Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Differential Identification of
Mycobacterium tuberculosis
Complex and
Nontuberculous Mycobacteria by Duplex PCR Assay Using the RNA
Polymerase Gene (rpoB)
Bum-Joon Kim,
1Seong-Karp Hong,† Keun-Hwa Lee,
1Yeo-Jun Yun,
1Eui-Chong Kim,
2Young-Gil Park,
3Gil-Han Bai,
3and Yoon-Hoh Kook
1*
Department of Microbiology and Cancer Research Institute, Institute of Endemic Diseases, SNUMRC, Seoul National University College of Medicine, and Clinical Research Institute,1and Department of Clinical Pathology, Seoul National University
College of Medicine, Seoul 110-799,2and The Korean Institute of Tuberculosis, Korean
National Tuberculosis Association, Seoul 137-140,3Korea
Received 8 July 2003/Returned for modification 20 August 2003/Accepted 18 November 2003
A novel duplex PCR method that can amplify the 235- and 136-bprpoBDNAs ofMycobacterium tuberculosis
complex and nontuberculous mycobacteria (NTM), respectively, with two different sets of primers was used to differentially identify 44 reference strains and 379 clinical isolates of mycobacteria in a single-step assay. Showing 100% sensitivity and specificity, the duplex PCR method could clearly differentiateM. tuberculosis
complex and NTM strains. In addition, restriction fragment length polymorphism analysis and direct sequenc-ing of the amplicon of NTM could be used to supplement species identification.
With the recent global resurgence of mycobacterial infec-tions, especially of tuberculosis, attributed to increased human immunodeficiency virus infection, there is an increasing de-mand for rapid, sensitive, and specific diagnostic methods for the detection and identification ofMycobacterium tuberculosis
and nontuberculous mycobacteria (NTM) in a clinical setting (2, 3, 4). NTM infection can cause clinical problems, as its pathogenic potential and susceptibilities to antituberculosis treatments vary (22). In addition, mixed infections ofM. tuber-culosis and NTM have been reported (16). Therefore, it has become important to be able to differentiate between the two during the early stage of the diagnostic procedure.
The diagnosis of mycobacterial infection is accomplished by culture-based identification. Primary culture of slowly growing mycobacteria, without using the BACTEC culture system, usu-ally takes 4 to 6 weeks or longer (10). However, recent meth-odological advances in molecular biology have provided alter-native rapid approaches, e.g., the PCR and PCR-linked methods. For the rapid detection or identification ofM. tuber-culosis, target genes specific to mycobacteria are used in a PCR (7, 8, 17, 19).
Because the incidence of NTM infection is increasing, any methods capable of simultaneously determining the presence ofM.tuberculosisand/or NTM would be useful. For this pur-pose, multiplex PCR, which simultaneously uses two or three different genes, has been frequently used, as the technique can specifically detect and identify different species of the genus
Mycobacterium(6, 15, 18) and differentiate members of theM.
tuberculosiscomplex (6, 9) in the routine diagnostic laboratory by using Mycobacterium genus- and species-specific genes.
However, some of these genes have been found to lack spec-ificity forM. tuberculosis. In addition, IS6110PCR has been reported to produce false-negative (23) and false-positive (11) results, and themtp40gene is not present in allM.tuberculosis
strains (21). These reports suggest that the multiplex PCR targeting of these genes has associated problems.
In the present study, we used a simplified multiplex PCR assay, basically a duplex PCR (DPCR) assay, to differentiate
M.tuberculosiscomplex and NTM by using a single gene, the RNA polymerase-subunit-encoding gene (rpoB). To demon-strate the efficiency and usefulness of the DPCR assay in this context, we used it to identify reference strains and clinical isolates of mycobacteria.
Forty-four mycobacteria and 17 non-mycobacteria were used as reference strains (Table 1). Three hundred seventy-nine clinical isolates (193 M. tuberculosis complex [183 M.
tuberculosisand 10M.bovis] and 186 NTM isolates), which had been isolated by the Korean Institute of Tuberculosis and the Seoul National University Hospital, were identified by conven-tional biochemical tests and provided for blind testing. An IS6110 PCR assay and analysis of partial 16S rRNA gene (rDNA) sequences were performed separately, and the results were compared with those of the DPCR assay.
Two different kinds of DNA extraction protocols were used for type strains and clinical isolates. The DNAs of the refer-ence strains were purified by using the bead beater-phenol extraction method as previously described (12, 13). The DNAs of clinical isolates were prepared by the boiling method with-out a purification step to reduce the DNA preparation time and to minimize the risk of cross contamination (1). A loopful culture of each sample was suspended in a screw-cap tube with 50l of TEN buffer (10 mM Tris-HCl, 1 mM EDTA, 100 mM NaCl [pH 8.0]), and the tube contents were then boiled on a hot plate for 10 min. Without further purification, 5l of the boiled mycobacterial suspension was used directly as a tem-plate for the DPCR assay.
* Corresponding author. Mailing address: Department of Microbi-ology, Seoul National University College of Medicine, 28 Yongon-Dong, Chongno-Gu, Seoul 110-799, Korea. Phone: 82-2-740-8306. Fax: 82-2-743-0881. E-mail: [email protected].
† Present address: Department of Microbiology, College of Medi-cine, The Catholic University of Korea, Seoul 137-701, Korea.
1308
on May 15, 2020 by guest
http://jcm.asm.org/
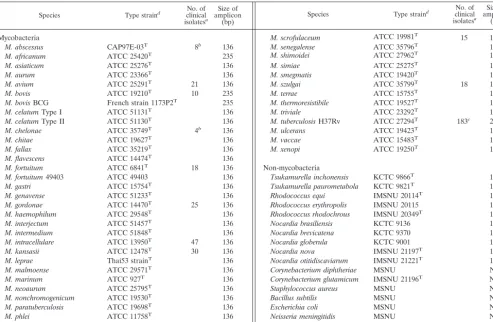
For construction of the two DPCR primer sets, rpoB se-quences of 44 mycobacteria (GenBank accession no. AF057449 to AF057493) were aligned by using the multiple alignment algorithm in the MegAlign package (Windows, ver-sion 3.12e; DNASTAR, Madison, Wis.). The specific nucleo-tides of theM.tuberculosisor NTM strains were located at the 3⬘-hydroxyl end of each primer, as previously reported (12). The developed Tbc1 (5⬘-CGT ACG GTC GGC GAG CTG ATC CAA-3⬘)-TbcR5 (5⬘-C CAC CAG TCG GCG CTT GTG GGT CAA-3⬘) and M5 (5⬘-G GAG CGG ATG ACC ACC CAG GAC GTC-3⬘)-RM3 (5⬘-CAG CGG GTT GTT CTG GTC CAT GAA C-3⬘) primer sets amplify a 235-bp DNA sequence from theM.tuberculosiscomplex and a 136-bp DNA sequence from NTM, respectively (Fig. 1A). Primers (10 pmol of Tbc1-TbR5 and 20 pmol of M5-RM3) and 5l of bacterial DNA were added to a PCR mixture tube (AccuPower PCR PreMix; Bioneer, Daejeon, Korea) containing 2 U ofTaq poly-merase, 250 M each deoxynucleoside triphosphate, 10 mM Tris-HCl (pH 8.3), and 1.5 mM MgCl2, and water was added to a final volume of 20l per reaction mixture. PCR was per-formed with an initial denaturation of 95°C for 5 min, 30 cycles of amplification (30 s at 95°C, 60 s at 72°C), and a final elon-gation at 72°C for 5 min (model 9600 thermocycler;
Perkin-Elmer Cetus). Denaturation was extended to 15 min for the clinical isolates.Escherichia coliDNA was used as a negative control. The PCR products were analyzed by agarose gel (1.5%) electrophoresis.
After completion of the DPCR assay, all 186 NTM isolates showing a 136-bp DNA amplicon were further analyzed by restriction fragment length polymorphism (RFLP) and by di-rect sequencing for species identification. On the basis of the
rpoB sequences of mycobacteria (GenBank accession no. AF057449 to AF057493), two restriction enzymes,MspI and
HaeIII were selected by using MapDraw (version 3.14; DNASTAR). Two enzymes,MspI (TaKaRa, Shiga, Japan) and
HaeIII (TaKaRa) were independently applied to the PCR products. Ten microliters of the PCR products, 2 U of each enzyme, and restriction buffer were transferred to a fresh mi-crocentrifuge tube, and water was added to a final volume of 20 l per reaction mixture. Digestion was performed for 2 h at 37°C. Following digestion, the mixtures were electrophoresed in a 3% agarose gel.
[image:2.603.47.541.82.404.2]NTM isolates were separately identified by determining 87-bp amplicon sequences, which excluded the primer se-quences, and by comparing these with sequences in the Gen-Bank database. Sequencing reactions with primers M5 and
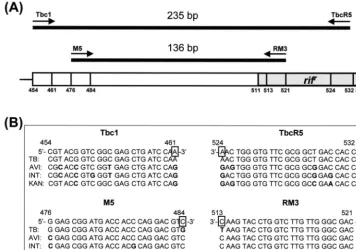
TABLE 1. Mycobacteria and non-mycobacteria used for DPCR and their PCR amplification products
Species Type straind clinicalNo. of
isolatesa
Size of amplicon
(bp) Species Type strain
d clinicalNo. of
isolatesa
Size of amplicon
(bp)
Mycobacteria
M. abscessus CAP97E-03T 8b 136
M. africanum ATCC 25420T 235
M. asiaticum ATCC 25276T 136
M. aurum ATCC 23366T 136
M. avium ATCC 25291T 21 136
M. bovis ATCC 19210T 10 235
M. bovisBCG French strain 1173P2T 235
M. celatumType I ATCC 51131T 136
M. celatumType II ATCC 51130T 136
M. chelonae ATCC 35749T 4b 136
M. chitae ATCC 19627T 136
M. fallax ATCC 35219T 136
M. flavescens ATCC 14474T 136
M. fortuitum ATCC 6841T 18 136
M. fortuitum49403 ATCC 49403 136
M. gastri ATCC 15754T 136
M. genavense ATCC 51233T 136
M. gordonae ATCC 14470T 25 136
M. haemophilum ATCC 29548T 136
M. interjectum ATCC 51457T 136
M. intermedium ATCC 51848T 136
M. intracellulare ATCC 13950T 47 136
M. kansasii ATCC 12478T 30 136
M. leprae Thai53 strainT 136
M. malmoense ATCC 29571T 136
M. marinum ATCC 927T 136
M. neoaurum ATCC 25795T 136
M. nonchromogenicum ATCC 19530T 136
M. paratuberculosis ATCC 19698T 136
M. phlei ATCC 11758T 136
M. peregrinum ATCC 14467T 136
aClinical isolates were identified by biochemical tests and analysis of partial 16S rDNA sequences. bDifferentiation ofM. abscessusandM. chelonaewas performed byrpoB(306-bp) sequence analysis (13). cComposed of 40 rifampin-resistant and 143 rifampin-susceptible strains.
dATCC: American Type Culture Collection; KCTC: Korean Collection for Type Cultures; MSNU: Department of Microbiology, Seoul National University College
of Medicine; IMSNU: Institute of Microbiology, Seoul National University. Superscript T indicates type reference strain. There were a total of 379 clinical isolates.
eNA, not amplified.
M. scrofulaceum ATCC 19981T 15 136
M. senegalense ATCC 35796T 136
M. shimoidei ATCC 27962T 136
M. simiae ATCC 25275T 136
M. smegmatis ATCC 19420T 136
M. szulgai ATCC 35799T 18 136
M. terrae ATCC 15755T 136
M. thermoresistibile ATCC 19527T 136
M. triviale ATCC 23292T 136
M. tuberculosisH37Rv ATCC 27294T 183c 235
M. ulcerans ATCC 19423T 136
M. vaccae ATCC 15483T 136
M. xenopi ATCC 19250T 136
Non-mycobacteria
Tsukamurella inchonensis KCTC 9866T 136
Tsukamurella paurometabola KCTC 9821T 136
Rhodococcus equi IMSNU 20114T 136
Rhodococcus erythropolis IMSNU 20115 136 Rhodococcus rhodochrous IMSNU 20349T 136
Nocardia brasiliensis KCTC 9136 136
Nocardia brevicatena KCTC 9370 136
Nocardia globerula KCTC 9001 136
Nocardia nova IMSNU 21197T 136
Nocardia otitidiscaviarum IMSNU 21221T 136
Corynebacterium diphtheriae MSNU NAe
Corynebacterium glutamicum IMSNU 21196T NA
Staphylococcus aureus MSNU NA
Bacillus subtilis MSNU NA
Escherichia coli MSNU NA
Neisseria meningitidis MSNU NA
Haemophilus influenzae MSNU NA
on May 15, 2020 by guest
http://jcm.asm.org/
RM3 were performed as previously described (13). Analysis of the partial 16S rDNA sequence was performed separately to identify the NTM strains, and the results were compared with the results of therpoBsequence analysis. Briefly, 16S rDNA fragments were amplified by using forward primer 285 and reverse primer 264 and then directly sequenced by using se-quencing primer 244, as previously described (14).
The specificity of each primer set was assessed by a separate PCR. When a PCR using each specific primer set (Tbc1-TbcR5 or M5-RM3) was applied to the 44 reference strains at the same annealing temperature, an amplicon of either 235 or 136 bp of DNA was observed from theM.tuberculosiscomplex or
NTM, respectively. When this method was applied to 17 non-mycobacteria, only 10 strains of Tuskamurella, Rhodococcus, andNocardiaspecies, which are phylogenetically close to my-cobacteria, produced a 136-bp DNA amplicon (Table 1). Fi-nally, a DPCR assay with a mixture of the two primer sets was performed on the reference strains. While the 235-bp DNAs were amplified from only four strains ofM.tuberculosis com-plex, the 136-bp DNAs were amplified from all of the 40 NTM strains (Fig. 2). Nothing was amplified from the negative con-trol,E.coliDNA. Therefore, DPCR assay allowed the differ-ential identification ofM.tuberculosiscomplex and NTM in a single reaction. Despite the mismatching of several nucleotides with those of the M5 or RM3 primer in therpoBsequences of NTM (Fig. 1B), all of the NTM strains tested produced 136-bp amplicons.
[image:3.603.113.469.72.322.2]In order to determine the lower limit of the sensitivity of the DPCR method,rpoBDNAs were amplified from the serially diluted DNAs (10 ng to 10 fg) ofM.tuberculosisandM.avium.
FIG. 1. Locations (A) and sequences (B) of the DPCR primers on therpoBgene. DPCR products amplified by anM.tuberculosis-specific primer set (Tbc1-TbcR5) and an NTM-specific primer set (M5-RM3) were 235 and 136 bp long, respectively.rifrindicates the region associated with the rifampin resistance ofM.tuberculosis. Nucleotides of four mycobacteria differing from those of the primers are shown in bold letters. Numbers indicate the amino acid position of the RNA polymerasesubunit ofE.coli. Note that TbcR5 and RM3 are displayed in reverse direction. TB, M. tuberculosis H37Rv (GenBank accession no. AF057454); AVI, M. avium (GenBank accession no. AF057457); INT, M.
intracellulare(GenBank accession no. AF057472); KAN,M.kansasii(GenBank accession no. AF057473).
FIG. 2. DPCR assay results performed with the reference strains of
M.tuberculosiscomplex and NTM. Two amplicons of different sizes (235 and 136 bp) were amplified fromM.tuberculosiscomplex (lanes 1 to 4) and NTM (lanes 5 to 14) strains by a single DPCR. Lanes: M, marker DNA (50-bp ladder); 1,M.tuberculosis; 2,M.africanum; 3,M.
bovis; 4,M.bovis BCG; 5, M. avium; 6,M. paratuberculosis; 7, M.
[image:3.603.43.284.561.641.2]scrofulaceum; 8,M.intracellulare; 9,M.terrae; 10,M. nonchromogeni-cum; 11,M.triviale; 12,M.gordonae; 13,M.asiaticum; 14,M. thermore-sistibile; 15,E.coli(negative control).
TABLE 2. Differentiation ofM. tuberculosiscomplex and NTM clinical isolates by DPCR and conventional biochemical testing
DPCR product (size [bp])
No. of isolates identifiedaas:
M. tuberculosis
(n⫽193) (nNTM⫽186)
M. tuberculosisspecific (235) 193 0
NTM specific (136) 0 186
aIdentification was separately performed by niacin and heat-stable catalase
tests (pH 7.0, 68°C).
on May 15, 2020 by guest
http://jcm.asm.org/
The amplification product was obtained from 10 pg of M.
tuberculosisDNA, whereas in the case ofM. aviumDNA, as little as 1 pg was detected on an ethidium bromide-stained gel (data not shown). Taking 5 fg of DNA as a mycobacterial cell equivalent (5), this method could detect 2,000 bacillus equiv-alents.
The usefulness of the DPCR assay was demonstrated by applying the technique to the identification of 379 clinical isolates. Of the 379 culture isolates examined, 193 strains were identified asM.tuberculosiscomplex and the other 186 strains were identified as NTM (Table 1). These results were com-pletely concordant with those obtained by conventional culture testing (Table 2). Although primer TbcR5 is located in therifr
region (20) (Fig. 1A), in which mutations are related to the rifampin resistance of M.tuberculosis, all of the 40 rifampin-resistant M. tuberculosis isolates produced 235-bp amplicons without PCR interference.
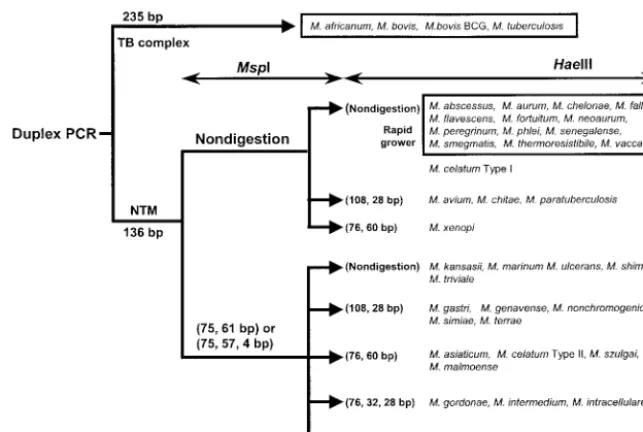
Since DPCR produces only one (136-bp)rpoBDNA from NTM, restriction analysis and sequencing could be used for further species identification. An algorithm for species identi-fication by DPCR-linked restriction analysis with MspI and
HaeIII was developed (Fig. 3). All of the 186 NTM isolates analyzed in this study were tentatively identified in accordance with this scheme. However, NTM isolates, exceptM.abscessus
andM.chelonae, which have identical sequences in the 87-bp
rpoB region, could be exactly identified by comparing their determined rpoB sequences to those of reference strains. These results are concordant with those obtained by 16S rDNA analysis and culture-based biochemical testing.
In the present study, we developed a novel DPCR assay based onrpoB sequences. Unlike previous methods, two dif-ferently sized DNAs were amplified from a single target gene of M. tuberculosisand NTM. The advantages of this DPCR assay over other multiplex PCRs are as follows. (i) rpoB nu-cleotides specific forM. tuberculosiscomplex or NTM are in-variably constant. Therefore, false-positive or -negative results
due to sequence variations do not occur. (ii) DPCR yields only one product, irrespective of theMycobacteriumspecies tested. Even the coexistence ofM.tuberculosisand NTM can be de-tected by the presence of two different PCR products in a single reaction mixture. (iii) Moreover, the (136-bp) rpoB
DNAs of NTM can be further analyzed by RFLP or by direct sequencing to supplement species identification.
In conclusion, the DPCR assay based onrpoB provides a rapid and reliable means for the differential identification of
M.tuberculosisand NTM in culture with a single reaction.
B.-J. Kim and S.-K. Hong contributed equally to this work. This study was supported by a grant from the Korean Health 21 R&D Project, Ministry of Health & Welfare, Republic of Korea (01-PJ10-PG6-01GM03-0002), and in part by the BK21 project for Medi-cine, Dentistry, and Pharmacy.
REFERENCES
1. Afghani, B., and H. R. Stutman.1996. Polymerase chain reaction for
diag-nosis ofM.tuberculosis: comparison of simple boiling and a conventional method for DNA extraction. Biochem. Mol. Med.57:14–18.
2. Barnes, P. F., A. B. Bloch, P. T. Davison, and D. E. Sneider, Jr.1991.
Tuberculosis in patients with human immunodeficiency virus infection. N. Engl. J. Med.324:1644–1649.
3. Bloom, B. R.1992. Back to a frightening future. Nature (London)358:538– 539.
4. Bloom, B. R., and C. J. L. Murray.1992. Tuberculosis: commentary on a
reemergent killer. Science257:1055–1064.
5. Cole, S. T., R. Brosch, J. Parkhill, T. Garnier, C. Churcher, D. Harris, S. V. Gordon, K. Eiglmeier, S. Gas, C. E. Barry III, F. Tekaia, K. Badcock, D.
Basham, D. Brown, et al.1998. Deciphering the biology ofMycobacterium
tuberculosisfrom the complete genome sequence. Nature393:537–544.
6. Del Portillo, P., M. C. Thomas, E. Martínez, C. Maran˜on, B. Valladares,
M. E. Patarroyo, and M. C. Lo´pez.1996. Multiprimer PCR system for
differential identification of mycobacteria in clinical samples. J. Clin. Micro-biol.34:324–328.
7. Eing, B. R., A. Becker, A. Sohns, and R. Ringelmann.1998. Comparison of
RocheCobas AmplicorMycobacterium tuberculosisassay with in-house PCR and culture for detection ofM.tuberculosis. J. Clin. Microbiol.36:2023–2029.
8. Gunisha, P., H. N. Madhavan, U. Jayanthi, and K. L. Therese.2001.
Poly-merase chain reaction using IS6110 primer to detectMycobacterium tuber-culosisin clinical samples. Indian J. Pathol. Microbiol.44:97–102.
[image:4.603.131.453.67.283.2]9. Herrera, E. A., O. Perez, and M. Segovia.1996. Differentiation between
FIG. 3. Algorithm for species identification by DPCR-RFLP (136-bp amplicons) withMspI andHaeIII.
on May 15, 2020 by guest
http://jcm.asm.org/
Mycobacterium tuberculosisandMycobacterium bovisby a multiplex-poly-merase chain reaction. J. Appl. Bacteriol.80:596–604.
10. Jeager, H., Jr., J. Lacy, L. R. Smith, and C. A. LeMaistre.1967. Quantitative studies of mycobacterial population in sputum and saliva. Am. Rev. Respir. Dis.95:998–1004.
11. Kent, L., T. D. McHugh, O. Billington, J. W. Dale, and S. H. Gillespie.1995. Demonstration of homology between IS6110ofMycobacterium tuberculosis and DNAs of otherMycobacteriumspp.? J. Clin. Microbiol.33:2290–2293. 12. Kim, B. J., K. H. Lee, B. N. Park, S. J. Kim, E. M. Park, Y. G. Park, G. H.
Bai, S. J. Kim, and Y. H. Kook.2001. Detection of rifampin-resistant
My-cobacterium tuberculosisin sputa by nested PCR-linked single-strand confor-mation polymorphism and DNA sequencing. J. Clin. Microbiol.39:2610– 2617.
13. Kim, B. J., S. H. Lee, M. A. Lyu, S. J. Kim, G. H. Bai, G. T. Chae, E. C. Kim, C. Y. Cha, and Y. H. Kook.1999. Identification of mycobacterial species by comparative sequence analysis of the RNA polymerase gene (rpoB). J. Clin. Microbiol.37:1714–1720.
14. Kirschner, P., B. Springer, U. Vogel, A. Meier, A. Wrede, M. Kiekenbeck,
F. C. Bange, and E. C. Bottger.1993. Genotypic identification of mycobac-teria by nucleic acid sequence determination: report of a 2-year experience in a clinical laboratory. J. Clin. Microbiol.31:2882–2889.
15. Klemen, H., A. Bogiatzis, M. Ghalibafian, and H. H. Popper.1998. Multiplex polymerase chain reaction for rapid detection of atypical mycobacteria and Mycobacterium tuberculosiscomplex. Diagn. Mol. Pathol.7:310–316. 16. Libanore, M., R. Bicocchi, and F. Ghinelli.1992. Mixed bronchial infection
due toMycobacterium tuberculosisandMycobacterium avium-intracellularein an AIDS patient. Infection20:298–299.
17. Montenegro, S. H., R. H. Gilman, P. Sheen, R. Cama, L. Caviedes, T.
Hopper, R. Chambers, and R. A. Oberhelman.2003. Improved detection of
Mycobacterium tuberculosisin Peruvian children by use of a heminested IS6110polymerase chain reaction assay. Clin. Infect. Dis.36:16–23.
18. Mustafa, A. S., A. T. Abal, and T. D. Chugh.1999. Detection of
Mycobac-terium tuberculosiscomplex and non-tuberculous mycobacteria by multiplex polymerase chain reactions. East Mediterr. Health J.5:61–70.
19. Suffys, P., J. C. Palomino, S. Cardoso Leao, C. Espitia, A. Cataldi, A. Alito, M. Velasco, J. Robledo, J. Fernandez, P. da Silva Rosa, and M. I. Romano.
2000. Evaluation of the polymerase chain reaction for the detection of Mycobacterium tuberculosis. Int. J. Tuberc. Lung. Dis.4:179–183. 20. Telenti, A., P. Imboden, F. Marchesi, D. Lowrie, S. Cole, M. J. Colston, L.
Matter, K. Schopfer, and T. Bodmer.1993. Detection of rifampin-resistance mutations inMycobacterium tuberculosis. Lancet341:647–650.
21. Weil, A., B. B. Plikaytis, W. R. Butler, C. L. Woodley, and T. M. Shinnick.
1996. Themtp40gene is not present in all strains ofMycobacterium tuber-culosis. J. Clin. Microbiol.34:2309–2311.
22. Wolinsky, E.1992. Mycobacterial diseases other than tuberculosis. Clin.
Infect. Dis.15:1–10.
23. Yuen, L. K., B. C. Ross, K. M. Jackson, and B. Dwyer.1993. Characterization ofMycobacterium tuberculosisstrains from Vietnamese patients by Southern blot hybridization. J. Clin. Microbiol.31:1615–1618.