A 60-Kilodalton Protein Component of the Counting Factor Complex Regulates Group Size in Dictyostelium discoideum
Full text
Figure

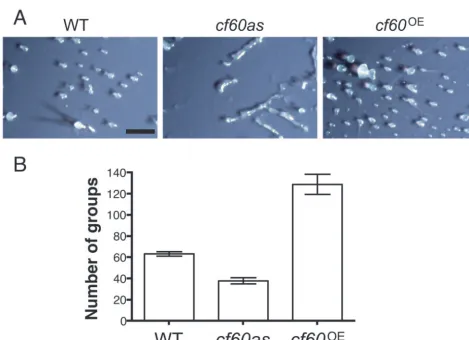
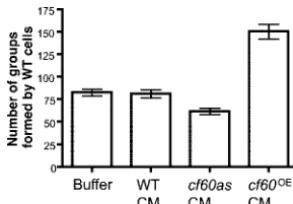
Related documents
Machine learning issued in security related tasks involving classification [7], such as intrusion detection systems [2], spam filters[4], biometric
As the aim of this paper is to examine the impact of foreign direct investment on stock market development in Ghana, we make use of market capitalization as a proportion of GDP,
The main focus of the pension insurance due to Germany’s social laws is to sustain the ability to work and to sustain the population’s earning capacity through medical and
First, we find evidence that both of the variables used to measure relative income (income predicted from a wage equation and a worker's prior expected income) have the
truncatus can successfully complete its development on sorghum grain, with whole grain substrate and high-yield improved sorghum cultivars showing higher
If you don’t know an administrator user password and you have the Mac OS X Install Disc 1 CD, start up your computer using the CD, choose Reset Password from the Installer menu,
In this study, preparation of porous ceramics made from sediment by gelcasting method was tried and the properties of the ceramics filter, such as porosity,
