TIME ON CARBON ASSIMILATION OF RICE PLANT DURING THE RIPENING PERIOD
K. S. MuRTY*
National Institute
of
Agricultural Sciences, TokyoSuMMARY
Investigations were carried out on the effect of top-dressing nitrogen at full heading time on carbon assimilation of a Japonica variety, Gimmasari; by using tracer technique (C'4). Top~dressing of nitrogen at full heading time enhanced carbon assimilation in the leaves by 33 per cent but reduced the movement of assimilated carbon from shoot to grain by 2 to 3 times over that of the control. The nitrogen content of plant increased by 80 per cent due to top-dressing nitrogen at heading with a corresponding increase in the percentage of nitrogen content in leaf by 52 per cent. It was inferred that the high concentration of nitrogen in the shoot might have interfered with the movement of carbon from shoot to grain in the plants top-dressed with nitrogen at heading time.
INTRODUCTION
Recently in japan, there is a tendency amongsome farmers to apply nitrogen after heading stage in rice cultivation. This is popularly called ''the top-dressing for grain". These techni-ques are being tried for higher yield due to the betterment of nutritional status during the ripening stage. Matsushima ( 195 7, 1966) found that top-dressing with nitrogen at full heading time serves to raise the percentage of ripened grains due to promotion of carbon assimilation in the plant. Murayama (1967) also pointed out that N supplied at ripening stage is effective in enhancing carbon nutrition status of rice plant.
The present investigation is aimed at assessing the influ-ence of top~dressing of nitrogen at full heading stage on carbon assimilation by rice during the ripening period.
•
MATERIALS AND METHODS
The seedlings of a Japonica var. Gimmasari were trans-planted at 4-5 leaf stage on May 13, 1968, in pots (4 seedlings per pot) containing 5 · 0 kg of alluvial soil to which 0 · 4 g each of N, P205 and K20 per pot as ammonium sulphate, sodium phosphate and potassium chloride were added at the time of planting. The plants were grown normally upto full heading stage, till July 24, 1968, and were subsequently divided into two series (i) control, no top-dressing, and (ii) top-dressed (0 · 36 g N/pot). Top"dressing was done with N'5 labelled ammonium sulphate (10% N'5 excess) on the surface of soil. A week later, the plan.ts were exposed to C'40
2, liberated by the addition of lactic acid to C'4 tagged barium carbonate of 1 me. activity, in a closed transparent chamber ( 1
x
1 X 1 · 25 m) ~ The treatment was continued for one hr und.er natural sun light. Sixteen pots (eight each in control and treated series) were exposed to C'402 and the treatment was repeated with another lot of 16 pots ·at the 4-week stage after top dressing. Only one plant per pot was exposed to C'4 treatment, while the side plants were covered with aluminium foil during the period of treatment. Samples were drawn from the exposed plants, after 0, 3 and 7 days later and at harvest stage (2 pots for each sample) from the individual treatments and the dry weights of leaf blade, leaf sheath, culm and grain were recorded.Photosynthetic ifficienry (C'4 activiry).-The C'4 activity in
the individual parts i.e. leaf blade, leaf sheath, culm and grain was examined by taking the radioactivity counts with dried and powdered samples. The activity was measured in an automatic sample changer using an end window G.M. tube (1·5 mg/cm2 ) . Twenty mg of powdered material was taken in a stainless steel planchet (in triplicate for each sample) for activity determina-tion. The counts per min (CPM) per g of dry matter were calculated and expressed as specific activity. The dry matter accumulation per plant, the total activity and the proportion of activity in individual parts and the translocation rate (activity in grain/activity in shoot X 100) were calculated. The data are presented in Figs. 1 to 5.
determination, (using N/7 H2S04 for neutralising the ammonia released during distillation with alkali) the solution in the flask was slightly acidified with H2S04 to prevent losses of ammonia
and finally concentrated to about 5 ml on a water bath. The extract containing 2-5 mg of nitrogen was analysed for N'5
excess by the Mass Spectrometer at the Radiation Laboratory, Komagome, Tokyo. The data are presented in Figs. 6 and 7.
Radioautography.-The trend of accumulation of photo-synthates into grain from individual leaves of the control and nitrogen top-dressed plants was also examined at two-week stage after heading. The treatments consisted of (1) top leaf (leaf-1) exposed, (2) leaf-4 exposed and (3) ear exposed to radioactive carbon. A few pots set-apart from the above experiment were utilised for these tests. The plants were covered with aluminium foil except the treated organs and exposed to C'4 for one hr as detailed in the above experiment. The samples were drawn 72 hrs after treatment and the individual leaves, leaf sheath, culm and ear were separated, immediately pressed with electric iron and subsequently exposed to x-ray film for 8 days. The radioautographs were prepared for examination. •
RESULTS AND DISCUSSION
Dry
matter accumulation.-The results are presented in Fig. 1.The accumulation of dry matter into grain was completed within
"f
-H22 / ' 18
"
!;: 10
~
.
'
~ 8!': 7
T
----
G0· .. c ..
'•,
T
.---.G
... - .... ---·-::>LIS
5.
j
~---~~~
r
---c
~---~---_---
:s
' • 1 ' 0 1
WE'El(S AfTER, HEADING
•
4 weeks after flowering even in the N top~dressed plants. The loss in dry matter of the shoot (leaf blade, leaf sheath and culm) at harvest was more in control than in N-treated plants. The dry weight of the culm did not decrease even at harvest under N top-dressing. The dry weight of the plant increased by nearly
30 per cent at harvest time by N top~dressing, while the increase in grain weight was only 10-14 per cent. The results, thus, indicate that the application of N at heading mostly increased the shoot weight with less favourable influence on grain yield.
Photosynthetic activity
of
the plant and the movementof
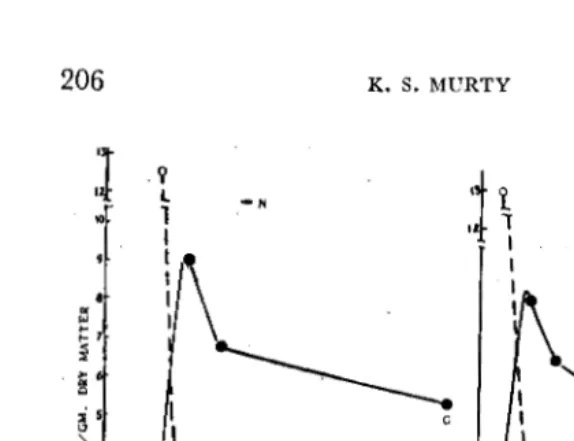
photo-synthates to grain.-The results of photosynthetic efficiency, as assessed from C'4 activity in individual parts of the plant, arepresented in Figs. 2 to 5.
11
10
-N
,·,~
io
I-'
~ 5
u
I
I
I
I
I
I
I
I I
T
f
-\~ ~-·~---·~
.. __
---~--- ---oLb
'X-lC -4LS
o•T
;c
WF.EK~ APT£R HEAD! NG
Fm. 2. Total Ct4 activity (CPM/Plant) in the plant.
The activity of the whole plant increased by 36 percent under N top-dressing (as observed immediately after C'4 feeding), the increase in individual parts being 33% in leaf blade, 52%
9
l
1 I
\\
~
I
I
I.
I . 1 "
I
·~
e--i.',o_ . •rx
,_:-,-~--:..,..,._,_________ ... _ ,.
~
.... '0---
--..3:8
•
----.:::~L>.... '9...---~fs c
I• 6
.r-;-,--z---.._,.
W~EKS. AFTER JlJ!,\.D I :'<G
FIG. 3. Specific activity of QI4 (CPM/gm dry wt.).
at harvest was considerably higher in control plants (86%) than in N-treated ones (63%). Greater proportion of the activity was retained in vegetative organs under N top-dressing (Fig. 4). The translocation rate was found to be much higher in control (2 to 3 times) than in the treated plants (Fig. 5). The radio-autographs also showed lower movement of C'4 from leaf to
grain under N top-dressing (Fig. 8). The samples collected after C'4 feeding at 4 week stage also showed similar results.
100
XI
00
?
0
t.,ro
~"'
~ ;:so
ii!
~., :!!
Q"'
"'
-N
G
~
.
.
.
.
•.+N
q
0
.
.
.
I~·.-G
~
__ .,.:o_-_-_-: ::-_-:-
-~=----
---~8 - --·LSc
-~
•10 1
5 CA• TI.EATMENT 2 WL-:: EKS
AFTEW KEADING
-·-~
,,.
...
o//0
I
---~
C 14 TR EATMEPl'f • WEEXS
AFTD HF.A.Oir«i
~~---t---~.---
,~~7-~~--~--~-9.-EEKS AFTER HEADING
Fro. 5. Translocation of carbon from shoot to grain.
The increased rate of accumulation of carbon by N top" dressing at heading time has been reported earlier (Mitsui and Ishii, 1939; Noguchi, 1949; Matsushima, 1957; Mitsui and Nishigaki, 1963 and Murayama, 1967). However, it is evident from this data that inspite of enhanced carbon fixation by N top-dressing, considerable proportion of assimilated carbon was either retained in vegetative organs or was lost in respiration as suggested by Kumura and Takeda (1962) resulting in poor movement to grain.
Dastur and Chinoy (1932), Suzuki (1953) and Matsushima ( 1966) have indicated that the translocation of assimilated products progressed parallel with carbon assimilation in leaf blade. The results presented in this paper show that inspite of greater C assimilation under N top-dressing, less proportion of the assimilated carbon had moved into grain, indicating there-by that the low rate of translocation was not due to lack of suffi-cient carbon in the leaf (source). Goodal (1946), Bonner and Gals ton ( 1952) and Matsushima ( 1966) have indicated that translocation is also greatly influenced by the sugar gradient from leaf to grain. The sugar concentration was not assessed in the present experiment but the fact remains that the sugar content would be fairly low in the grain as compared to leaf due to rapid conversion of accumulated sugars to starch by phos-phorylase (Aimi, 1967) under the prevailing high temperatures
lower rate of translocation as observed under N top-dressing might not be due to the lack of sugar gradient from leaf to grain. It might be possible that due to application of N at heading stage the nitrogen concentration in the vegetative organs was enhanced and this excess N might have interfered with trans-location of carbon as discussed hereunder.
Nitrogen uptake by plant.-The data on N uptake are presented in Figs. 6 to 8.
The N uptake by the plant was considerably increased (by 80 per cent) under N top-dressing, the increase in individual parts being 52 per cent in leaf blade, 100 per cent in culm and sheath and 28 per cent in grain. The high concentration of N was maintained even up to harvest (Fig. 6). Considerable pro-portion of applied N accumulated into grain and culm (Fig. 7). In general, a .reduction in N'5 content at harvest was evident possibly due to greater absorption of N'4-N from soil or due
to loss of absorbed N from plant by leaching (Tanaka and Navasero, 1964).
I
~f
Ti
---·--~
~---T
~
..
"
~ IDO
!
z
··~.~.~~~---. ~.~.~~~---~
WUl<S AFTf.R HEADING
FIG. 6. Total nitrogen content of the plant.
Matsushima (1966) and Murayama (1967) also pointed out that various factors were associated with the response to applied nitro-gen at heading time. However, the prevalent opinion, that high
l
.
N16 LAflf.LED N l.N PLANT ( .. ) G Nl' EXCESS ~ !D - - ·-:::::--~
#
.~~
~Go,.~...---
0..., ·--._~: o---o-- ___________ : - : : [ 'o,,:::_~
~ LB 0 ·- .... ~S
i. I 1;) -~----~
LB
0 • 7 0 I 7
WEfXS APTER HEADING
FIG. 7. Uptake ofNrLN by the plant.
nitrogen concentration in the plant at heading might disturb the translocation of carbon from leaves to grain, was strongly contradicted by Matsushima and his associates (Matsushima, 1957; Yamada et al., 1957; Matsushima and Wada, 1959). They attributed the adverse effect, if any, to a shortage of carbo-hydrates supplied to grain or to other factors like root damage. However, the present results show that the high concentration of N in the shoot was responsible for poor translocation of carbon to gram. Further work is warranted to substantiate these findings.
No nitrogen Nitrogen top-dressed
FIG. 8. Radioautograph-showing movement of assimilated carbon from leaf to the ear.
AcKNOWLEDGEMENT
National Institute of Agricultural Sciences, Tokyo as a parti-cipant under Colombo Plan. The help rendered by the staff of Plant Nutrition Division of National Institute of Agricultural Sciences, Tokyo is greatly acknowledged. Grateful thanks of the author are due to Drs. N. Murayama, T. Kiuchi, S. Koba-yashi, A. Suzuki and T. Yoshida for their constant guidance during the course of present investigation.
REFERENCES
Aimi, R. (1967). Cell physiological and biochemical aspects in the physiology of ripening. IRC News Letter, Special Issue, p. 106-11.
Bonner, J. and Galston, A. W. (1952). Translocation: The redistribution of nutrients.
Principles of Plant Physiology W. H. Freeman and Co. San Francisco.
p. 148-70.
Dastur, R. H. and Chinoy, J. J. (1932). Carbon dioxide assimilation of the leaves of the rice plant, Oryza Sativa L. Ind. J. Agric. Sci., 11: 431-54.
Goodal, D. W. (1946). The distribution of weight change in the young tomato plant. Ann. Bot., 10: 305-38.
Kumura, A. (1956). Studies 0':1 the effect of internal nitrogen concentration of rice plants on the constitutional factor of yield. Proc. Crop. Sci. Soc. Japan,
24:324-30.
- - - - , a n d Takeda, T. (1962). Analysis of grain production in rice plant. VII. Influence of nitrogen on grain production of rice plant. Proc. Crop. Sci. Soc., Japan, 30: 261--65.
Matsushima, S. (1957). Analysis of developmental factors determining yield and yield predictions in low land rice. Bull. Natl. Inst. Agric. Sci., Japan, Ser.
A, 5: 1-271.
- - - - . (1966). Top dressing with nitrogenous fertilisers at full heading time.
Crop Science in rice. Fuji Pub!. Co. Ltd., Japan, pp. 241-47.
- - - - and Wada, G. (1959). Relation of the amount of carbohydrates and nitrogen contents in plant to the percentage of ripened grains and yield of rice with special reference to nitrogen top dressing at full heading time.
Agric. and Hortic. 34(1), (2), 1-4: 303-06.
Mitsui and Ishii, Y. (1939). Effect of nitrogen top-dressing upon carbon assimi-lation of rice plants. J. Sci. Soil and Manu. Japan., 13(5): 309-13.
- - - - and Nishigaki (1963). Quoted from the theory and practice of growing rice
Overseas Technical Co-operation Agency, Tokyo, Japan, 1963, p. 95.
Murata, Y. (1967). Analysis of growth in relation to ripening of rice. IRC News Letter, Spl. issue, 1967, p. 43.
Murayama, N. (1967). Nitrogen nutrition of rice plant. J. A. R. Q., 2(2): 1-5.
Noguchi, Y. (1949). Studies on nutritional physiology in rice plants. Agric. and Hortic, 24: 757--60.
Suzuki, N. (1953). Cited from crop science in rice by Matsushima, S. Fuji Pub!.
Co. Ltd., Japan, 1966. p. 188.
Tanaka, A. and Navasero, S. A. (1964). Loss of nitrogen from the rice plant through rain or dew. Soil Science and Pl. Nutrition, 10: 36-39.
- - - - , , Garcia, C. V., Parao, F. T. and Ramirez, E. (1964). Growth habit of rice plant in the tropics and its effect on nitrogen response. Tech. Bull. No. 3, IRRI, Philippines.
Yamada, N., Ota, Y. and Kushibushi, K. (1957). Studies on ripening of rice (I) Role of nitrogen in process of ripening of rice. Proc. Crop. Sci. Soc. Japan,