Signalling Pathways Mediated by
the Bombesin/GRP Receptor
Amanda Charlesworth
A Thesis submitted for the degree of Doctor of Philosophy
at the University of London
1 9 9 6
G ro w th R egulation Laboratory
D epartm ent o f O ncology ^
Im p e ria l C ancer
ProQuest Number: 10106528
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10106528
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
A
b s t r a c t
Neuropeptides, and in pa rticu la r peptides o f the bom besin fa m ily, in clu d in g gastrin releasing peptide, stim ulate DNA synthesis and cell p ro h fe ra tio n in cultured cells and are im p lic a te d as ce llular g ro w th facto rs in a va rie ty o f fu n d a m e n ta l biological processes including development, tissue regeneration and tumourigenesis.
Bombesin elicits m u ltip le signalling pathways in various cell types, in clu d in g activa tio n o f phosphohpase C, w hich leads to activation o f p ro te in kinase C and elevation o f in tracellular calcium; tyrosine phosphorylation o f in tra ce llu la r substrates and reorganisation o f the actin cytoskeleton, and cell p ro life ra tio n . It is n o t clear however w hether these responses are m ediated by a single receptor subtype or by d iffe re n t subtypes that couple preferentially to specific pathways. To resolve this the mouse bom be sin/GRP re ceptor was tra n sfected in to Rat-1 fib ro b la s ts and the pathw ays a ctivated b y bom be sin were in vestigated. Here it is show n th a t phosphohpase C activation, ceh grow th and tyrosine phosphorylation emanate fro m a single class o f bombesin receptor.
M itogen A ctiva ted Protein Kinase (MAPK) a c tiv ity is a com m on p o in t o f convergence fo r m itogenic signals. While the mechanism by w hich receptor tyrosine kinases activate MAPK is weU estabhshed, the mechanism by which G protein-coupled receptors activate MAPK is less weU understood. The involvem ent o f Ras and Raf in activa tio n o f MAPK has n o t been described in bom besin-induced signalhng. This thesis shows th a t in Rat-1 cells tran sfected w ith the bom besin/GRP receptor, bom besin can activate MAPK and Raf-1 in a p ro te in kinase C-independent fashion. Also it is dem onstrated fo r the firs t tim e the bom besin stim ulates GTP loading o f
p 2 i r a s
A
c k n o w le d g e m e n ts
I w o u ld lik e to th a n k everyone who made th is w o rk possible. F irst and forem ost I w ould like to thank Hemy fo r his insight, supervision and guidance, and fo r the m any valuable lessons he has taught me about staying ahve in science.
In a d d itio n I w o uld like to thank M ark Bouzyk, Simon Broad, Theresa Higgins and Jim Sinnett-Smith fo r getting me started in the lab and also Thomas Seuffelein and Dom inic W ithers fo r keeping me going.
Special thanks go to Sara Rankin fo r showing me what can be achieved and together w ith Theresa and Ahson McDowell fo r m y "personal development". Many thanks to Perugino's fo r keeping me fed during m y long evenings by the com puter and to the Canadian M u ffin Co. who kept me going during the long Sundays.
1 w o u ld also hke to acknowledge everyone who had to p u t up w ith m y bad m oods and constant m oaning d u rin g the home straight. In p a rtic u la r 1 am m ost grateful to the Lab-down-the-end who took me away when 1 needed it most, and when 1 didn't.
And I m ust n o t forget the guys in photography who had to do just-one-m ore- emergency on several occaisions. Finally 1 m ust thank Sharon Matthews fo r giving this thesis a rather excellent once over.
C
o n t e n t s
A
b s t r a c t2
A
ck n o w led g em en ts3
C
o n t e n t s4
L
is t o fF
igures9
L
is t o fT
ables12
A
b b r e v ia tio n s13
Chapter 1
INTRODUCTION
16
1.1.
G
r o w t hR
eg u l a t io n17
1.1.1. Growth Factors...18
1.1.2. Synergistic Signalling Pathw ays...21
1.1.3. Regulatory Signals and Obligatory Events... 22
1.1.4. Receptor Tyrosine Kinases... 22
1.1.5. G Protein-Coupled Receptors...24
1.1.5.1. Growth Regulation by G Protein-Coupled Receptors... 25
1.1.5.2. S tructu re... 26
1.1.6. Heterotrim eric G-Proteins... 28
1.1.6.1. S tructu re... 28
1.1.6.2. The G Protein Cycle... 29
1.1.6.3. Classes o f G P ro te in ... 31
1.1.6.4. py su b u n its...31
1.1.6.5. M odifying T o x in s ...33
1.2. B
om besin34
1.2.1. Bombesin/GRP Receptor... 361.3.
B
om besin-M
ed ia t edS
ig n a lT
r a n s d u c t io n3 7
1.3.1. Phosphohpase C ...371.3.2. Inositol 1,4,5-trisphosphate and Ca^+ Mobihsation... 38
1.3.2.1. Calcium H om eostasis... 39
1.3.2.2. Calcium Release fro m Intracellular Stores...42
1.3.2.3. Calcium In flu x fro m the Extracellular M edium ...44
1.3.2.4. Calcium G radients...46
1.3.2.5. Calcium in Growth C o n tro l... 46
1.3.2.6. Calcium Effectors...49
1.3.3. Protein Kinase C ...50
1.3.3.1. Structure and A c tiv a tio n ... 51
1.3.3.2. Substrates... 52
1.3.3.4. Protein Kinase D ...53
1.3.4. Monovalent Ion Fluxes...54
1.3.5. Phospholipase A2 and Arachidonic Acid Release... 55
1.3.6. A denylyl Cyclase, cAMP and PK A ... 56
1.3.7. Tyrosine Phosphorylation...57
1.3.7.1. Tyrosine Phosphorylation o f M ultiple Substrates...57
1.3.7.2. Focal Adhesions...58
1.3.7.3. Stress Fibres... 59
1.3.7.4. S rc ...60
1.3.7.5. Focal Adhesion Kinase... 61
1.3.7.6. pl30c=«...61
1.3.7.7. P axillin... 61
1.3.7.8. Bombesin-Induced FAK and Paxillin Tyrosine P h o sp h o ryla tio n 62 1.3.7.9. Role o f FAK and Paxillin Tyrosine Phosphorylation...63
1.3.8. The MAPK P athw ay... 64
1.3.8.1. O verview ...64
1.3.8.2. M APK...65
1.3.8.3. MEK...66
1.3.8.4. R a f...66
1.3.8.5. Ras...68
1.3.8.6. Effects o f MAPK... 70
1.3.8.7. Activation o f the MAPK Pathway by G Protein-Coupled Receptors . 70 1.3.8.8. Bombesin-Induced MAPK A c tiv a tio n ...73
1.3.8.9. Cellular C o nte xt...74
1.3.8.10. Function o f GPy...75
1.3.8.11. Stress-Activated Protein Kinases... 76
1.3.9. Induction of Early Genes... 77
1.3.9.1. AP-1 ... 77
1.3.9.2. Induction o f AP-1 C om ponents... 78
1.4.
COUPUNG TO MULTIPLE PATHWAYS8 0
1.5.
A im s
OF THESIS8 3
Chapter 2 - MATERIALS AND METHODS
85
2 .1 .
M
a te r ia ls85
2 .2 .
M
e t h o d s8 6
2.2.1. Transfection of Rat-1 cells... 862.2.2. CeU C u ltu re ... 86
2.2.22. Swiss 3T3 CeUs...86
2.2.3. [125IJGRP binding assay... 87
2.2.4. Measurement of Total Inositol Phosphates...87
2.2.5. Measurement of Ca^+... 88
2.2.5.1. Using the fluorescent Ca^+ indicator, Fur a-2 ...88
2.2.5.2. Measurement o f Total ^^^Ca^+ C o n te n t... 89
2.2.6. Measuring PKC Activation hy 80K/MARCKS Phosphorylation...89
2.2.7. MAPK A c tiv a tio n ...90
2.2.7.1. Immune Complex Assay fo r p42”^®P*^ A c tiv a tio n ... 90
2.2.7.2. p42™apkBandshift... 90
2.2.8. Raf-1 Kinase Assay...91
2.2.8.1. Preparation o f Substrates...91
2.2.8.2. Raf-1 Kinase Assay...91
2.2.9. GTP Loading of Ras... 92
2.2.10. Analysis of Tyrosine Phosphorylated Proteins... 93
2.2.10.1. Tyrosine Phosphorylation o f M ultiple P roteins...93
2.2.10.2. Tyrosine Phosphorylation o f FAK...93
2.2.10.3. Tyrosine Phosphorylation of P a xillin ...93
2.2.11. Measurements of cellular proliferation... 94
2.2.11.1. [^HlThymidine Incorporation Assay... 94
2.2.11.2. Autoradiography o f Labelled N u c le i... 95
2.2.11.3. Measurement o f cell p ro h fe ra tio n ...95
2.2.12. Pretreatm ents... 95
Chapter 3 - Signalling Pathways Coupled to The
Bombesin/GRP Receptor
97
3.1. Binding Characteristics of the Expressed Bomhesin/GRP Receptor 97 3.1.1. Expression o f the bombesin/GRP receptor in Rat-1 cells... 973.1.2. Timecourse o f [i^^IjGRP Binding to Transfected Rat-1 Cells...98
3.1.3. Ligand Recognition p ro file o f the Transfected Bombesin/GRP Receptor... 98
3.2. The Transfected Bomhesin/GRP Receptor Functionally Couples to PLC-Activated Signalling Pathways...103
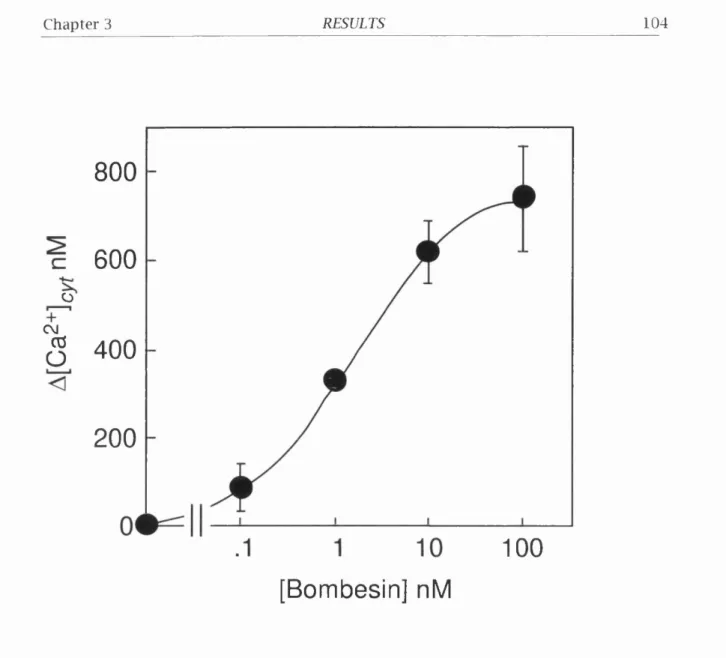
3.2.1. Bombesin Stimulates Intracellular Ca^+ M o b ilis a tio n ...103
3.2.2. Bombesin stimulates 80K/MARCKS P hosphorylation...103
3.3. Expression of the homhesin/GRP receptor confers a grow th response to bombesin in Rat-1 cells...107
3.3.2. Bombesin Induces Cell P ro life ra tio n ... 107
3.4. Bom besin Stim ulates Tyro sin e Phosphorylation in Rat-1 cells Transfected w ith the Bombesin/GRP Receptor... I l l 3.4.1. Bombesin induces tyrosine phosphorylation o f m u ltip le proteins in BOR 5 and BOR 15 cells... I l l 3.4.2. Bombesin-Induced Tyrosine Phosphorylation is in h ib ite d by the specific bombesin receptor antagonist...116
3.4.3. Bombesin-Induced Tyrosine Phosphorylation is Dependent on the In te g rity o f the A ctin Cytoskeleton...116
3.4.4. Tyrosine Phosphorylation induced by Bombesin is independent o f PKC...120
3.4.5. Bombesin stimulates tyrosine phosphorylation o f FAK and paxillin in BOR 5 and BOR 15 cells...120
3.5. Summary and Discussion...123
3.5.1. M ultiple Coupling fro m one Receptor...123
3.5.2. M ultiple Coupling via py Subunits...124
3.5.3. M ultiple coupling via a d istinct Ga s u b u n it...125
Chapter 4 - The MAPK Pathway Stim ulated By
Bombesin in Rat-1 Cells
130
4.1. Bombesin Stim ulation of MAPK a c tiv ity in Rat-1 Cells Transfected w ith the Bombesin/GRP Receptor... 1304.1.1. Bombesin Stimulates MAPK a c tiv ity ... 130
4.1.2. Dose Response o f Bombesin-Induced MAPK activity in BOR 15 CeUs...131
4.1.3. Bombesin-stimulated MAPK activity is independent o f PKC in BOR 1 5 ... 131
4.2. Bombesin Stimulation of Raf-1 A c tivity in Transfected Rat-1 cells 135 4.2.1. Bombesin Stimulates Raf-1 activation in BOR 15 CeUs... 135
4.2.2. Bombesin-Induced Raf-1 A ctivation is n ot Dependent on PK C 135 4.2.3. Pertussis Toxin does n ot In h ib it Bombesin-Induced Raf-1 A c tiv ity ... 138
4.3. Bombesin Stimulation of Ras.GTP Loading in Transfected Rat-1 CeUs. 138 4.4. Summary and Discussion...142
4.4.1. M ultiple MEK A ctiva tors...142
4.4.2. Bombesin activation o f Ras. Mediation by Gpy?... 143
4.4.3. CeUular C o nte xt... 143
5.1. Effect of Thapsigargin and DBHQ on PDB-Stimulated DNA Synthesis.. 146
5.1.1. Thapsigargin and DBHQ. Stimulate [^HJThymidine Incorporation in synergy w ith PDB...146
5.1.2. Kinetics o f Thapsigargin and DBHQ Stimulated [^HJthymidine Incorporation is Consistent w ith Entry to S-Phase... 150
5.1.3. Thapsigargin and DBHQ increase the Proportion o f Cells that Reinitiate DNA Synthesis... 150
5.2. Effect of DBHQ w ith other m itogens... 153
5.2.1. Sub-maximal Concentrations o f Bombesin Synergise w ith Thapsigargin and DBHQ... 154
5.3. Exam ination o f the Synergism Between PKC and Thapsigargin or DBHQ... 154
5.3.1. The Synergistic Stim ulation o f DNA Synthesis Requires Functional PKC...154
5.3.2. Thapsigargin and DBHQ do not potentiate PDB-mediated signal tran sduction... 158
5.4. Effect of Thapsigargin and DBHQ on intracellular calcium sto res 161 5.4.1. Thapsigargin and DBHQ in h ib it Bombesin-Induced Ca^+ M o b ü isa tio n ... 161
5.4.2. Kinetics o f Thapsigargin and DBHQ in h ib itio n o f Bombesin-Induced Ca^+ M obihsation... 161
5.4.3. Thapsigargin and DBHQ do not Prevent Bombesin-Induced Inositol Phosphate A ccu m ulation... 165
5.4.4. Thapsigargin and DBHQ cause Prolonged Depletion o f Total Cellular Ca^+... 165
5.5. Effect of extracellular calcium on DNA synthesis... 167
5.6. Summary and Discussion...172
5.6.1. Capacitative C a^+E ntry... 172
5.6.2. Synergy w ith PKC... 173
5.6.3. Cytotoxic e ffe c ts ...174
5.6.4. Possible Pathways A c tiv a te d ... 175
Chapter 6 - SUMMARY AND PERSPECTIVES
178
REFERENCES
183
L
is t
o f
F
ig u r es
Figure 1.1. Diagram o f PDGF receptor w ith sites o f interaction w ith signal
transduction molecules...24
Figure 1.2. Schematic diagram o f a seven transmembrane G-protein-coupled re c e p to r... 27
Figure 1.3. Probable arrangement o f behces in seven transmembrane dom ain receptors... 27
Figure 1.4. G pro te in cycle... 30
Figure 1.5. A m ino acid sequence o f mammaban and am phibian bom besin-like p e p tid e s... 35
Figure 1.6. Bombesin-stimulated signal transduction pathw ays... 37
Figure 1.7. Calcium Homeostasis and Release...42
Figure 1.8. Structure o f PKC isozym es...52
Figure 1.9. lin e a r diagram o f p a xillin structure... 64
Figure 1.10. Receptor tyrosine kinase activation o f the MAPK pathw ay... 71
Figure 1.11. Mechanism o f MAPK activation by G protein coupled receptors...73
Figure 1.12. M ultiple mammaban MAPK m o d u le s... 77
Figure 1.13. A ctivation o f AP-1 tran scription fa c to r... 79
Figure 1.14. Possible mechanisms o f m u ltip le coupling fo r bombesin re c e p to rs ...81
Figure 3.1. Specific [i^^l]GRP binding to d ifferent clo n e s... 99
Figure 3.2. Binding o f [i^sij-GRP as a fu n ctio n o f GRP concentration... 100
Figure 3.3. Time course o f [i^^IjGRP binding to BOR 5 c e b s ...101
Figure 3.4. Com petition Binding o f [i^^ljGRP to BOR 5 cebs...102
Figure 3.5. Bombesin-induced increase in [Ca^+J^yt in BOR 5 cebs... 104
Figure 3.6. Antagonist in h ib itio n o f bombesin-induced increase in [Ca^+J^yf... 105
Figure 3.7. Bombesin-induced phosphorylation o f 80K/MARCKS... 106
Figure 3.8. Kinetics o f [^Hjthym idine incorporation induced by bom be sin... 108
Figure 3.9. Effect o f antagonist on bombesin-induced [^H]thym idine in corp oration... 109
Figure 3.10. Ceb prohferation induced by bombesin in BOR 5 cebs... 110
Figure 3.11. Bombesin-induced tyrosine phosphorylation o f m u ltip le proteins ( I) ...112
Figure 3.12. Bombesin-induced tyrosine phosphorylation o f m u ltip le proteins (b )... 113
10
Figure 3.14. Kinetics o f bombesin in duction o f tyrosine phosphorylation o f
m u ltip le proteins (h )... 115 Figure 3.15. Effect o f antagonist on bombesin-induced tyrosine
phosphorylation o f m u ltip le p ro te in s ...117 Figure 3.16. Effect o f Cytochalasin D on bombesin-induced tyrosine
phosphorylation o f m u ltip le p ro te in s ...118 Figure 3.17. Effect o f GF 109203X on bombesin-induced tyrosine
phosphorylation o f m u ltip le p ro te in s ...119 Figure 3.18. Bombesin-induced tyrosine phosphorylation o f FAK and Paxillin
(1)...121 Figure 3.19. Bombesin-induced tyrosine phosphorylation o f FAK and Paxillin
(H)...122 Figure 3.20. D ifferent areas in the 3rd cytoplasmic loop o f a2A-AR-clO are
required fo r coupling to G a i4 vs G a q /n ...128 Figure 4.1. p4 2“ apk activation induced by various fa c to rs ... 132 Figure 4.2. Bombesin-induced increase in p42“^®P*^ a c tiv ity ... 133 Figure 4.3. Effect o f PKC in h ib itio n and down regulation on
bombesin-stim ulated MAPK a c tivity...134 Figure 4.4. Time course o f bombesm-induced Raf-1 a c tiv ity ... 136 Figure 4.5. Effect o f PKC in h ib itio n on Raf-1 activity induced by various
m itogens... 137 Figure 4.6. Effect o f pertussis toxin on Raf-1 activity induced by various
m itogens... 139 Figure 4.7. Bombesin stim ulation o f loading (I)... 140 Figure 4.8. Bombesin stim ulation o f Ras.GTP loading (11)... 141 Figure 5.1. Dose response curve fo r the stim ulation o f DNA synthesis by
DBHQ. or th a p sig a rg in ... 147 Figure 5.2. Effect o f the length o f preincubation w ith DBHQ and thapsigargin
on the subsequent stim ulation o f DNA synthesis by these
compounds in synergy w ith PDB... 148 Figure 5.3. Dose response curve fo r the stim ulation o f DNA synthesis by PDB.... 149 Figure 5.4. Time course fo r in corporation o f [^Hjthym idine in the presence o f
thapsigargin and DBHQ...151 Figure 5.5. Effect o f thapsigargin and DBHQ on DNA synthesis assessed by
autoradiography and hqu id scin tillation counting o f
[^H]thymidine in c o rp o ra tio n ...152 Figure 5.6. Effect o f DBHQ on DNA synthesis induced by bombesin assessed
b o th by autoradiography and hquid scin tilla tio n counting o f
11
Figure 5.7. Effect o f down regulating PKC on [^H]thym idine incorp oration
induced by DBHQ and either PDB or b om be sin ...156 Figure 5.8. Effect o f in h ib itin g PKC on [^Hjthym idine incorp oration induced
by DBHQ and either PDB or bom besin...157 Figure 5.9. Effect o f thapsigargin and DBHQ on PDB-mediated signal
transduction ( I ) ...159 Figure 5.10. Effect o f thapsigargin and DBHQ on PDB-mediated signal
transduction ( II) ...160 Figure 5.11. Reduction o f bombesin induced Ca^+ m obüisation by thapsigargin
and DBHQ... 162 Figure 5.12. The mean increase in [Ca^+]^ induced by bombesin after
pretreatm ent w ith thapsigargin or DBHQ...163 Figure 5.13. Time course o f depletion o f bombesin induced calcium response
by thapsigargin and DBHQ...164 Figure 5.14. Effect o f Thapsigargin and DBHQ on tota l intraceUuIar calcium
measured by 4sca^+... 166 Figure 5.15. Effect o f extraceUuIar calcium on [^H]thymidine incorporation
induced by d ifferent m ito g e n s... 169 Figure 5.16. Effect o f econozole on [^Hjthym idine incorporation induced by
d ifferent m ito g e n s ... 170 Figure 5.17. Effect o f extraceUuIar calcium on [^Hjthym idine incorporation
induced by bom besin... 171 Figure 6.1. The bombesin/GRP receptor couples to m u ltip le signal
12
L
is t
o f
T
ables
Table 1.1. Stim ulation o f Swiss 3T3 fibroblast DNA synthesis by various fa c to rs ... 20
Table 1.11. Examples o f G protein coupled receptors and th e ir heterogeneity...25
Table 1.111 Examples o f heterotrim eric G pro te in s u b u n its ...29
Table I.IV. Function o f py subunits... 33
Table l.V . PKC isoform s in mammalian tissues... 51
Table 3.1. Seven transmembrane receptors that couple to m u ltip le G p ro te in s ....126
Table 3.11. Comparison o f receptor-hgand EC50 values between diffe re n t Ga mediated signals...127
Table 3.111. Comparison o f EC50 values o f cellular effects induced by b o m b e sin ....128
Table 5.1. Effect o f DBHQ on DNA synthesis induced by various m ito g e n s...153
13
A
b b r e v ia t io n s
5-HT 80K/MARCKS AA Ach ADP aPKC AR ATP PARK BSA [Ca2+]^„f CaM CaMK cAMP GIF cPKC CRE CREB CTX DBHQ
D-Fs-Phe®, D-Ala^i-B omb esin( 6-13 )OMe DMEM
DMSO DTT
ECso
E D 5 0
EDTA EGF EGTA ER ERK FAK FBS S-hydroxytryptamine (serotonin)
80K/m yristoylated alanine ric h C-klnase substrate arachidonic acid
acetylcholine
adenosine diphosphate atypical p rotein kinase C
adrenergic receptor e.g. ai-AR : a i adrenergic receptor adenosine triphosphate
p-adrenergic receptor kinase bovine serum albumin
concentration o f cytoplasmic Ca^+ concentration o f extracellular Ca^+ calm odulin
calmodulin kinase
cychc adensoine 5' monophosphate calcium in flu x factor
classical/conventional protein kinase C cAMP response element
cAMP response element binding p ro te in cholera toxin
di- ferf-butyUiydroquinone
D-Fs-Phe-Gln-Trp-Ala-Val-D-Ala-His-Leu-OMe
Dulbecco's m odified Eagle's m edium dim ethylsulphoxide
d ith io th re ito l
concentration that gives h a lf m axim um effect dose that gives h a lf m axhnium effect
ethylenediamine tetraacetic acid epidermal grow th factor
ethylene glycol-bis[p-aminoethyl ether]-N,N,N',N',-tetraacetic acid
endoplasmic reticulum
extracellular signal regulated p rotein kinase focal adhesion kinase
14
FGF FDGF Fura-2/AME G pro te in GAP GDP GEF GPCR GRP GST GTP HEPES IBMX
I C 5 0
Ins
In s (l,3,4,5)P4 In s (l,4)P2 In s (l,4,5)P3 IPTG M-Ach receptor m Ab MAP-2 MAPK MBP MEK NGF NmB NmC nPKC PBS PDB PDGF PH PI3K pn>2 PKA
fibroblast grow th factor
fibroblast-derived growth factor fu ra-2 acetoxymethylester
guanine nucleotide binding p rotein GTPase activating protein
guanosine diphosphate
guanine nucleotide exchange factor G protein-coupled receptor
gastrin releasing peptide glutathione-S-transferase guanosine triphosphate
N-[2-hydroxyethyl]piperazine-N'-[2ethanesulfonic acid] isobutyhnethylxanthine
concentration that gives h a lf m axim um in h ib itio n in ositol
in o sito l 1,3,4,5-tetrakisphosphate in o sito l 1,4-bisphosphate
in o sito l 1,4,5-trisphosphate isopropylthio- p-D-galactoside
muscarinic acetylcholine receptor e.g. M^-Ach receptor : M i muscarine acetylcholine receptor
monoclonal antibody
m icrotubule associated protein 2
m icrotubule associated protein 2 kinase/m itogen activated protein kinase
m yelin basic protein
m itogen activated protein kinase/extraceUular signal regulated pro te in kinase kinase
nerve growth factor neuromedin B neurom edin C
novel/new p ro te in kinase C phosphate buffered saline phorbol 12,13-dibutyrate platelet-derivedgrowth factor pleckstrin hom ology
phosphatidyhnositol 3' kinase
15
PKC p ro te in kinase C
PKD p ro te in kinase D
PLAz phosphohpase A2
PLC phosphohpase C
PMSF phenylm ethylsulfonyl fluorid e
PS phosphatidyl serine
PTX pertussis toxin
SCLC smaU ceh lung cancer
SDS sodium dodecyl sulphate
SDS-PAGE SDS-polyacrylamide gel electrophoresis
SE standard error
SERCA sarcoplasmic or endoplasmic re ticu lu m Ca^+ ATPase
SIE sis inducible element
SOS son o f sevenless
SRE serum response element
SH2 src hom ology 2
SH3 src homology 3
TG thapsigargin
TPA tetradecanoyl phorbol acetate
TSH th yro id stim ulating hormone
16
Chapter I
Chapter 1_______________________INTRODUCTION_____________________________ 17
1 .1 .
G
r o w t h
R
e g u l a t io n
M ulticellu lar organisms have developed h ig h ly efficien t regulatory networks to c o n tro l cell p ro life ra tio n . These involve c e llu la r in te ra ctio n s w ith p o sitive and negative d iffu s ib le m odulators as well as w ith the extracellular m a trix proteins. In fu lly m ature organisms, the cells o f many tissues and organs are m aintained in a non- prohferating state (the Gq/Gi phase o f the cell cycle), b u t can be stim ulated to resume DNA synthesis and cell division in response to external stim uh such as hormones, antigens or grow th factors. In this manner the grow th o f in d ivid u a l cells is regulated according to the needs o f the whole organism . The re g u la tio n o f n o rm a l cell p r o life ra tio n is th e re fo re ce n tra l to m any p h y s io lo g ic a l processes in c lu d in g embryogenesis, grow th and development, selective cell survival, haemopoeisis, tissue repair and im m une responses.
It has become evident that cultured cancer cells w hich are characterised by unrestra ine d p ro life ra tio n acquire complete or p a rtia l independence o f m itogenic signals in the extracellular environm ent th ro u g h d iffe re n t mechanisms (Cross and Dexter, 1991; W estermark and Heldin, 1991). These include p ro d u c tio n o f grow th factors that act on the same cells that produced them (autocrine loop) or on adjacent cells (paracrine com m unication), alterations in the num ber or structure o f cellular receptors and changes in the activity o f post-receptor signalling pathways that either stim ulate or suppress cell grow th (Sager, 1989; Bishop, 1991). For these reasons the id e n tific a tio n o f the extracellular factors w hich m odulate cell p ro h fe ra tio n and the elu cid a tio n o f the m olecular mechanisms in volve d have emerged as fundam ental problem s in cancer biology.
The observation that animal cells duphcate their DNA during a discrete m terval between cell divisions allowed the cell cycle to be divided in to fo u r classical phases: G% (grow th phase), S (DNA synthetic period), G2 and M (mitosis). G% is the gap between m ito sis and the in itia tio n o f DNA synthesis, and G2 is the perio d between S and M (reviewed by Baserga 1976 and Pardee et al, 1978). It was fo u n d that, in m ost cases, cells respond to sub-optim al grow th conditions by reversibly arresting in the Gi or Go/Gi) phase o f the cell cycle (Pardee, 1974; Baserga, 1976). This arrest is observed in n orm al fibroblasts in culture (Pardee, 1974). Since the crucial control events fo r the re g u la tio n o f g ro w th seem to reside in Gi (Pardee, et a l, 1978; Pardee, 1989), elucidation o f the mechanisms leading to re stim u la tio n o f DNA synthesis is therefore a necessary step in the understanding o f the control o f cell p ro life ra tio n in vivo.
Chapter 1_______________________INTRODUCTION_____________________________ 18
p ro life ra te and became quiescent in Gi/Gq when they depleted serum o f its grow th p ro m o tin g activity. Furthermore, add ition o f fresh serum caused the cells to reinitiate a program o f RNA and p ro te in synthesis, DNA synthesis and ceU division (reviewed in Pardee, et a l, 1978). Many viraUy and chem ically tran sform ed cells were fo u n d to have either p a rtia lly or w h olly lost the abihty to arrest in G% and exhibited a decreased dependence on serum fo r prohferation. These fin d in g s led to the hypothesis that anim a l sera contained "g ro w th fa c to rs " th a t c o n tro lle d c e llu la r p ro life ra tio n (Wolstenholme and Knight, 1971; Temin, et a l, 1972; Pardee and Rozengurt, 1975).
1 .1 .1 .
G
r o w t h f a c t o r sW ith in the last decade rapid progress has been made toward the elucidation o f the mechanisms o f action o f the diverse extracellular factors that co n tro l the grow th o f ceUs. The availability o f cell culture in n u trie n t media together w ith the p u rific a tio n o f various polypeptide grow th factors (reviewed in Rozengurt, 1980; Rozengurt and Collins, 1983; James and Bradshaw, 1984) allowed the investigation o f m itogen action under chemically defined conditions (Bottenstein, et a l, 1979; Shipley and Ham, 1981). From this w o rk it has become clear that a variety o f m itogenic factors can regulate the p ro life ra tio n o f norm al cells. In th is respect the Swiss 3T3 fib ro b la st has proved a p a rticu la rly useful system fo r id e n tific a tio n o f grow th p ro m oting factors (Rozengurt, 1980; Rozengurt, 1986).
Swiss 3T3 cells are a m u rin e fib ro b la s t cell lin e th a t was established by repeated sub-culture o f disaggregated em bryonic cells (Todaro and Green, 1963). Cultures o f this 3T3 cell hne become "quiescent" in the Gi to Gq phase o f the ceU cycle when the cultures reach confluence at a saturation density determ ined by the serum concentration (HoUey, 1975). This arrest appears therefore to be due to depletion o f g ro w th p ro m o tin g a c tiv ity in the serum. R eaddition o f fresh serum to quiescent cultures o f these ceUs stim ulates ceUular metabohsm and after a variable lag period (10 - 15 hours) in itia tio n o f DNA synthesis.
The use o f Swiss 3T3 ceUs in culture has a num ber o f advantages fo r the study o f grow th control. Identical monolayer cultures o f a clonal p op ula tion o f ceUs can be easUy and consistently produced, aUowing fo r re producibih ty o f results. In a d d itio n a chemicaUy-defined n u trie n t media has been devised fo r these ceUs th a t allows large and reproducible increases in DNA synthesis to be m o n ito re d in the presence o f grow th prom oting agents. Also, since m ost ceUs are arrested in the same phase o f the ceU cycle ( Gi/ G q), on re s tim u la tio n they may be regarded as a synchronised cell
Chapter 1_______________________INTRODUCTION_____________________________ 19
It should be noted, however, that estabhshed cell lines often display abnormal karyotypes and therefore cannot be d irectly correlated w ith any ceh type in vivo. In this respect, the 3T3 cell can be considered as a general m odel fo r grow th control, b u t re s u lts obtained fo rm th is cell lin e necessarily require fu rth e r in v e s tig a tio n to determ ine th e ir relevance in vivo. However, m any mechanisms o f g ro w th c o n tro l id e n tifie d initiaU y in 3T3 ceUs have been demonstrated to be o f general im portance. For example, monovalent io n transport across the plasma membrane (Rozengurt and Heppel, 1975) has since been accepted as a universahy im p o rta n t part o f grow th factor action (Moolenaar, et a l, 1983).
The 3T3 ceh system has proved p a rtic u la rly u se fu l in tw o m a jo r areas o f grow th control research. The firs t is in the id e n tifica tio n o f agents that m odulate the grow th state o f the ceh, and th e ir p u rific a tio n fro m various biological sources. The second js the elucidation o f the mechanisms by which serum, p u rifie d grow th factors and other m itogens in itia te DNA synthesis. A tte n tio n has been focused on the early signalhng events in mitogenesis as the in itia l steps in the prohferative response.
It is now recognised that the p ro h fe ra tio n o f 3T3 ceUs may be regulated by several classes o f m itogen (table 1.1). These m clude members o f a large famQy o f polypep tide grow th factors such as PDGF, EGF and insulin-hke g row th factors and m embers o f a fa m ily o f smaU regulatory peptides that classicahy behave as local horm ones or fast acting neuro transm itte rs. In a d d itio n various pharm acological agents in clu d in g phorbol esters and synthetic diacylglycerols can act as m itogens fo r these cehs (table 1.1).
Both the polypeptide grow th factors and the neuropeptides stim ulate th e ir grow th p ro m oting effects by binding to specific receptors located on the ceh surface and activating m u ltip le signahing pathways. A t least two m a jo r signal tran sduction pathways in itia te the cascades o f molecular events stim ulated by these two classes o f mitogens. The polypeptide grow th factors e.g. PDGF and EGF b in d to receptors w ith in trin s ic tyrosine kinase a c tiv ity w hile the neuropeptides act th ro u g h receptors coupled to heterotrim eric guanine nucleotide binding proteins (G proteins) to effector activation.
Chapter 1 INTRODUCTION 20
Table l.I. Stim ulation of Swiss 3T3 fibroblast DNA synthesis by various factors
Class of Growth Factor Name of Factor Reference
Polypeptide growth factors Epidermal Growth Factor Insulin-like Growth Factors Platelet Derived Growth Factor
Carpenter and Cohen, 1979 Froesch, et al, 1979
Rozengurt, et al, 1983c; Heldin and Westermark, 1984; Lopez- Rivas, etal, 1984
Neuropeptides Adrenomedullin Bombesin Bradykmin EndotheUn Vasopressin Vasoactive Intestinal Contractor Vasoactive Intestinal Peptide
Activators of Protein kinase C Phorbol esters, teleocidin, 1 -Oleoyl- 2 -acetylgly cerol
Cychc nucleotide elevating agents
Cholera toxin. Adenosine agonists, cAMP derivatives, Forskolin, IBMX,
Prostaglandin E^
Withers, etal, 1996
Rozengurt and Sirmett-Smith, 1983
WoU and Rozengurt, 1988b Brown and Littlewood, 1989;
Takuwa, etal, 1989
Rozengurt, et al, 1979; Dicker and Rozengurt, 1980;
Rozengurt and Mendoza, 1980; Rozengurt, etal, 1981a Fabregat and Rozengurt, 1990 Zurier, etal, 1988
Dicker and Rozengurt, 1978; Dicker and Rozengurt, 1980; Dicker and Rozengurt, 1981; Collins and Rozengurt, 1982a; Collins and Rozengurt, 1982b; Rozengurt, etal, 1984
Rozengurt, 1981; Rozengurt, et al, 1981b; Rozengurt, 1982b; Rozengurt, 1982a; Rozengurt, et al, 1983a; Rozengurt, etal,
1983c Microtubule disrupting agents Colchicine, Colemid,
Nocodazole, Podolotoxin, Vinblastine
Friedkin and Rozengurt, 1981; Wang and Rozengurt, 1983
Vitamin A derivatives
Permeabihty modulators
Retinoic acid
Mellitin, Mastoparan
Dicker and Rozengurt, 1979; Dicker and Rozengurt, 1980) Gelehrter and Rozengurt, 1980;
Chapter 1_______________________INTRODUCTION_____________________________ 21
1 .1 .2 .
S
y n e r g is t icS
ig n a l u n gP
a t h w a y sThe use o f Swiss 3T3 cehs as a m odel fo r the Investigation o f grow th factors has revealed the fundam ental concept o f synergistic signal tra n sd u ctio n pathways. D efined mitogens such as those summarised in table 1.1 all exhibit potent synergistic in te ra ctio n s when added to cell cultures m aintained in m ediu m devoid o f serum (reviewed in Rozengurt, 1986). In contrast to m any other m itogens PDGF and bom besin stim ulate DNA synthesis and cell d ivisio n in the absence o f any other synergistic factor. Studies w ith PDGF id e n tifie d that this grow th facto r activates PKC (Rozengurt, et a l, 1983b) and causes elevation o f cAMP (Rozengurt, et a l, 1983c). It was predicted that i f an agent ehcits the same set o f early events as PDGF it should act as a grow th factor fo r Swiss 3T3 cells in m edium devoid o f serum or other mitogens. Agents th a t e licit p a rt o f the early responses stim ulated by PDGF should become m itogen ic when added in com binations th a t re co n stitu te the other early events (Rozengurt, 1986). The availability o f a panel o f defined mitogens that are biologically active (see table 1.1) provides a tool fo r elucidating the nature o f the regulatory signals and m olecular events im phcated in these synergistic effects (reviewed in Rozengurt, 1986). For example, a group o f agents such as p h o rb o l esters, diacylglycerol and vasopressin elicit a common set o f early events; namely they activate PKC b u t do not alter the basal level o f cAMP. A d d itio n o f any o f these agents either in d iv id u a lly or in com bination to Swiss 3T3 cells fails to induce a m itogenic response. Agents that increase in tra c e llu la r cAMP such as p ro sta g la n d in Ei, cholera to x in and cAMP derivatives do n o t activate PKC and do n o t stim ulate DNA synthesis when added singly or in com bination. Therefore agents that share a com mon signalling system cannot act synergisticaUy to stim ulate in itia tio n o f DNA synthesis.
Crucially the agents m entioned above become potent mitogens when added to quiescent Swiss 3T3 cells in com binations that ehcit the generation o f b o th types o f signals and thereby reconstitute the complex p attern o f signalling events elicited by PDGF. In s u lin w h ich can synergise w ith b o th groups o f extra cellula r fa cto rs at supramaximal concentrations (that is in hen o f insulin-like grow th factor) does n o t act id e n tica lly to either group o f agents. In fact th is horm one does n o t activate PKC, increase in tracellular Ca^+ or increase the level o f cAMP in in tact 3T3 cells (Rozengurt, 1986). Similar conclusions can be drawn when EGF is added instead o f insulin.
Chapter 1_______________________INTRODUCTION_____________________________ 22
1 .1 .3 .
REGULATORY SIGNALS AND OBLIGATORY EVENTS
In line w ith the hypothesis o f grow th control discussed above the key events elicited by grow th factors in quiescent cells can be b road ly divided in to two m ajor categories: regulatory signals and obhgatory events (Rozengurt, 1986). The fo rm e r class represent in tra ce llu la r processes th a t mediate the action o f specific growth- p ro m o tin g agents; although they are crucial in ehciting biological responses by a given fa cto r they can be bypassed by another group o f factors. In contrast, oblig a to ry events are envisaged as m olecular steps that m ust take place fo r the s tim u la tio n o f DNA synthesis and ceU division to occur regardless o f the regulatory signals utihsed to activate the cells. This d is tin c tio n is p a rtic u la rly relevant to section 1.3.2.5. In order to distinguish between regulatory signals and obhgatory events one approach is to ascertain the effect o f a wide panel o f defined m itogenic agents. W hile regulatory signals are elicited by certain mitogens b u t n o t others, obhgatory events should be stim ulated by ah m itogenic combinations. This principle is utihsed in section 5.5.
The next section iUustrates the mechanism by w hich receptors w ith in trin s ic tyrosine kinase activity can ehcit m u ltip le signals; this is due to their m u ltip le docking sites fo r signal transduction molecules. The mechanism by w hich G protein-coupled receptors can ehcit m u ltip le signal transduction pathways is more elusive. This aspect o f m itogenic signalhng w h l be one o f the m ajor subjects o f the present thesis. In particular, because bombesin, hke PDGF, is a sole m itogen fo r Swiss 3T3 cells m uch atte n tio n has been given to elucidating its cellular mechanisms o f activation. This thesis wiU describe signalhng pathways m ediated by the bombesin/GRP receptor w h ich is a seven transmembrane, G p ro te in coupled receptor. However, at poin ts thro ugho ut this thesis G protein-coupled receptor signalhng shall be compared and contrasted to pathways ehcited by receptor tyrosine kinases. The sahent p o in ts o f receptors w ith in trin s ic tyrosine kinase a ctivity shah be b rie fly described fir s t and then G protein-coupled receptors shah be described in more detah.
1 .1 .4 .
RECEPTOR TYROSINE KINASES
Receptor tyrosine kinases b in d large polypeptide grow th factors such as PDGF and EGF. PDGF is a 30 kDa homo- or heterodim er o f A or B chains (reviewed in Ross,
Chapter 1_______________________INTRODUCTION_____________________________ 23
PDGF as a grow th factor is hlghhghted by the v-sis oncogene fro m sim ian sarcoma virus w hich is homologous to the PDGF-B chain. The inappropriate p ro d u ctio n o f v-sis
b y in fe c te d cells m ay re su lt in autocrine or paracrine s tim u la tio n w h ich could c o n trib u te to the tum ourigenesis observed w ith th is oncogene (W estermark and Heldin, 1991). Unlike PDGF, EGF is ubiquitous being fo u n d in nearly all body flu id s and the EGF receptor is fo u n d in m any cell types (Carpenter, 1985). The v-erbB
oncogene fro m avian erythroblastosis virus is a truncated EGF receptor w h ich lacks p a rt o f the EGF regulated extracellular domain (Downward, et a l, 1984).
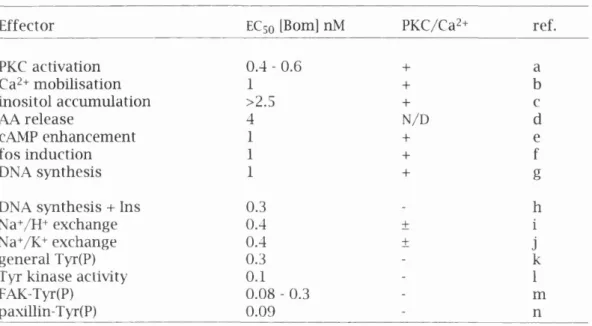
Receptors fo r EGF and PDGF contain a large extra cellula r lig a n d b in d in g domain, a single transmembrane region and a cytoplasmic p o rtio n w ith a conserved p ro te in tyrosine kinase dom ain (reviewed in Fantl, et a l, 1993; Malarkey, et a l, 1995). Upon ligan d b in d in g the receptor chains dimerise and th is activates th e ir in trin s ic protein tyrosine kinase activity. The active receptor autophosphorylates m u ltip le sites by an interm olecular mechanism. The tyrosine residues that become phosphorylated serve as docking sites fo r m u ltip le signal transduction molecules. This has been best characterised fo r the PDGFp receptor. A series o f m uta tions m apped the sites at w hich the d iffe re n t molecules interact w ith the in tracellular p o rtio n o f this receptor, see figure 1.1.
The PDGF receptor binds Src, p85, SHPTP-2 Grb2, GAP and PLCy. Some o f these proteins have enzymatic activity such as PLCy and Src, w h ilst others appear to have no enzym atic a ctivity b u t instead serve as linkers or adapters between receptor tyrosine kinases and enzymes, e.g. p85 and Grb2. It is possible that these associations sim ply serve to localise the s ig n a llin g m olecules to sites near th e ir substrates e.g. phosphohpids in membranes are hydrolysed by PLCy or phosphorylated by PI3K. In a d d itio n phosphorylation o f substrates may m o d ify the conform ation and a ctivity o f these m olecules (Malarkey, et a l , 1995). It has been show n th a t ty ro s in e phosphorylation o f PLCy regulates its enzymatic activity (Lee and Rhee, 1995) and see
section 1.3.1.
Chapter 1 INTRODUCTION 24
like SH2 domains there is specificity between proline rich sequences and SH3 domains determined by variable amino acids that surround the conserved m o tif (Cohen, et al,
1995). The adaptor molecules Grb2 is composed of several of these modules, SH3— SH2—SH3 (Lowenstein, et al, 1992).
PDGF receptor
extracellular intracellular
I
Y1009
PTP-21P
V1021
Kinase domain
Phosphotyrosine residues
Figure 1.1. Diagram of PDGF receptor with sites of interaction with
signal transduction molecules.
Based on Malarkey, et al, (1995). For simplicity only one of the chains is shown. The phosphorylation of the receptor occurs upon ligand induced dimérisation (see text for details).
1 .1 .5 .
G P
r o t e i n-C
o u p l e dR
e c e p t o r sChapter 1 INTRODUCTION 25
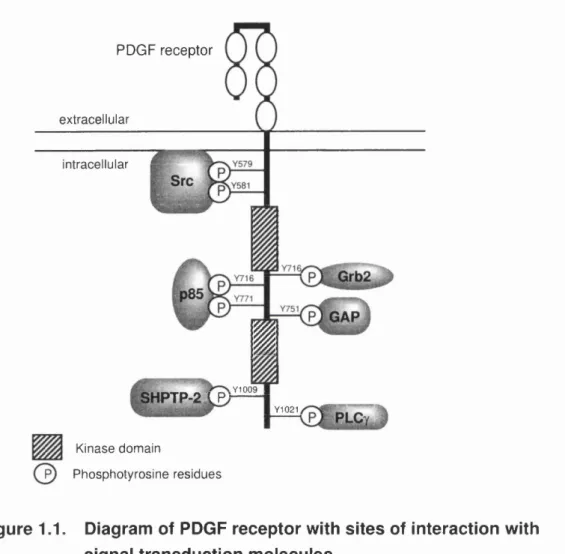
Table l.II. Examples of G protein coupled receptors and their heterogeneity
Receptor/hgand Subtype predom inant G p ro te in coupling
m uscarinic acetylcholine M l, M3, M5 Gq/11
M2, M4 Gi/o
adenosine A l, A3 Gq/11
A2A1 A2B Gs
adrenaline/noradrenaline aiA, aiB, o^iD Gq/11
Ct2A. «28» «2C Gq/11
Pi. P2, P3 Gs
angiotensin A T i Gq/11
AT2 >lcGMP
bombesin GRP (BB2), NmB (BBi), BRS3 Gq/11
bradykm in Bi, B2 Gq/11
cholecystokinin/gastrin CCKa, CCKb Gq/11
dopamine D i, D5 Gs
Dz, D3, D4 Gq/11
endothehn ETa, ETb Gq/11
glutamate m G lui, mGlus Gq/111
mGlu2, mGlu3, mGlu4, mGl%, m Glu; Gq/n
histam ine H i Gq/11
H2 Gs
serotonin 5-HTia, 5-HTib, 5-HTid, 5-HT IE, 5-H Tif Gq/11 5-HT2A, 5-HT2B. 5-HT2C Gq/11
5-HT4, S-HTe, 5-HT7 Gs
prostanoid DP, IP, EP2, EP4 Gs
FP, TP, EPi, EP3 Gq/11
EP3 (alternatively sphced) Gi/o
som atostatin SSti, SSt2, SSt3, SSt4, SStg Gi/o
NK1 - Substance P receptor, NK2 - Substance K receptor, NK3 - neurokinin B receptor. Table
based on TIPS receptor and ion channel nomenclature supplement 1995.
1 .1 5 .1 . GROWTH REGULATION BY G PROTEIN-COUPLED RECEPTORS
Chapter 1_______________________INTRODUCTION_____________________________ 26
Rozengurt, 1991b; Sethi and Rozengurt, 1991a; Sethi and Rozengurt, 1992; Sethi , et a l, 1993). It is know n th a t GRP, vasopressin, cholecystokinin and neurotensin are secreted by some SCLC tum ours (for refs, see Sethi et. a l 1992). Other peptides may be released by a variety o f norm al cells in the lung, or lik e b ra d ykin in , produced e xtra ce llu la rly as a re su lt o f the p ro te o ly tic cleavage o f plasma precursors in the damaged tissue su rro unding tum o urs (Steranka, et a l, 1989). C o llectively these fin d in g s su p p o rt the hypothesis th a t SCLC g ro w th is sustained b y an extensive ne tw o rk o f autocrine and paracrine in teractio ns in vo lvin g m u ltip le neuropeptides w hich signal thro ugh G protein-coupled receptors (WoU and Rozengurt, 1988a; Sethi,
et a l, 1992).
In add ition , receptors that are coupled to G p rotein s have been shown to tra n s fo rm fib ro b la sts in vitro. The serotonin receptor 5-HTic was shown to be a co n d ition al oncogene by having agonist-dependent tran sform ing p o te n tia l (Juhus, et a l, 1989) and this was also shown fo r certain m uscarinic receptors (Gutkind, et a l,
1991). In other studies m utated «ig-AR receptors which were co n stitutively activated and hgand independent were transform ing (AUen, et a l, 1991) and activated m utants o f the TSH receptor (Parma, et a l, 1993) were hnked to th y ro id hyperplasia. Thus the study o f cellular events ehcited by G protein -cou pled receptors is im p o rta n t fo r understanding neoplastic growth.
I.I.5 .2 . STRUCTURE
The m ain characteristic o f G protein-coupled receptors is seven hydrophobic regions o f approxim ately 23 - 28 amino acids and the p erio dic d is trib u tio n o f the h yd ro p h o b ic ity is consistent w ith an a-hehcal conform ation. These hehces, w hich contain the areas o f highest homology, are postulated to span the plasma membrane and are connected by hydrophihc extraceUular and intraceUular loops th a t are less weU conserved (Strader, et a l, 1995), see figure 1.2.
The arrangement o f the hehces w ith in the membrane is modeUed u p o n the pro je ctio n map o f rhodopsin (Schertler, et a l, 1993) and the probable order deduced fro m the chemical and physical prope rties o f conserved features o f a ll G -protein lin ke d receptors sequenced at that tim e (Baldwin, 1993).
Chapter 1 INTRODUCTION 27
N - terminal 1st 2nd 3rd extracellular loops
intracellular loops 1 st 2nd
C - terminal
Figure 1.2. Schematic diagram of a seven transmembrane G-protein
coupled receptor
Figure 1.3. Probable arrangement of helices in seven transmembrane
domain receptors.
Viewed from the extracellular surface and modelled on rhodopsin projection map. The shaded area between helices I, II and III represents these helices lying at a slant in the membrane.
Biogenic amines b in d w ith in the IV - VII pocket. For p ep tid e and p ro te in ligands helix II and III, and the extracellular domains are used as well. For example the C -term inal o f substance P binds in the IV - VII pocket and the N -term inal lays across the slanted heUx II (Strader, et a l, 1995).
C hapter 1_______________________INTRODUCTION_____________________________ 28
synthetic peptides derived fro m the C-term inal o f the 3rd cytoplasm ic dom ain are capable o f e fficie n tly stim ulating G-proteins in vitro (Lefkowitz, et a l, 1993). Thus the receptor m ust shield these peptides fro m the G p ro te in when in an inactive, hgand free conform ation.
The other m ain feature o f G protein-coupled receptors is th a t they have no enzym atic a c tiv ity themselves, u n like the receptor tyrosine kinases, b u t they are nucleotide exchange factors fo r heterotrim eric G proteins.
1 .1 .6 .
HETEROTRIMERIC G-PROTEINS
Seven transmembrane receptors are coupled to heterotrim e ric G-proteins that are com prised o f an a, a p and a y subunit. The p and y subunits can only be separated u p o n d é n a tu ra tio n and so they are generally considered a fu n c tio n a l m onom er. Molecular cloning has id e n tifie d 20 a subunits, 5 p subunits and 12 y subunits to date. See table l. I I l fo r examples o f diversity and tissue distribu tion.
i.1 .6 .1 . STRUCTURE
The structure o f heterotrim eric G proteins has been reviewed by Neer (1995). Ga are proteins o f 39 - 50 kDa and consist o f a GTPase dom ain and a largely a-hehcal dom ain. Areas where the m ost co n fo rm a tio n a l change occurs u p o n n u cleotid e exchange are called switch regions and this is where the p subunit binds to Ga. The receptor binds to the C-terminal end o f Ga.
Gp are approxim ately 35 kDa and are members o f the fa m ily o f WD-repeat p ro te in s (Neer, et a l, 1994) characterised by 4 - 8 re cu rrin g h ig h ly conserved u n its ending w ith Trp-Asp (WD). In p subunits the length o f the repeat is about 40 amino acids and so is sometimes know n as the WD-40 repeat. Very recently the crystal structure o f p has been described (Wah, et a l, 1995; Lam bright, et a l, 1996; Sondek, et a l, 1996). The p subunit resembles a 7-bladed propeller w ith each blade made o f 4 antiparallel strands in a p-sheet. The WD-40 repeat comprises the outer strand o f one p-sheet together w ith the 3 inner strands o f the next p-sheet. The m ost variable sequences in the repeats are found in the outer strand and this is consistent w ith the proposal that WD proteins b u ild m u ltip ro te in complexes (Neer, et a l, 1994). A t the N- te rm in a l is an a-helix w hich form s a coded cod w ith the N -term inal a-hehx o f the y subunit.
Gy is smad, about 6 - 9 kDa and shows the m ost heterogeneity o f the G pro te in subunits and thus has been postulated to determine the specificity o f Py com binations (Neer, 1995). Not ad py combinations can occur. For example pi form s dimers w ith yi
Chapter 1 INTRODUCTION 29
or 72 (Iniguez-U uhi, et a l, 1992; Pronin and Gautam, 1992; Schmidt, et a l, 1992), and see section 1.1.6.4.
Table l .I II Examples of heterotrim eric G protein subunits
Subunit Toxin Distribution Typical receptor Effector
‘^s(s) “ s(l) « o l f
CTX ubiquitous
olfactory neuroepithelium
p-AR, TSH
odorant
stimulate adenylyl cyclase
G l ttQ
«12
«13 ® oA “ oB
t t f i
ttt2 “ g ttz PTX CTX, PTX CTX, PTX CTX(?), PTX nearly ubiquitous ubiquitous nearly ubiquitous brain, others brain, others retinal rods retinal cones taste buds
brain, adrenal, platelets
M2-Ach, tt2-AR tt2-AR rhodopsin cone opsin taste (?) M2-Ach
inhibit adenylyl cyclase
stimulate cGMP phosphodiesterase
?
inhibit adenylyl cyclase
Cfq Ctq “ 11 “ 14 “ 16 “ 15 nearly ubiquitous nearly ubiquitous lung, hver, kidney T cehs, myeloid ceUs B cehs, myeloid cehs
MpAch, ttj-AR ? ? ? stimulate PLCp ?
Gi2 ai2
“ 13 ubiquitous ubiquitous ? ? ? ?
P Pi
P2
P3 P4
ubiquitous nearly ubiquitous
Y Yl
Y2 Y3 Y4 Ys Y6 Yz
retina, other (?) brain, adrenal, other (?) brain, testis
kidney, retina (?) widely expressed brain, other (?) widely expressed
Based on Hepler and Gilman (1992), additional information from Cali, etal. (1992)
I.I.6 .2 . THE G PROTEIN CYCLE
Chapter 1 INTRODUCTION 30
change in the switch region of Ga at the Py b in din g site and causes dissociation o f the com plex in to Ga, GPy and receptor. The crystal structure o f py does n ot change when Ga is dissociated (Lam bright, et a l, 1996; Sondek, et a l, 1996). It has been proposed that the a ctivity o f Py is due to the active bin din g site being exposed (Clapham, 1996). Ga has in trin s ic GTPase a c tiv ity and is therefore deactivated a fte r a certain tim e. Phospholipase C can enhance the in trin s ic GTPase a c tiv ity o f Gaq (Berstein, et a l,
1992). When GTP is hydrolysed to GDP, Ga reassociates w ith py and the receptor. Thus the a ctivity o f b o th a and Py is lim ite d by the hydrolysis o f GTP (see figure 1.4) (Neer, 1995; Hamm and G ilchrist, 1996).
ligand
GTP GDP
Figure 1.4. G protein cycle.
Chapter 1_______________________INTRODUCTION_____________________________ 31
The assembled receptor-G protein complex results in a high a ffin ity state o f the receptor fo r its ligand. However, when the G p ro te in in its GTP b o u n d state has dissociated, the receptor agonist complex is less stable and the receptor changes to a low er a ffin ity state. Thus, in membrane preparations in the presence o f GTP there is a decrease in the a ffin ity o f the receptor fo r its hgand (Spiegel, 1988). Use has been m ade use o f th is observation to id e n tify receptor coupling to G pro te in s (e.g. see Sinnett-Smith, et a l, 1990 and section 1.2.1).
1 .1 .6 .3 . CLASSES OF G PROTEIN
A lth ough Py transduce signals also (Clapham and Neer, 1993) (see below) it is the a subunit which determines the functiona l class o f G protein. These fa ll in to fo u r groups (see table l.lll) : Gg because they stim ulate adenylyl cyclase, Gi because they in h ib it adenylyl cyclase, Gq which activates PLCp and G%2 fo r w hich the direct effector is unkn ow n although recent w o rk has im phcated the G%2 fam üy in Na+/H+ exchange and stress fib re assembly (Dhanasekaran, et a l, 1994; Voyno-Yasenetskaya, et a l ,
1994a; Buhl, et al, 1995). Ga regulation o f these effectors wiU be described in la ter sections.
It was o riginally thought that receptors only coupled to one species o f Ga b u t this now appears n ot to be the case. This w ü l be discussed fu rth e r in Chapter 3.
1 .1 .6 .4 . Pys u b u n it s
Chapter 1_______________________INTRODUCTION_____________________________ 32
the am ino term in al p o rtio n o f the PH dom ain binds specifically to PIP2 (Harlan, et a l,
1994) in keeping w ith the m a jo rity o f PH dom ain-containing p rotein s being fo u n d locahsed to cellular membranes.
It was m entioned above that n o t all py com binations can occur. This could indicate that specific py combinations are required fo r coupling to distin ct receptors or fo r activating d is tin c t effectors. D uring th is thesis aspects o f py signalling shall be discussed, therefore some evidence regarding Py specificity shall be presented here.
A series o f experiments in which selected endogenous subunits were targeted by m icro in je ctio n o f antisense oligonucleotides revealed some degree o f specificity. It was shown that in neuronal cells the M4 m uscarinic receptor couples to GaoiPsyé whereas a som atostatin receptor couples to Gao2Piy3 (Kleuss, et a l, 1991; Kleuss, et a l,
1992; Kleuss, et a l, 1993). More recently Yan, et a l (1996) have used the yeast two h y b rid system to show diffe re n tia l abüity o f p and y subunits to fo rm the G p ro te in Py
complex. A ll isoform s were tested (prPs and yi-ys andy?) and i t was dem onstrated that
Pi and p2 can fo rm tig h t complexes w ith m ost y except (yi and y2) and that there is wide heterogeneity between the other combinations. This shows that there could be specific Py com binations b u t does not reveal anything about the selection o f specific signalling pathways.
The effect o f py subunits on various enzymes and signalling pathways has been the subject o f many studies. Table l.IV shows some events which specific Py subunits regulate. It should be noted that n ot all Py com binations have been assessed. In general the com binations w hich have been tested so far show sim ilar activities. Some
py com binations appear to show specificity: Piyi, the re tin a l py fo r Gat (Iniguez-Uuhi,
et a l, 1992; Ueda, et a l, 1994) is recognised to be d iffe re n t (see Clapham and Neer, 1993) and P2yz w hich does all the signalling described fo r other Py subunits except it does n o t activate the tyrosine kinases, Tsk and Btk. Some aspects o f specificity have been shown in other ways. L u ttre ll, et a l (1995) looked at the effect o f several PH
Chapter 1 INTRODUCTION 33
Table l.IV . Function of py subunits
e x is t P T X A C PLC M A P K she T s k B tk a c tin
P i Yl + -/+
4-72 + 4- 4- 4- 4- 4- 4-
4-73 + + 4- 4- 4- 4-
4-74
-1-75 -1- + 4-
4-77 + + 4-
4-P2 7 i - - - -
-72 -1- 4- 4- 4- 4- 4- -
-73 -1- 4- 4- 4- 4- 4-
4-75 -1- 4- 4-
4-77 + 4- 4- +
Pb 7 i - -
-72 - -
-73
P4 7 i -
-72 -
-73
refs. a b C d e f g h 1
The table shows a summary of the documented effects of Py subunits, (+) denotes an effect, (-)
denotes no effect, exist: not all py combinations can exist. (+) shows that they form a physical
dimer and (-) shows they do not; PTX: enhancement of pertussis toxin ADP ribosylation of Gj
or Goi AC: regulation of adenylyl cyclase. This includes both inhibition and stimulation of the
relevant subtypes as described in the introduction, both effects have been grouped; FLO:
activation of PLCp isoforms; MARK: activation of MAP kinase; she: tyrosine phosphorylation
of she; Tsk: activation of Tsk; Btk: activation of Btk; actin: formation of stress fibres, refs.: a
(Iniguez-Uuhi, etal., 1992; Kleuss, etal., 1992; Pronin and Gautam, 1992; Schmidt, etal.,
1992; Kleuss, etal., 1993); b (Ueda, etal., 1994); c (Iniguez-Uuhi, etal., 1992; Ueda, etal.,
1994); d (Ueda, etal., 1994; Hawes, etal., 1995); e (Faure, etal., 1994; Hawes, etal., 1995); f (Touhara, et al., 1995; van Biesen, etal., 1995); g and h (Langhans-Rajasekaran, et al.,
1995); l(Buhl, efa/., 1995).
I.I.6 .5 . MODIFYING TOXINS
Chapter 1_______________________INTRODUCTION_____________________________ 34
to o l is pertussis to x in fro m Bordetella pertussis. This to xin ADP-ribosylates a specific cysteine residue near the C-terminal o f Gi and Go (West, et a l, 1985). Gg and Gq do n ot have the necessary cysteine and are insensitive to pertussis to xin . The p re fe rre d substrate o f pertussis to x in is the trim e ric com plex rath er than ju s t the a subunit w hich means that py is no longer liberated fro m Gj. Thus cholera to x in locks Gg in the GTP state whereas pertussis toxin locks G^ in the GDP state and is used as an in h ib ito r o f Gi (van Dop, et al, 1984a; Spiegel, 1988).
1 .2 .
B
o m b e s in
Bombesin is a bioactive tetradecapeptide th a t was o rig in a lly discovered in extracts fro m the skin o f the European toad Bombina bombina (Anastasi, et a l, 1971) w hich had ple io tro p h ic pharmacological effects in a range o f d iffe re n t mammals (for general review see Tache, et a l, 1988). Bombesin produced a rise in systemic blood pressure and had stim u la n t action on isolated sm ooth muscle preparations o f the in te s tin e , colon, u terus and bladder. When a d m in iste re d c e n tra lly it affe cted therm oregulation, gastric fu n ctio n and behaviour. Bombesin also caused pancreatic secretions o f digestive enzymes (e.g. amylase), in s u lin and glucagon (Tache, et a l,
1988), and the release o f several gut hormones (Ghatei, et a l, 1982) Bombesin-hke im m u n o re a c tiv ity is fo u n d in the brain , spinal cord, in nerve fib re s along the g a s tro in te s tin a l tract, where it is th o u g h t to act as a n e u ro tra n s m itte r, and in neuroendocrine ceUs o f the lung and th y ro id (for refs, see Solcia, et a l, 1988; Sunday, 1988).
Chapter 1_______________________INTRODUCTION_____________________________ 35
Bom besin subfam ily
GRP Ala- Pro- Val- Ser- Val- Gly-Gly-Gly-Thr- Val- Leu-Ala-
Lys-Met-Tyr- Pro-Arg-Gly-Asn- His-Trp-Ala- l/aZ-Gly-Hls-Leu-Met-NHj
GRP-10 or Gly-Asn- His-Trp-Ala- l/aZ-Gly-Hls-Leu-Met-NHj
Neuromedin 0
Bombesin pGlu-Gln-Arg- Leu-Gly-Asn-GIn-Trp-Ala- V'aZ-Gly-Hls-Leu-Met-NHz
LItorIn subfam ily
Litorin pGlu- GIn-Trp-Ala- l/aZ-Gly-Hls-P/ie-Met-NHj
Neuromedin B Gly-Asn-Leu-Trp-Ala-TZir-Gly-Hls-Pfte-Met-NHa
Figure 1.5. Amino acid sequence of mammaiian and amphibian
bombesin-iike peptides
Amino acids in bold show the C-terminal tetrapeptide that identify the subfamily. Amino acids in
italics show where NmB and litorin differ from bombesin and GRP.
Bombesin, lito rin , ranatensin and p h y llito rin are all am phibian peptides. GRP, NmB and NmC are mammahan peptides. GRP and NmC have 9 o f the last 10 amino acids id e n tica l to bom besin and NmC is also know n as GRP-10 or GRPig.27. NmB belongs to the h to rin subfam ily and although the firs t fo und bad 9 amino acids a large NmB o f 32 am ino acids has also been described (M inam ino, et a l, 1988). By s u b m ittin g 27 bom besin-hke peptides to pa ra lle l bioassay it was shown th a t the m in im u m amino acid sequence required fo r bom besin-like a ctivity was the C-term inal beptapeptide, the tryp to p h a n residue being o f crucial im portance. FuU a ctivity was seen w ith the C-terminal nonapeptide (Erspamer, et a l, 1988).
Bombesin im m unoreactivity was found in the endocrine cehs o f the developing hum an lu ng (Wharton, et a l, 1978) and in small-cell lung carcinoma (SCLC) (Moody, et a l, 1981; Wood, et a l, 1981; Erism an, et a l , 1982) a cancer o f p u lm o n a ry neuroendocrine cehs characterised by a ra p id grow th rate, high m etastatic p o te n tia l and ectopic horm one p ro d u c tio n (Greco and Oldham, 1979). T his raised the p o s s ib ility th a t bom besin could particip ate in the c o n tro l o f cell p ro h fe ra tio n and indeed it was shown that bombesin was a sole m itogen fo r Swiss 3T3 cehs in culture (Rozengurt and Siim ett-Sm itb, 1983). This was fo llo w e d b y the observation that bombesin/GRP was a grow th factor fo r norm al bronchial epitbehal cehs (WiUey, et a l,
Chapter 1_______________________INTRODUCTION_____________________________ 36
b om be sin acted as an autocrine g ro w th fa c to r fo r SCLC. In a d d itio n [Leu^^- \|/(CH2NH)Leui4]bombesin, a specific bombesin/GRP receptor antagonist (WoU, et a l,
1988), was shown to in h ib it the g ro w th o f SCLC cell lines in v itro and in h ib it x e n o g ra p h t fo rm a tio n in nude m ice (M ahm oud, et a l, 1991). Since th e n bombesin/GRP has been im phcated in other tum ours such as pancreatic (Avis, et a l,
1993) and gastric tum ours (Qin, et a l, 1994) and has also been shown to be a grow th factor fo r gastric carcinoma ceU lines (Bold, et a l, 1994).
1 .2 .1 .
B
o m b e s in/G R P
r e c e p t o rBom besin/G R P m ediates its e ffe cts th ro u g h s p e c ific re c e p to rs f ir s t characterised on pancreatic acinar ceUs (Jensen, et a l, 1978). The bombesin/GRP re ce p to r in Swiss 3T3 cells is a 75 - 80 kDa cell surface g lyco p ro te in th a t was id e n tifie d b y a ffin ity crosslinking (Zachary and Rozengurt, 1987) w ith a p ro te in core o f 43 kDa (Sinnett-Smith, et a l, 1988) and functionaUy coupled to a GTP bind in g pro te in (Fischer and Schonbrunn, 1988; Sinnett-Smith, et a l, 1990) demonstrated by the ab ility o f GTP or GTP analogues to reduce the a ffin ity o f bom besin fo r its receptor (see se ctio n 1.1.6.2). Two types o f b om be sin re c e p to r were p h a rm a c o lo g ic a lly distinguished in rat oesophagus, a bombesin/GRP preferring receptor (Kj bombesin = 4 nM, Kj NmB = 156 nM) and a NmB preferring receptor (Kj NmB = 0.3 nM, Kj bom besin = 2 nM) (von Schrenck, et a l, 1989). Bombesin and GRP d iffe r fro m NmB by 2 am ino acids in the C -term inal beptapeptide. One o f these differences, the Phe o f NmB compared to the Leu o f bombesin and GRP at the penultim ate residue, is also fo u n d in lito rin (see figure 1.5), b u t lito rin has equal potency to bom besin in sm ooth muscle p reparations and in Swiss 3T3 ceUs (Erspamer, et a l, 1988; MUlar and Rozengurt, 1990b; Sinnett-Sm ith, et a l, 1990). The residue th a t d iffe rs between NmB and bombesin, GRP or h to rin is the Thr o f NmB five residues fro m the C-term inal end, see figure 1.5, and this could be the key residue that confers receptor specific recognition.
![Figure 3.9. Effect of antagonist on bombesin-induced pH]thymidine incorporation.](https://thumb-us.123doks.com/thumbv2/123dok_us/8640629.1430337/110.892.46.766.84.788/figure-effect-antagonist-bombesin-induced-ph-thymidine-incorporation.webp)