Groups of Manduca larvae were neck-ligatured at various times during day o and day 1 of the wandering stage. The larvae were then checked at daily intervals and the number moulting to pupae was noted. Usually the animals did not shed the larval skin, but the pupal characters were clearly evident beneath the thin larval cuticle. To assure that we were scoring moults which were driven by the tropic activity of the brain, only
Manduca forming pupae by day 9 after the beginning of the wandering stage were
considered. After this time the number of deaths among ligatured larvae was significant. Ligatures which were applied to wandering larvae during day o were very effective in stopping further development. None of these larvae formed pupae by day 9. The results were considerably different when the ligatures were applied during day 1. As seen in Fig. 4, the percentage of each group which subsequently formed pupae was a function of the time of ligation. When larvae were neck-ligatured at 12.30 of day 1, only 6% became pupae by day 9. But if ligatures were applied 14 h later, at 02.30 of day 2, then 97 % of the animals had pupated within the same period.
Fig. 4 shows that the rate of pupation was also a function of the time of ligation. The few Manduca which formed pupae after ligation at 14.30 did so during days 8 and 9. By contrast, the group ligatured at 02.30 (day 2) showed 50% pupation by day 4 - the time for the pupal ecdysis of control animals.
sub-376 JAMES W. TRUMAN AND LYNN M. RIDDIFORD
0
<-12.00 16.00 20.00 24.00 Time of ligation (AZT)
04.00
Fig. 5. The response of neck-ligatured wandering larvae as a function of the time of ligation; • — % those animals of each group which became pupae by day 9 after the beginning of wander-ing; • — A those which became pupae by day 4. Each point represents approximately 30 larvae. sequently became pupae by day 9. As stated above, these animals pupated significantly faster than did those animals which moulted solely in response to the autonomous activity of the prothoracic glands. Thus it appears that in the 'average larva' secretion of PTTH by the brain occurs at least during the 7 h span from 20.00 to 03.00 A.Z.T.
2. Isolated abdomens
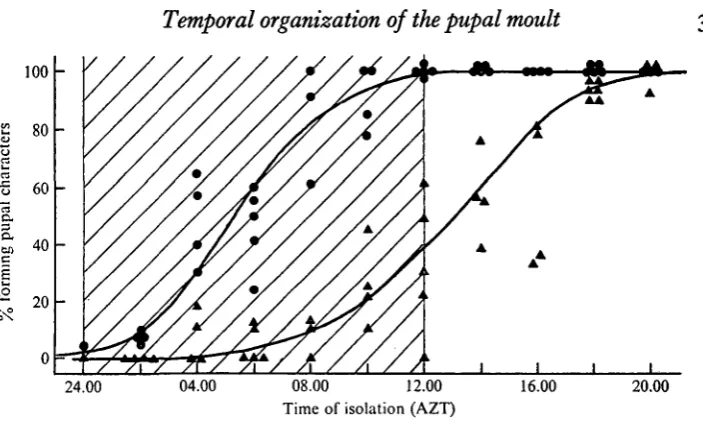
The time of activity of the prothoracic glands was determined by isolating abdomens at various times of day. A ligature was placed between the first and second abdominal segments, and the thorax was then cut away. Abdomens which subsequently formed pupae routinely did so on day 4 or 5 after the transition to the wandering stage (2 to 3 days after ligation). In a series of over 1000 isolated abdomens none became pupal later than day 6. This was expected since the abdomens contain no prothoracic glands. The percentage of abdomens which became pupal was clearly dependent upon the time of isolation (Fig. 6). By 13.00 of day 2, 50% of the abdomens were able to form normal pupal abdomens in the absence of the prothoracic glands. This time is only 10 h later than when the brain is no longer required to obtain the normal time-course of pupation.
377
24.00 04.00 08.00 12.00 Time of isolation (AZT)
[image:7.451.45.397.46.258.2]16.00 20.00
Fig. 6. The developmental response of isolated fifth-instar abdomens as a function of the time of isolation; • — • abdomens which showed at least the formation of the pupal 'gin traps'; A—A abdomens which became totally pupal. Each point represents about 25 abdomens.
first piece of epidermis becomes competent to moult. By approximately 05.00, one-half of the abdomens showed at least the production of the gin-trap cuticle. These data indicate that the prothoracic glands are required for at least 8 h to produce a complete pupal moult of the abdomen.
DISCUSSION
A. The temporal organization of hormone secretion by the fifth-instar larva
The ligation experiments reported here serve to define times at which certain endocrine centres are no longer required for a defined biological response. These responses are the complex result of a number of factors, and therefore the quantitative interpretation of the data solely in terms of changes in endocrine activity may eventually prove to be not entirely correct. However, we think that interpreted in this manner the data give a good approximation of minimum durations of endocrine activity and serve to identify times when events of endocrinological interest are occurring.
1. The transition to the wandering-stage larva
In the Lepidoptera and in many other holometabolous insects the behaviour of the larva changes dramatically at the end of the last instar. Feeding ceases and behaviour is directed towards finding or preparing a suitable site for pupation. This transition in behaviour is marked by a purging of the gut and, occasionally, by other morphological and physiological changes.
378 JAMES W. TRUMAN AND L Y N N M . RIDDIFORD
In both responses the involvement of the brain appears to be through its stimulation of ecdysone secretion by the prothoracic glands. From the work of Biickmann (1959) on Centra vinula we originally assumed that the pink coloration in Manduca was due to ecdysone. In homworm larvae this character is somewhat labile - the intensity of pigmentation varies greatly and the colour fades during day 1 of the wandering stage. It has proved to be very difficult to produce the pink pigment by injection of ecdysone into neck-ligatured feeding larvae. Single injections of various doses of ecdysone have not produced pink coloration in Manduca, but slow infusion of 40 fig of yff-ecdysone over 12 h has occasionally provoked a weak pink pigmentation (H. F. Nijhout, in preparation). The exposure of the heart has been an easier response to obtain. Injections of moderate doses of /?-ecdysone routinely cause exposure of the heart in isolated larval abdomens (H. F. Nijhout, in preparation).
In light of the above results the double ligation experiments become especially puzzling because they fail to demonstrate a need for the prothoracic glands. Indeed, even when the ligatures were applied during the middle of the PTTH release gate, the thoracic and abdominal compartments routinely showed the same response. The few instances in which the thorax progressed further than the abdomen seem to suggest that a thoracic centre is involved. But these cases are countered by other doubly ligatured larvae in which the abdomen showed more pronounced characters than did the thorax.
This dilemma can be resolved by assuming that the relationship between the brain and prothoracic gland is somewhat different in provoking the transition to the wander-ing stage than it is in causwander-ing a moult. In the latter case the prothoracic glands become independent of the brain at least in part through a positive feedback effect of the ecdysone which they have released (Williams, 1952). As will be discussed below, in provoking the pupal moult of Manduca the brain is active for at least 7 h before the pro-thoracic glands become ' turned-on' to their maximum extent. In the case of producing wandering larvae the 3-5 h activity period for the brain may simply not be long enough to produce enough ecdysone to turn-on the glands. Thus, when the brain's influence is blocked by ligation, the secretory activity of the prothoracic glands quickly subsides. Accordingly, the period of hormone release by these glands is essentially the same as that for the brain - i.e. from approximately 17.00 to 20.30.
2. The formation of the pupa
In the formation of the pupal stage by Hyalophora cecropia, approximately 4 days elapse between the critical period for the brain and that for the prothoracic glands (Williams, 1952). Calpodes ethlius is similar in that the time between the two critical periods is 90 h (Locke, 1970). The data from Fukuda (1944) indicate a value of approxi-mately 80 h for Bombyx mori, and in Galleria mellonella there is one to two days between the two events (Beck, 1970). Manduca sexta has the most compressed time-table yet reported - only 10 h separate the time after which the brain is no longer required for normal pupation and that after which the prothoracic glands are not necessary.
the rate of pupation on the time of ligation, these differences must then reflect changes in prothoracic gland activity. Williams (1968) and Sehnal (1972) have demonstrated that abnormally large amounts of ecdysone can accelerate the rate of development. It is therefore reasonable to presume that the slowed development observed in early ligatured Manduca is the result of the prothoracic glands secreting ecdysone either in smaller amount or at a slower rate than do those glands which have experienced the full tropic influence of the brain.
Fukuda (1944) reported that abdomens isolated from Bombyx mori larvae during a certain period would subsequently undergo an incomplete moult. In Manduca this response was seen when abdomens were isolated at approximately 05.00 on day 2. By 13.00 the entire abdomen had become competent to pupate after isolation. The spread of competence through the abdominal epidermis during this 8 h period requires the presence of the prothoracic glands and, most likely, gives a minimum estimate for the period of active secretion by these glands. The fact that ocellar retraction occurs at 23.00 on day 1 indicates that prothoracic gland secretion may begin as early as 20.00 - the time that the tropic activity of the brain is first detected. Thus these glands may be releasing hormone for as long as 17 h in order to produce a complete pupal moult.
B. Comparison of the timing of the endocrine events for a larval versus a pupal moult
In the fourth instar Manduca sexta larva PTTH secretion occurs at approximately 00.30 A.Z.T. This is followed by a juvenile hormone surge at 01-30 and ecdysone release from 03-30 to 06-30 (Truman, 1972). Fig. 7 compares the temporal relationship of endocrine happenings in the penultimate instar with those in the last-stage larva.
The most striking difference between the two moults is that the formation of the pupa is a two-step process. The first release of PTTH transforms the larva into the wandering stage, whereas the second initiates retraction of the epidermis and synthesis of the pupal cuticle. This is in accordance with the findings of Shaaya & Karlson (1965) for Cerura vinula in which they find that ecdysone appears in the blood on two occasions - the first when the larva becomes pink and the second during the prepupal period. Also Hintze (1968) has presented histological evidence in Cerura for increased pro-thoracic gland activity during these same two times.
Although the overt morphological changes are minor, the transition into the wander-ing stage is important for the eventual metamorphosis of the larva. In the Cecropia silkworm juvenile hormone can produce larval-pupal intermediates only if it is applied prior to the beginning of this wandering period; after the onset of this stage the epi-dermis is unresponsive to exogenous hormone (Riddiford, 1972). A similar relationship also holds for Manduca sexta (Truman, Riddiford & Safranek, in preparation). Ap-parently in response to ecdysone at the outset of the wandering stage the larval epidermis makes the irrevocable commitment to moult to the pupal condition.
The second, third, and fourth stage larva released PTTH during gates which occurred in the scotophase of a i2L:i2D photoperiod (Truman, 1972). In the last instar the PTTH gate shifted into the middle of the photophase. This pronounced change in the time of PTTH secretion may be related to the fact that during the last instar the tonic level of circulating juvenile hormone drops to vanishingly small titres
38o JAMES W. TRUMAN AND LYNN M. RIDDIFORD
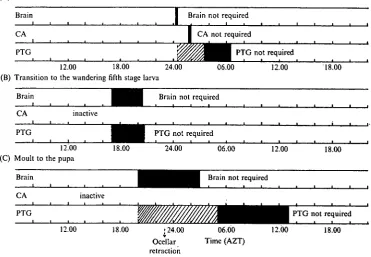
(A) Moult to the fifth larval instar
Brain Brain not required
CA CA not required
PTG PTG not required
12.00 18.00 24.00 (B) Transition to the wandering fifth stage larva
06.00 12.00 18.00
Brain
CA inactive
PTG
Brain not required
PTG not required
12.00 (C) Moult to the pupa
18.00 24.00 06.00 12.00 18.00
12.00 18.00 [24.00
Ocellar retraction
06.00 Time (AZT)
[image:10.451.42.414.84.341.2]12.00 18.00
Fig. 7. A schematic summary of the timing of the endocrine events which are required for: (A) the moult to the fifth instar; (B) the transition to the wandering larva; (C) the moult to the pupa. The black bars are minimal estimates of the time of active secretion of the glands. The cross-hatched bars indicate additional periods of probable secretion.
(H. F. Nijhout, in preparation). The significance of such a relationship, however, is uncertain.
In Truman (1972) it was thought that the PTTH required for the moult to the fifth stage was secreted during a relatively brief time. This conclusion was based on the failure to find any quantitative differences in developmental response when larvae were neck-ligatured at various times during the PTTH gate. With the transition to the pupal stage PTTH secretion is definitely not a brief event. During the first release, secretion occurs during at least a 3-5 h period from 17.00 to 20.30. The second release of PTTH lasts for at least 7 h.
The length of time that the prothoracic glands are active also differs in the fourth and fifth instars. Based on the frequency of mosaic moults in isolated abdomens, it was concluded that the prothoracic glands were actively secreting in the fourth instar for at least 3 h. More likely secretion began soon after the brain released PTTH which would give a duration of 6 h (Truman, 1972). In the transition to the wandering stage the minimum duration of prothoracic gland activity is relatively short - 3-5 h. However, in causing the onset of prepupal stage the glands secrete for 8 h and perhaps for as long as 17 h. Thus the prothoracic glands are active for almost 2-5 times longer for the pupal moult as compared with a moult to another larval stage.
the time course for the remainder of each moult is similar. In the moult to the fifth stage larva 44 h elapse between the time that the epidermis is independent of the prothoracic glands and the time of ecdysis (Truman, 1972). With pupation this period is approxi-mately 46 h. Manifestly, after the epidermis has become independent of the prothoracic glands, it requires no more time to make a pupal cuticle than it needs to synthesize a larval cuticle.
SUMMARY
1. Late in the fifth instar, Manduca sexta larvae cease feeding and become ' wander-ing larvae' which are morphologically characterized by an 'exposed heart' and the appearance of a pink pigment along the dorsal midline. Two days later ocellar retraction signals the beginning of the prepupal period and 3 days thereafter the pupal ecdysis occurs.
2. The timing of the endocrine events which are responsible for these changes was determined by ligaturing animals of the appropriate developmental stage at various times of day. The times of prothoracicotropic hormone (PTTH) release by the brain were determined by neck ligations. Estimates of the times of prothoracic gland activity were obtained through the isolation of abdomens.
3. It was found that the fifth stage larva releases PTTH on two occasions. The first release lasts approximately 3*5 h and triggers the transformation to the wandering stage. The second release occurs two days later, lasts at least 7 h, and provokes the onset of the pupal moult.
4. The prothoracic glands are involved in triggering the same two changes. In the first instance the glands apparently require the continuing influence of the brain and consequently secrete for about 3-5 h. During the stimulation of the pupal moult the prothoracic glands become 'turned-on' and continue to secrete for at least 10 h after the time when the brain is no longer required. In this latter instance the total time of prothoracic gland activity may be as long as 17 b.
We are grateful to Mr H. F. Nijhout for many discussions during the course of this study and for a critical reading of the manuscript. The technical assistance and sug-gestions of Mr Louis Safranek are also gratefully acknowledged. This work was sup-ported by the Harvard Society of Fellows, a grant from the Rockefeller Foundation, and NSF grants GB-35540, GB-24963, and GB-36645X.
Parts of this study were carried out at the Concord Field Station of Harvard University.
REFERENCES
BECK, S. D. (1970). Neural and hormonal control of pupation in Galleria mellonella (Lepidoptera: Galleriidae). Ann. ent. Soc. Am. 63, 144-9.
BOCKMANN, D. (1959). Die Auslosung der Umfa'rbung durch das Hautungshormon bei Cerura vinula L. (Lepidoptera, Notodontidae). jf. Insect. Physiol. 3, 159-89.
DOANE, W. W. (1973). Role of hormones in insect development. In Developmental Systems: Insects, vol. 2 (eds. S. J. Counce and C. H. Waddington), pp. 291-497. London: Academic Press.
FUKUDA, S. (1944). The hormonal mechanism of larval molting and metamorphosis in the silkworm.
J. Fac. Sci. Tokyo Imp. Univ. 6, 477-532.
HINTON, H. E. (1946). The 'gin-traps' of some beetle pupae; a protective device which appears to be unknown. Trans. Roy. ent. Soc. Lond. 97, 473-96.
382 JAMES W. TRUMAN AND LYNN M. RIDDIFORD
HINTZE, C. (1968). Histologische Untersuchungen iiber die AktivitSt der inkretorischen Organe von
Cerura vinula L. (Lepidoptera) wahrend der Verpuppung. Roux Archiv. 160, 313-43.
JUDY, K. J. (1972). Diapause termination and metamorphosis in brainless tobacco hornworms (Lepi-doptera). Life Sci. n , 605-11.
LOCKE, M. (1970). The molt/intermolt cycle in the epidermis and other tissues of an insect Calpodes
ethlius. Tissue Cell a, 197-223.
PITTENDRIGH, C. S. (1965). On the mechanism of entrainment of a circadian rhythm by light cycles. In
Circadian Clocks (ed. J. Aschoff), pp. 276-97. Amsterdam: North-Holland Publ. Co.
RIDDIFORD, L. M. (1972). Juvenile hormone in relation to the larval-pupal transformation of the Cecropia silkworm. Biol. Bull. mar. biol. Lab., Woods Hole 142, 310-25.
SEHNAL, F. (1972). Action of ecdysone on ligated larvae of Galleria mellonella L. (Lepidoptera): induc-tion of development. Ada ent. Bohemoslov. 69, 143-55.
SHAAYA, E. & KARLSON, P. (1965). Der Ecdysontiter wahrend der Insecktenentwicklung. IV. Die Entwicklung der Lepidopteren Bornbyx mori L. und Cerura vinula L. Devi. Biol. 11, 424-32. TRUMAN, J. W. (1972). Physiology of insect rhythms. I. Circadian organization of the endocrine events
underlying the moulting cycle of larval tobacco hornworms. J. exp. Biol. 57, 805-20.
WIGGLESWORTH, V. B. (1971). Insect Hormones, pp. 159. San Francisco: W. H. Freeman and Co. WILLIAMS, C. M. (1952). Physiology of insect diapause. IV. The brain and prothoracic glands as an
endocrine system in the Cecropia silkworm. Biol. Bull. mar. biol. Lab., Woods Hole 103, 120—138. WILLIAMS, C. M. (1968). Ecdysone and ecdysone-analogues: their assay and action on diapausing
pupae of the Cynthia silkworm. Biol. Bull. mar. biol. Lab., Woods Hole 134, 344-55.
WYATT, G. R. (1972). Insect hormones. In Biochemical Actions of Hormones, vol. 2 (ed. G. Litwack), pp. 385-490. London: Academic Press.
YAMAMOTO, R. T. (1969). Mass rearing of the tobacco hornworm. II. Larval rearing and pupation.