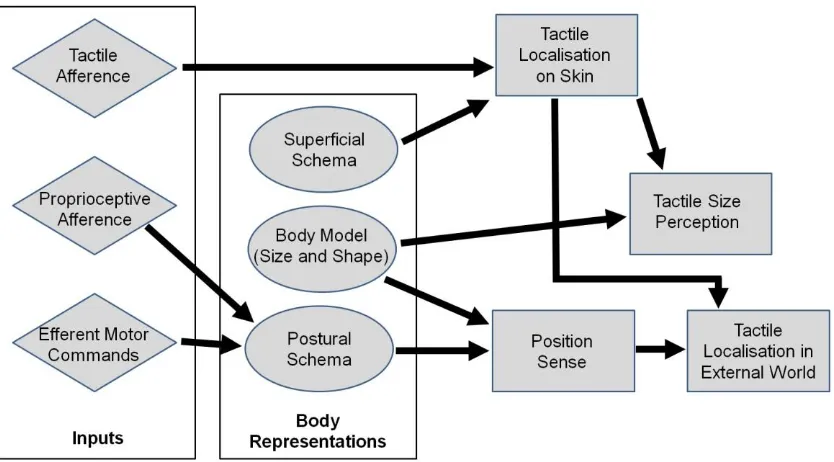
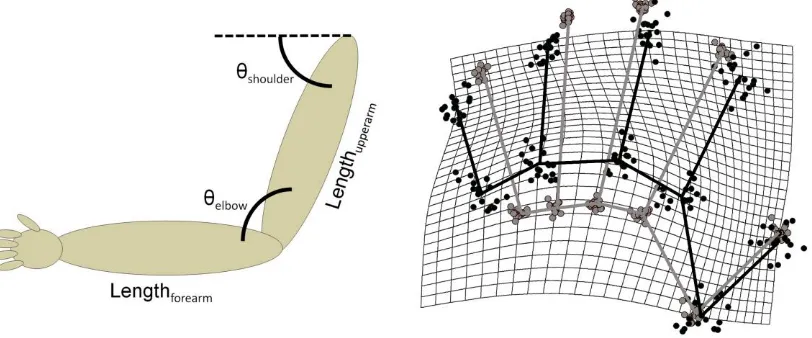
Implicit and explicit body representations
Full text
Figure
Related documents
BCG: Bacille de Calmette et Guérin; CER: cost effectiveness ratio; CFR: case fatality ratio; DALY: disability adjusted life year; DTP-Hep-Hib: diphteria, tetanus, pertussis,
1997 – 2002 Board Member- Rochester Community Eye Care Management Organization (RCEMO) 1998 – 2002 Board Member -Managed Care Committee – New York State Optometric Association
To estimate a possible impact of the first assistant on the perioperative blood loss, three parameters were analysed in this study: hemoglobin level before the surgery and 24h after
quer cetin, t etrah ydr ox ymetho xyfla vone , apigenin meth yl ether , dimeth ylquer - cetigetin, lut eolin Alum W ool Quer cetin, lut eolin meth yl ether , t etrah y- dr ox
regions may be negatively associated with the level of per capita income in a given region, because an increase in the level of human capital in the neighbouring
If you are in a Health Maintenance Organization (HMO), Exclusive Provider Organization (EPO), or Point of Service (POS) Plan and you move or are transferred out of that Plan’s
Figure 3 shows the depth of the primarily Cenozoic sedimen- tary cover to the acoustic basement (LS1 and ESS1, respec- tively) on the Laptev and East Siberian shelves in millisec-
Yeast cells pretreated with conditioned medium which had been diluted with a volume of fresh medium and then chal- lenged with a lethal dose of hydrogen peroxide exhibited
