RESEARCH ARTICLE
MicroRNAs regulate survival in oxygen-deprived environments
Simon G. English, Hanane Hadj-Moussa and Kenneth B. Storey*ABSTRACT
Some animals must endure prolonged periods of oxygen deprivation to survive. One such extreme model is the northern crayfish (Orconectes virilis), that regularly survives year-round hypoxic and anoxic stresses in its warm stagnant summer waters and in its cold, ice-locked winter waters. To elucidate the molecular underpinnings of anoxia resistance in this natural model, we surveyed the expression profiles of 76 highly conserved microRNAs in crayfish hepatopancreas and tail muscle from normoxic, acute 2 h anoxia, and chronic 20 h anoxia exposures. MicroRNAs are known to regulate a diverse array of cellular functions required for environmental stress adaptations, and here we explored their role in anoxia tolerance. The tissue-specific anoxia responses observed herein, with 22 anoxia-responsive microRNAs in the hepatopancreas and only four in muscle, suggest that microRNAs facilitate a reprioritization of resources to preserve crucial organ functions. Bioinformatic microRNA target enrichment analysis predicted that the anoxia-downregulated microRNAs in hepatopancreas targeted Hippo signalling, suggesting that cell proliferation and apoptotic signalling are highly regulated in this liver-like organ during anoxia. Compellingly,miR-125-5p, miR-33-5pand miR-190-5p, all known to target the master regulator of oxygen deprivation responses HIF1 (hypoxia inducible factor-1), were anoxia downregulated in the hepatopancreas. The anoxia-increased transcript levels of the oxygen-dependent subunit HIF1αhighlight a potential critical role for miRNA-HIF targeting in facilitating a successful anoxia response. Studying the cytoprotective mechanisms in place to protect against the challenges associated with surviving in oxygen-poor environments is critical to elucidating the vast and substantial role of microRNAs in the regulation of metabolism and stress in aquatic invertebrates.
KEY WORDS:Orconectes virilis, Anoxia tolerance, Metabolic rate depression, HIF1
INTRODUCTION
In the winter, the freshwaters of eastern North America become ice-locked, cutting off access to fresh nutrients, and thus rendering waters anoxic (Bradford, 1983), while in the summer, warm and stagnant waters present a hypoxia-stressed environment. As such, aquatic organisms living in these conditions are faced with the year-round challenge of oxygen-poor waters. Among the organisms that have adapted to these conditions is the northern crayfish (Orconectes virilis). Prevalent throughout much of North America, as far north as the coast of Hudson Bay and extending south into the northern USA,
this organism displays profound hypoxia and anoxia tolerance. Indeed, it was observed that while most fish species migrate downstream to escape ice-locked waters, O. virilis tolerate these oxygen-poor environments, conditions that were lethal to hibernating leopard frogs at the same location (Schueler and Karstad, 1996) Although the crayfishes’physiological response to anoxia has not been thoroughly studied, comparable species exhibit a drop in overall metabolic rate by approximately 90% when exposed to a similar oxygen-deprived environment (Storey and Storey, 1990). Anoxia tolerance is a complex balance of suppressing non-essential functions while simultaneously upregulating basic survival and protective mechanisms to conserve limited energy supplies until favourable conditions are restored (Seibel et al., 2018; Hochachka and Lutz, 2001).
Anoxia and hypoxia tolerance are two stress-induced survival responses that are characterized by a reduction in net ATP turnover, stabilization of macromolecules for extended preservation and the reprioritization of cellular resources (Storey, 2015). In oxygen-poor environments, cell functions such as vascularization and proliferation are controlled by transcription factors such as hypoxia-inducible factor (HIF)-1 (Ziello et al., 2007). A variety of invertebrates have adapted to extreme hypoxic and anoxic conditions and employ these mechanisms to promote survival (Dawson and Storey, 2011). Under such conditions, these processes have been found to be differentially regulated in a tissue-specific manner in response to the unique metabolic requirements of each organ (Ziello et al., 2007). For instance, anoxia-induced responses have been shown to be tissue specific in various other models of hypoxia and anoxia tolerance, such as turtles, frogs and squid (Mattice et al., 2018; Al-attar et al., 2017; Gerber et al., 2016; Hadj-Moussa et al., 2018; Krivoruchko and Storey, 2015).
The molecular mechanisms that underlie anoxia tolerance adaptations include: post-translational modifications, epigenetic regulation, transcription factor regulation, the accumulation of organic osmolytes, and the post-transcriptional regulation of small regulatory non-coding RNAs called microRNAs (miRNAs) (Szereszewski and Storey, 2018; Wijenayake et al., 2018; Biggar and Storey, 2017; Green and Storey, 2016; Kornfeld et al., 2012; Krivoruchko and Storey, 2010; Muir et al., 2007). miRNAs are highly conserved sequences of non-coding RNA approximately 22 nucleotides long that have been identified as critical regulators of a diverse range of cellular functions through their ability to suppress translation (Lund et al., 2004; Christopher et al., 2016). Indeed, the need for energetically inexpensive regulatory mechanisms to coordinate stress responses has drawn attention to the role of miRNAs: (i) as they can rapidly and reversibly bind and unbind their mRNA targets to exert short-term gene repression, and (ii) as the molecules are short, they are energetically inexpensive and disposable modulators. Moreover, there is large potential for regulation by these short nucleotide sequences, as one miRNA may bind to a variety of mRNA transcripts, and a single transcript may bind many miRNAs (Mazière and Enright, 2007).
To further understand the ubiquitous role of miRNAs within the cell, the stress response of the northern crayfish was examined by Received 13 August 2018; Accepted 7 October 2018
Institute of Biochemistry and Department of Biology, Carleton University, 1125 Colonel By Drive, Ottawa, ON, Canada K1S 5B6.
*Author for correspondence (kenneth.storey@carleton.ca)
S.G.E., 0000-0001-5896-9453; H.H., 3338-9046; K.B.S., 0000-0002-7363-1853
Journal
of
Experimental
quantifying an array of 76 miRNAs using quantitative real-time PCR (qPCR) under normoxic, 2 h acute anoxic and 20 h chronic anoxic conditions. Two tissues that exhibit different overall metabolic responses, the hepatopancreas and tail muscle, were selected for this study (Cowan et al., 2001). These differences were highlighted by the changes in anoxia-responsive miRNAs identified, where hepatopancreas anoxia responses were found to be heavily mediated by miRNAs, while muscle tissues displayed a lesser dependence. Taken together, the differentially expressed miRNAs were found to be involved in facilitating an adaptive response to anoxia tolerance.
MATERIALS AND METHODS Animals
Freshwater Orconectes virilis(Hagen 1870) were caught in the Rideau River in Ottawa, ON, Canada, near Carleton University. Crayfish were acclimated in aerated buckets for 7 days at 15°C and then randomly separated into three containers, four animals per condition: normoxic (control), 2 h anoxia exposed (acute anoxia) or 20 h anoxia exposed (chronic anoxia). Prior to introducing crayfish to their experimental containers, water was aerated by bubbling for 45 min with air for the control condition and with 100% nitrogen gas for both anoxic stress conditions as previously described (Lant and Storey, 2011). Anoxia was defined as≤1% O2 per 2 Torr (∼266.6 Pa). The crayfish were killed by
decapitation and tail muscle and hepatopancreas tissues were excised rapidly, flash frozen in liquid nitrogen and stored at−80°C until use.
Total RNA isolation
Tail muscle and hepatopancreas tissues fromn=4 different animals were weighed (100 mg), powdered in liquid nitrogen and homogenized in 1 ml of Trizol (cat. no. 15596-018; Invitrogen, Carlsbad, CA, USA) using a Polytron PT1200 homogenizer (Kinematica, Lucerne, Switzerland). Aliquots of 200 µl of chloroform were added prior to centrifugation at 10,000 rpm for 15 min at 4°C. The RNA-containing supernatant was extracted and precipitated on ice for 10 min using 500 µl of isopropanol. Samples were spun in a centrifuge at 12,000 rpm for 15 min at room temperature and the supernatant was then discarded. The RNA pellet was washed with 70% ethanol before spinning again at 7500 rpm for 5 min at room temperature. The supernatant was decanted, and pellets were air dried and then resuspended in 50 µl RNase-free water. Purity of the samples was ensured by verifying the absorbance ratio at 260 nm/280 nm using a Take3 micro-volume quantification plate (BioTek, Winooski, VT, USA) and PowerWave HT spectrophotometer (BioTek). Only samples with ratios between 1.8 and 2.2 were used. RNA integrity was verified by running samples on a 1% agarose gel stained with SYBR Green and examining the presence of sharp 28S and 18S ribosomal RNA bands. Samples were stored at−80°C until use.
Polyadenylation and stem-loop reverse transcription
RNA was prepared for miRNA analysis by polyadenylation, stem-loop primer ligation and reverse transcription, as described by Hadj-Moussa et al. (2016). The EpiBio PolyA tailing kit (cat. no. PAP5104H; Epicentre, Madison, WI, USA) was used for polyadenylation. For each reaction, 3 µg of RNA (1 µg µl−1) was
mixed with 1 µl of ATP (10 mmol l−1), 1μl of 10×E. colipoly A
polymerase buffer (0.1 mol l−1Tris-HCl pH 8.0, 0.25 mol l−1NaCl
and 10 mmol l−1MgCl
2) and 5 µl of RNase-free water for a final
volume of 10 µl. Polyadenylation was performed using an Eppendorf
thermocycler (Mississauga, ON, Canada) at 37°C for 30 min and reactions were terminated at 95°C for 5 min; samples were immediately chilled on ice. Specialized miRNA stem-loop primers (see Table S1 for sequence) were ligated by adding 5 µl of 250 pmol l−1stem-loop adapter primers to the polyadenylated RNA
samples and heated in a thermocycler at 95°C for 5 min to denature RNA, incubated at 60°C for 5 min to allow primer annealing and then immediately chilled on ice (Biggar et al., 2014). For reverse transcription, each polyadenylated and stem-loop ligated RNA sample was combined with 1 µl mouse Maloney leukaemia virus (M-MLV) reverse transcriptase (2 U; cat. no. 18080044; ThermoFisher Scientific, Waltham, MA, USA), 1 µl of 25 mmol l−1deoxynucleotide triphosphate (dNTP) mixture (cat. no.
R1121; ThermoFisher Scientific), 2 µl 0.1 mol l−1 dithiothreitol
(DTT) and 4 µl 5× first-strand buffer (cat. no. 18080044; ThermoFisher Scientific). Reverse transcription was initiated with 30 min of incubation at 16°C, 30 min at 42°C and 5 min at 85°C on a thermocycler. The cDNA samples were then serially diluted and stored at−20°C until use.
Relative miRNA quantification
Primers (Integrated DNA Technologies, Coralville, IA, USA) were designed based on DNA sequence alignments with 5–7 species including invertebrates such as the red swamp crayfish (Procambarus clarkia), the kuruma prawn (Marsupenaeus japonicus), the western honeybee (Apis mellifera), and vertebrates such as zebrafish (Danio rerio) and mouse (Mus musculus). miRNA targets were selected based on previously studied arrays inP. clarkia (Xu et al., 2014), where 75 were analysed in hepatopancreas and 71 were analysed in tail muscle. miRNA sequences were obtained from miRbase (Release 21, http://www.mirbase.org/). All miRNA-specific forward primer sequences, reference gene primers and the universal reverse primer are listed in Table S3. qPCR reactions were performed using a CFX Connect™ Real-Time PCR Detection System (cat. no. 1855201; Bio-Rad, Hercules, CA, USA), in accordance with MIQE guidelines (Bustin et al., 2009). Each 20 µl PCR reaction contained: 2 µl of diluted cDNA, 10.115 µl of ddH2O,
4 µl of 1 mol l−1trehalose (cat. no. TRE222; BioShop Canada Inc.,
Burlington, ON, Canada), 2 µl qPCR buffer (100 mmol l−1
Tris-HCl pH 8.5, 500 mmol l−1KCl, 1.5% Triton X-100, 20 mmol l−1
MgCl2, 2 mmol l−1dNTPs and 100 nmol l−1fluorescein), 0.5 µl
formamide (cat. no. FOR001; BioShop Canada Inc.), 0.5 µl of 25 mmol l−1miRNA-specific forward primer, 0.5 µl of 25 mmol l−1
universal reverse primer, 0.1 µl of 100× SYBR green mix diluted in dimethyl sulfoxide (DMSO) (cat. no. S7585; Invitrogen), 0.16 µl of 25 mmol l−1dNTPs and 0.125 µl of 5 U µl−1Taq polymerase (cat.
no. TAQ001.1; BioShop Canada Inc.). The miRNA amplification and quantification qPCR program used was as follows: an initial denaturation for 3 min at 95°C, then 40 cycles of 15 s at 95°C and 1 min at 60°C. A post-run melt curve analysis was performed to validate primer specificity; any reactions that displayed the amplification of more than one PCR product were rejected from our analyses and study.
miRNA binding site predictions
ovi-miR-33-5p binding site predictions were made using HIF1α mRNA sequences of four closely related invertebrate species – Litopenaeus vannamei, Palaemon pugio, Macrobrachium nipponenseand Scylla paramamosain–with SFold 2.2 STarMir bioinformatics software (http://sfold.wadsworth.org/cgi-bin/ starmirtest2.pl). ALG-1 CLIP (worm) was the model used for site
predictions.
Journal
of
Experimental
Relative mRNA transcript quantification
HIF1α mRNA primers were designed using DNA multi-species sequence alignments; species included: whiteleg shrimp (L. vannamei), daggerblade grass shrimp (P. pugio), oriental river prawn (M. nipponense) and green mud crab (S. paramamosain). See Table S4 for HIF1αand reference gene GAPDH primer sequences. qPCR reactions were carried out as described above (see‘Relative miRNA quantification’). The qPCR program used was as follows: an initial denaturation for 3 min at 95°C, then 49 cycles of 15 s at 95°C, 20 s at 57°C and 20 s at 72°C. A post-run melt curve analysis was used to validate primer specificity and single-product amplification.
Data analysis and statistics
Relative miRNA expression levels were calculated by the comparative ΔΔCqmethod. Raw Cqvalues were linearized (2−Cq) to standardize
against endogenous controls. The miRNA reference genes used were ribosomal 5S RNA in tail muscle and U6 spliceosomal RNA in the hepatopancreas. These targets were designated as controls because of their stable expression in control and stress conditions (Schmittgen and Livak, 2008). Data were analysed as mean relative expression (mean±s.e.m.), with n=4 independent biological replicates at each sampling point and two highly comparable technical replicates for each quantification. miRNA relative expression between control, 2 h anoxia and 20 h anoxia conditions was considered significantly different (P<0.05, P<0.01) when one-way ANOVA and Dunnett’s post hoc tests yielded P<0.05 using the RBIOPLOT statistics and graphing R package (Zhang and Storey, 2016).
Relative mRNA transcript abundance was also calculated by the comparativeΔΔCqmethod. Raw Cqvalues were linearized (2−Cq) to
standardize against endogenous controls. GAPDH was the designated reference gene because of its stable expression in control and stress conditions (Schmittgen and Livak, 2008). Data were analysed as mean relative expression (mean±s.e.m.), with n=4 independent biological replicates at each sampling point and two highly comparable technical replicates for each quantification. mRNA abundance between control, 2 h anoxia and 20 h anoxia conditions was considered significantly different (as above) when one-way ANOVA and Dunnett’s post hoc tests yielded P<0.05 using the RBIOPLOT statistics and graphing R package (Zhang and Storey, 2016).
RESULTS
Anoxic tail muscle miRNA quantification
In tail muscle tissue, 71 miRNAs were quantified under control (normoxic), acute 2 h anoxic and chronic 20 h anoxic conditions, and a total of four miRNAs were differentially expressed between the three conditions (Fig. 1). In the 2 h anoxic group, ovi-miR-81a-3pwas downregulated andovi-miR-3853-3pwas upregulated. In the 20 h anoxic condition, two miRNAs were upregulated: ovi-miR-2788-3p and ovi-miR-2812. For a complete list of miRNAs quantified (mean relative expression and s.e.m.) in muscle tissue, refer to Table S1.
Anoxic hepatopancreas miRNA quantification
In the hepatopancreas, an array of 75 miRNA targets were quantified under normoxic and 2 h and 20 h anoxic conditions, of which 21 showed significantly decreased expression under acute anoxia compared with the normoxic condition:ovi-bantam-3p, ovi-let-7-3p, ovi-miR-100-5p, ovi-miR-125-5p, ovi-miR-12-5p, ovi-miR-133-3p,ovi-miR-1-3p,ovi-miR-190-5p,ovi-miR-235-3p, ovi-miR-263a-5p,ovi-miR-276-5p,ovi-miR-276a-3p,ovi-miR-279-3p, ovi-miR-283-5p,ovi-miR-2944c-3p,ovi-miR-305-5p,
ovi-miR-307-3p,ovi- miR-317-3p,ovi-miR-33-5p,ovi-miR-750a-3pand ovi-miR-8-3p(Fig. 2). Of these 21 targets,ovi-bantam-3p,ovi-miR-133-3p, ovi-miR-1-3p, ovi-miR-263a-5p, ovi-miR-276-5p, ovi-miR-2944c-3p, ovi-miR-307-3p and ovi-miR-33-5p remained downregulated after 20 h anoxic exposure, while ovi-miR-375-3p became downregulated in addition to these after 20 h anoxic exposure. For a complete list of miRNAs quantified (mean relative expression and s.e.m.) in hepatopancreas tissue, refer to Table S2.
Bioinformatic analyses of pathways regulated by miRNA during anoxia
miRNAs in hepatopancreas that were significantly downregulated in O. virilis under 2 h anoxic exposure were analysed using the DIANA-microT-CDS algorithm (DIANA-miRPath v.3; Vlachos et al., 2015) for the identification of KEGG pathways that are targeted by these miRNAs. UsingDrosophila melanogasteras the reference species, DIANA-miRPath identified 12 of the 21 differentially expressed miRNA homologues,miR-1-3p,let-7-3p, miR-8-3p, miR-87-3p, miR-283-5p, miR-317-3p, miR-12-5p, miR-279-3p,miR-133-3p,miR-305-5p,miR-190-5p and bantam-3p, as regulators of the Hippo signalling pathway, targeting 17 genes within this pathway (P<0.001). In the hepatopancreas under 20 h anoxia, five out of nine downregulated miRNAs were found to target 12 genes within the Hippo pathway (P<0.001). Other pathways identified as potentially regulated (P<0.01) by these miRNAs include: mucin type O-glycan biosynthesis, glycerophospholipid metabolism, Jak-STAT signalling and MAPK signalling. In tail muscle, because of an absence of D. melanogaster homologues of the differentially regulated miRNAs, target enrichment analysis was not possible.
miRNA–mRNA binding site predictions
miR-33-5pwas predicted to bind to four mRNA transcript variants of HIF1α in closely related invertebrate species. Seedless-site binding was predicted in the 3′-UTR and the CDS of each transcript variant, as well as the 5′-UTR ofP. pugio,M. nipponenseandS. paramamosain. Fig. 3 illustrates a predicted binding site for miR-33-5pwith the 3′-UTR ofM. nipponenseHIF1αmRNA transcript variant.
Downstream HIF1αmRNA abundance levels
The relative transcript abundance of HIF1α was quantified in hepatopancreas under normoxic, and 2 h and 20 h anoxic conditions using qPCR. Transcripts levels were significantly (P<0.05) more abundant during 2 h anoxia exposure with a 1.51±0.03-fold increase, while levels were unchanged during 20 h anoxia exposure compared with control.
DISCUSSION
The ubiquitous role of miRNAs as regulators of cellular functions is widely accepted but their role in the stress response in naturally anoxia-tolerant organisms has not been deeply explored. Although exact mechanisms have yet to be deciphered, recent research has linked differential miRNA expression to various stress responses, including the anoxia response in adaptive invertebrates (Hadj-Moussa et al., 2018; Biggar et al., 2012; Lyons et al., 2015; Morin et al., 2008). In this study, 76 miRNAs were examined, and their potential involvement in coordinating the unique suite of adaptations required to survive and overcome anoxia-associated challenges was investigated. Both a tissue-specific and a stress-specific response was observed, where acute anoxia exposures and hepatopancreas tissues displayed more pronounced differential
Journal
of
Experimental
miRNA changes, while fewer changes were observed during both anoxia exposures in tail muscle tissue (Figs 1 and 2).
One central aspect of the stress response is the reprioritization of limited resources (Storey, 2015). Fig. 1 demonstrates this phenomenon in revealing few significant changes in tail muscle tissue miRNAs in response to acute and chronic anoxia. The hepatopancreas, a more metabolically active and vital organ, demonstrates considerable miRNA modulation (Fig. 2), suggesting that muscle tissue does not heavily rely on miRNA-mediated cytoprotective mechanisms to facilitate anoxia tolerance. Additionally, miRNA expression
signatures differed between acute and chronic anoxia in both tissues. As the animal acclimated to chronic stress conditions, fewer significant changes were observed (Fig. 2), probably because the acute response to insult includes a myriad of behavioural and physiological changes, such as increased ventilation, shunting of haemolymph to the anterior portion of the animal to promote oxygen delivery to sensitive tissues and, later in the response, decreased heart rate and increased stroke volume (Reiber, 1995). Oxygen-deprived environments have been shown to induce a preparatory response to oxidative stress, stimulating the upregulation of antioxidant defence mechanisms in a ovi-miR-81a-3p
2 h anoxia 20 h anoxia 2 h anoxia 20 h anoxia
ovi-bantam-3p
ovi-miR-2837 ovi-miR-2944c-3p ovi-miR-29-3p ovi-miR-2a-3p ovi-miR-305-3p ovi-miR-305-5p
ovi-miR-315-5p ovi-miR-316-5p ovi-miR-317-3p ovi-miR-3722
ovi-miR-3897-3p ovi-miR-48-5p ovi-miR-4942-3p ovi-miR-61-3p ovi-miR-6494-5p ovi-miR-71-5p ovi-miR-71-3p ovi-miR-7-3p ovi-miR-745-3p ovi-miR-750b-3p ovi-miR-750a-3p
ovi-miR-83-3p ovi-miR-8-3p ovi-miR-87-3p ovi-miR-965-3p ovi-miR-981-3p ovi-miR-993b-3p ovi-miR-993a-3p ovi-miR-996-3p ovi-miR-995-3p ovi-miR-9a-5p ovi-miR-9b/79-3p
Relative abundance
<0.5 1 >2
ovi-miR-285 ovi-miR-3853-3p ovi-miR-307-3p ovi-bantam-5p
ovi-let-7-5p ovi-let-7-3p ovi-miR-100-5p ovi-miR-1000-5p ovi-miR-10-3p ovi-miR-10-5p ovi-miR-1175-3p ovi-miR-125-5p ovi-miR-12-5p ovi-miR-133-3p ovi-miR-133-5p ovi-miR-137-3p ovi-miR-13b-3p
ovi-miR-184-3p ovi-miR-190-5p ovi-miR-193-3p ovi-miR-235-3p ovi-miR-252-5p ovi-miR-2568a-3p ovi-miR-263a-5p
ovi-miR-275-3p ovi-miR-2765-5p ovi-miR-276-5p ovi-miR-276a-3p ovi-miR-2779-5p ovi-miR-2788-3p ovi-miR-279-3p ovi-miR-2812 ovi-miR-281-2-5p ovi-miR-282-5p ovi-miR-283-5p ovi-miR-273 ovi-miR-1-3p
**
*
*
[image:4.612.47.383.55.612.2]*
Fig. 1. Heatmap of relative expression levels of 71Orconectes virilismicroRNAs (miRNAs) in muscle tissue under anoxia.miRNA expression was measured in muscle tissue of crayfish under 2 h and 20 h anoxic stress relative to that in normoxic conditions using quantitative PCR (qPCR). Data are means±s.e.m. of four independent biological replicates. Statistical analyses used a one-way ANOVA with Dunnett’s post hoctest: *P<0.05, **P<0.01. Refer to Table S1 for the relative expression ±s.e.m. values of all 76 miRNA species.
Journal
of
Experimental
diverse array of species (reviewed by Moreira et al., 2017). This may account, in part, for the differential stress response observed in the hepatopancreas (Fig. 2).
The differentially expressed hepatopancreas miRNAs were predicted to target stress-responsive pathways involved in maintaining homeostasis in response to changing oxygen concentrations. One such pathway that was predicted to be significantly affected by the downregulation of these miRNAs
during acute anoxia exposures in hepatopancreas was the Hippo signalling pathway. Hippo signalling is a highly conserved regulatory pathway of organ growth, cell proliferation and apoptosis that has recently been implicated in the hypoxia stress response (Ma et al., 2015; Reddy and Irvine, 2008). Moreover, the Hippo signalling promotes angiogenesis and proliferation and restricts tissue growth, while decreased signalling of this pathway promotes transcription of hypoxia-responsive genes, in part through the stabilization of HIF1α ovi-miR-81a-3p
2 h anoxia
20 h anoxia
20 h anoxia 2 h
anoxia
ovi-bantam-3p
ovi-miR-2944c-3p ovi-miR-29-3p ovi-miR-2a-3p ovi-miR-305-3p ovi-miR-305-5p
ovi-miR-315-5p ovi-miR-316-5p ovi-miR-317-3p ovi-miR-33-5p ovi-miR-34-5p ovi-miR-3722 ovi-miR-375-3p
ovi-miR-3897-3p ovi-miR-48-5p ovi-miR-4942-3p ovi-miR-61-3p ovi-miR-6494-5p ovi-miR-71-5p ovi-miR-71-3p ovi-miR-7-3p ovi-miR-745-3p ovi-miR-750b-3p ovi-miR-750a-3p
ovi-miR-83-3p ovi-miR-8-3p ovi-miR-87-3p ovi-miR-965-3p ovi-miR-981-3p ovi-miR-983b-3p ovi-miR-983a-3p ovi-miR-996-3p ovi-miR-995-3p ovi-miR-9a-5p ovi-miR-9b/79-3p
Relative abundance
<0.2 1 >1.5
ovi-miR-79-3p ovi-miR-3853-3p ovi-miR-307-3p ovi-bantam-5p
ovi-let-7-5p ovi-let-7-3p ovi-miR-100-5p ovi-miR-1000-5p ovi-miR-10-3p ovi-miR-10-5p ovi-miR-1175-3p ovi-miR-125-5p ovi-miR-12-5p ovi-miR-133-3p ovi-miR-133-5p ovi-miR-137-3p ovi-miR-13b-3p
ovi-miR-184-3p ovi-miR-190-5p ovi-miR-193-3p ovi-miR-235-3p ovi-miR-252-5p ovi-miR-2568a-3p ovi-miR-263a-5p
ovi-miR-275-3p ovi-miR-2765-5p
ovi-miR-276a-3p ovi-miR-276-5p
ovi-miR-2779-5p ovi-miR-2788-3p ovi-miR-279-3p ovi-miR-279b-3p ovi-miR-2812 ovi-miR-281-2-5p ovi-miR-282-5p ovi-miR-283-5p ovi-miR-2837 ovi-miR-273 ovi-miR-1-3p
**
*
*
*
*
**
**
**
***
**
*
*
*
*
**
**
**
**
*
*
*
*
**
**
**
**
*
*
*
[image:5.612.50.381.56.630.2]*
Fig. 2. Heatmap of relative expression levels of 75O. virilismiRNAs in hepatopancreas tissue under anoxia.miRNA expression was measured in hepatopancreas tissue of crayfish under 2 h and 20 h anoxic stress relative to that in normoxic conditions using qPCR. Data are means±s.e.m. of four independent biological replicates. Statistical analyses used a one-way ANOVA with Dunnett’s post hoctest: *P<0.05, **P<0.01, ***P<0.001. Refer to Table S2 for the relative expression ±s.e.m. values of all 76 miRNA species.
Journal
of
Experimental
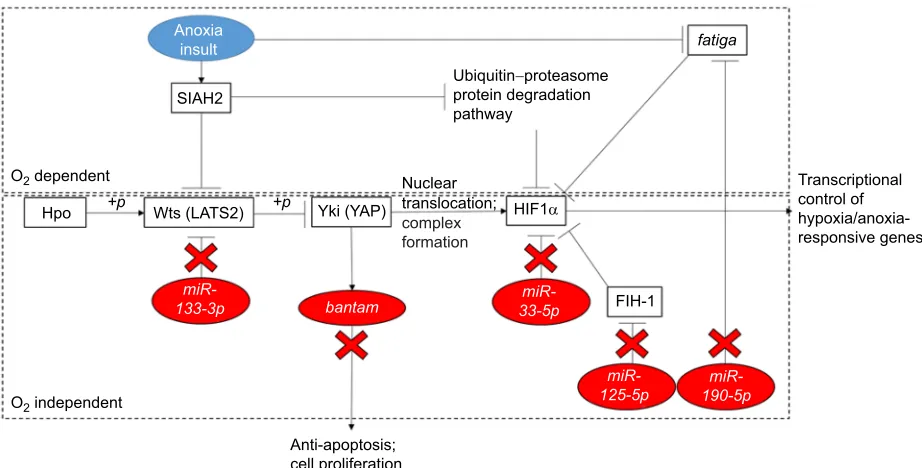
(Meng et al., 2016; Morin et al., 2005). Hippo signalling has been researched extensively in both mice and fruit flies, where the core components of the pathway are conserved between vertebrates and invertebrates, including the kinases Hippo (Hpo) and Warts (Wts), and cofactors Salvador (Sav) and Yorkie (Yki) (Halder and Johnson, 2011; Huang et al., 2005; Harvey et al., 2003; Pantalacci et al., 2003; Udan et al., 2003; Wu et al., 2003). Fig. 4 illustrates the transcriptional regulation occurring through this kinase cascade, in which Hpo activates Wts through phosphorylation, which in turn phosphorylates and thus inactivates Yki, resulting in the inhibition of transcription of cell-cycle and cell death regulators (Huang et al., 2005).miR-133-3p was predicted to regulate Wts expression as in Fig. 4 and, as indicated in Fig. 2, this miRNA was downregulated in anoxic hepatopancreas tissue. This suggests the increased activity of Wts and the subsequent inhibition of Yki. It has been shown that overexpression of Yki results in defective apoptosis, cell growth and proliferation (Huang et al., 2005). Additionally, the same phenotype has been observed in systems with overexpression of the miRNA bantam, which was shown to be downregulated in hepatopancreas tissues under anoxic
conditions (Figs 1 and 2) (Nolo et al., 2006). The gene encoding the bantam miRNA is activated by Yki, further suggesting that cell growth and proliferation are arrested in the hepatopancreas during oxygen stress (Nolo et al., 2006; Thompson and Cohen, 2006). The predicted overexpression of Yki via miRNA modulation suggests that cell growth and proliferation are simultaneously arrested to promote energy conservation in the hepatopancreas of the northern crayfish under acute anoxic stress (Zhou et al., 2015).
There are multiple inputs regulating HIF, but the role of hypoxic stress is of great interest; hypoxia was found to deactivate this signalling cascade through ubiquitination and protein destabilization (Ma et al., 2015; Nakayama et al., 2004). Consequently, YAP, the vertebrate orthologue of Yki, is stimulated by this process and is translocated to the nucleus to form a stable complex with HIF1α(Ma et al., 2015). HIF1 plays a critical role in the hypoxia response through transcriptional regulation of a variety of processes including metabolic adaptation, vascular remodelling, angiogenesis and cell survival (Nakayama et al., 2004; Bendinelli et al., 2013; Pugh and Ratcliffe, 2003; Semenza, 1998). Additionally, HIF1αis targeted by miRNAs that were found to be differentially regulated during anoxia (Fig. 2). The HIF1 heterodimer is composed of alpha and beta subunits, the beta subunits being constitutively expressed and the alpha subunits having a short half-life in normoxic conditions, resulting in saturated expression of the inactive protein under standard physiological conditions (Nakayama et al., 2004; Pugh and Ratcliffe, 2003). Under normoxic conditions, rapid turnover of HIF1αoccurs through the ubiquitin–proteasome protein degradation pathway (Kallio et al., 1999; Salceda and Caro, 1997). HIF1α stability is determined in large part by cellular oxygen concentration but also by the YAP/Yki protein, which forms a stable complex with the α-subunit (Nakayama et al., 2004; Huang et al., 1998). In particular, and as reflected in Fig. 2, three differentially expressed miRNAs quantified in this study,miR-33-5p,miR-125-5pandmiR-190-5, were shown to directly or indirectly target the alpha subunit of the HIF1
|
C G U U G A UG U
U
A C G UA
UG C A A UA G
UA A
U A C A U G C A U
U 3⬘
−−−
5⬘
5⬘
−−−3⬘
4609 4627
ovi-miR-33-5p
[image:6.612.71.277.59.145.2]G=−15.5 kcal mol–1
Fig. 3. Predicted seedless-site binding of miR-33-5p withMacrobrachium nipponensehypoxia inducible factor-1α(HIF1α) mRNA transcript variant.
Image generated bySFold 2.2 (see Materials and Methods). Nucleotides in red indicate seed-region (nucleotides 2–7) of miR-33-5p. Logistical probability of binding was reported at 0.489 whereΔG=−15.5 kcal mol−1.
miR-133-3p
Anoxia insult
SIAH2
Hpo
FIH-1
Wts (LATS2) Yki (YAP) HIF1α
O2 dependent
O2 independent
fatiga
Ubiquitin−proteasome protein degradation pathway
Anti-apoptosis; cell proliferation
Transcriptional control of hypoxia/anoxia-responsive genes Nuclear
translocation; complex formation
+p +p
miR-33-5p
miR-125-5p 190-5p miR-bantam
Fig. 4. Schematic diagram of miRNA regulation of the Hippo signalling pathway with multiple inputs and key interactions with HIF1α.Vertebrate orthologue proteins are shown in parentheses. Adapted in part from miRPath v.3 KEGG pathway (see Materials and Methods). WithDrosophila melanogaster used as the reference species, 12 of the 21 differentially expressed miRNA homologues,miR-1-3p,let-7-3p,miR-8-3p,miR-87-3p,miR-283-5p,miR-317-3p, miR-12-5p,miR-279-3p,miR-133-3p,miR-305-5p,miR-190-5p and bantam-3p, were identified as statistically significant regulators of the Hippo signalling
pathway (P=8.70e−07).
Journal
of
Experimental
[image:6.612.76.537.447.681.2]master regulator of oxygen stress (De Lella Ezcurra et al., 2016; Li et al., 2016; Zhou et al., 2015).
There is strong evidence to suggest that HIF1αis a direct target of miR-33-5pand, as such, is inhibited bymiR-33-5pexpression (Zhou et al., 2015). Results from binding site analysis indicated that ovi-miR-33-5phas potential seedless-site binding in the 3′-UTR of invertebrate HIF1α mRNA (Fig. 4). It was recently proposed that seedless-site binding of miRNAs may account for as much as 40% of total miRNA–mRNA binding in some instances (Loeb et al., 2012). Fig. 2 shows decreased abundance of miR-33-5p in the hepatopancreas in response to acute and chronic anoxia, suggesting an active role for HIF1αin the sustained regulation of anoxia in this tissue. One recent study found that decreased levels ofmiR-33-5p resulted in increased proliferation, and conversely overexpression of this miRNA resulted in decreased proliferation of certain cell lines (Zhou et al., 2015). miRNAs have been studied for their vast role in the simultaneous arrest of cell proliferation and apoptosis in metabolically depressed animals (Biggar and Storey, 2011). We hypothesize that this mechanism, in conjunction with other elements of the Hippo signalling pathway, may be activated in anoxic crayfish to regulate proliferation and inhibit apoptosis in critical organs. To avoid fatal ATP depletion, animals under reduced metabolic states must arrest energetically expensive processes such as proliferation and growth (Biggar and Storey, 2011). It was also speculated that other animal models of anoxia tolerance exhibit regulation of apoptosis and proliferation under stress via miRNA control (Morin et al., 2008; Biggar et al., 2009). Energy metabolism and cell survival are among the responses regulated by HIF1; therefore, it follows that this transcription factor would be regulated during anoxia, as was previously observed in studies on metabolically depressed animals (Morin et al., 2005; Maistrovski et al., 2012).
Interestingly, miR-190-5, which has the opposite effect on HIF1α, was shown to decrease significantly only under acute anoxia in the hepatopancreas (Fig. 2). The oxygen-dependent degradation of HIF1α occurs through prolyl hydroxylation by catalytic enzymes that use oxygen as a co-substrate under normoxic conditions (Jaakkola et al., 2001). This mechanism is further regulated by miRNAs and, specifically,miR-190has been identified as targeting the prolyl hydroxylase fatiga, an essential negative regulator of hypoxic responses, and was found to be overexpressed in response to hypoxia (De Lella Ezcurra et al., 2016). One explanation proposed for the unexpected decreased abundance of this positive HIF1αregulator is that while the study by De Lella Ezcurra et al. (2016) examined the response to hypoxia (11%, 8% and 5% oxygen), the present study examined the response to anoxia (≤1% O2 per 2 Torr). Furthermore, because the fatiga gene is
inactivated by anoxia, the additional negative regulation bymiR-190 may not serve an energetic benefit to the cell if HIF1αwill not be degraded under anoxia, regardless of the abundance of the prolyl hydroxylase enzyme.
miR-125-5pis known to target HIF1αindirectly, where decreased levels of this miRNA result in the inhibition of proliferation and activation of hepatic stellate cells (Li et al., 2016). This miRNA binds to factor inhibiting HIF1 (FIH-1), which in turn accumulates and negatively regulates HIF1αtranscriptional activity (Li et al., 2016; Mahon et al., 2001). The complex repression of HIF1αby FIH-1 and its co-repressor VHL has been characterized in normoxic conditions; however, these novel findings suggest that, under anoxia, a different mechanism of expression is engaged (Mahon et al., 2001). Because miR-125-5p levels were shown to decrease in the hepatopancreas under acute anoxia (Fig. 2), FIH-1 levels were expected to increase, and thus increase the negative regulation of HIF1α.
Transcript levels of HIF1α have previously been shown to increase in response to low-oxygen stress in an anoxia-tolerant insect (Morin et al., 2005). The activation of HIF1α in another invertebrate suggests the upregulation ofhif1awe measured during acute anoxia in crayfish hepatopancreas is a potentially conserved response for the adaptation to anoxia-associated challenges. Previous studies have observed that HIF1α transcript abundance peaked between 1 and 2 h exposure but decreased to near-basal levels after 8 h exposure before finally increasing again following 16 h anoxia exposure (Wang et al., 1995). This agrees with the present findings of HIF1αmRNA abundance over the anoxia time course. These findings also reinforce the hypothesis that the HIF1 mechanism of regulating anoxia-responsive genes is highly conserved throughout the animal kingdom and is under miRNA regulation (Morin et al., 2005; Maistrovski et al., 2012; Gorr, 2004). In accordance with the findings of Morin et al. (2005), the increased transcript levels ofhif1aalso suggest that the transcription factor activation is promoted both through the increased transcription of the hif1α gene and simultaneously, the inhibition of HIF1α degradation.
We also foundovi-miR-133-5pto be significantly downregulated in the hepatopancreas, emphasizing that anti-apoptotic mechanisms are activated in this vital organ under anoxia. Indeed, studies have shown that miR-133 promotes hypoxia-induced cell death through the suppression of anti-apoptotic proteins while promoting the activation of pro-apoptotic proteins (Li et al., 2015). Also downregulated under anoxia stress in the hepatopancreas aremiR-263,miR-276and miR-317, all of which are known to be upregulated at various stages of development among invertebrates (Surridge et al., 2011). This suggests that these miRNAs are downregulated in the hepatopancreas to suppress cell proliferation and growth while the animal is in an anoxia-induced metabolically depressed state.
In contrast to the hepatopancreas, tail muscle may experience a smaller effect of anoxia insult as muscle tissue is less oxygen sensitive compared with hepatopancreas and therefore has a lesser effect on the survivorship of the animal (Reiber, 1995). This is a potential explanation for the total of four differentially expressed miRNAs during anoxia in tail muscle, three of which exhibited upregulation and one of which exhibited downregulation (Fig. 1). The role of these four miRNAs has not yet been characterized and there are currently no known homologues in D. melanogaster. However, miR-2788 has been previously identified in an invertebrate, exhibiting upregulation during development, so a similar suppression in proliferation and cell growth to that proposed in hepatopancreas is also proposed in tail muscle (Surridge et al., 2011). Presently, there is a lack of functional targeting information available for these miRNAs and this is probably due to their crustacean-specific functions that have yet to be explored deeply.
Overall, this study elucidates the central and newfound role of miRNAs as modulators of gene translation during anoxia exposure. Indeed, these findings highlight the tissue-specific nature of miRNA targeting, where pronounced responses were observed in the hepatopancreas under acute anoxic stress. This suggests that more metabolically active tissues such as the hepatopancreas are more dependent on miRNA-mediated anoxia responses than the metabolically inactive muscle, and that a rapid response during acute anoxia exposure is required to provide immediate protection against anoxia insult. Key to the hepatopancreas response is the regulation of Hippo signalling, which modulates proliferation and apoptotic pathways, and the HIF1 transcription factor, which was found to be upregulated during acute anoxia and was also targeted by a variety of miRNAs itself. These novel findings inO. virilishave
Journal
of
Experimental
major implications for how the anoxia response modulates–and is modulated by–miRNAs in aquatic invertebrates.
Acknowledgements
We thank Jan Storey for her scientific insight and editorial review of the manuscript.
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: H.H.-M.; Methodology: S.G.E., H.H.-M.; Software: K.B.S.; Investigation: S.G.E.; Resources: K.B.S.; Data curation: S.G.E., H.H.M.; Writing -original draft: S.G.E.; Writing - review & editing: S.G.E., H.H.-M., K.B.S.; Supervision: H.H.-M., K.B.S.; Project administration: H.H.-M., K.B.S.; Funding acquisition: K.B.S.
Funding
This work was supported by a Discovery grant from the Natural Sciences and Engineering Research Council of Canada (NSERC) (no. 6793) to K.B.S. K.B.S. holds the Canada Research Chair in Molecular Physiology. H.H.-M. holds a Natural Sciences and Engineering Research Council of Canada PhD post-graduate scholarship.
Supplementary information
Supplementary information available online at
http://jeb.biologists.org/lookup/doi/10.1242/jeb.190579.supplemental
References
Al-attar, R., Zhang, Y. and Storey, K. B.(2017). Osmolyte regulation by TonEBP/ NFAT5 during anoxia-recovery and dehydration–rehydration stresses in the freeze-tolerant wood frog (Rana sylvatica).PeerJ5, e2797.
Bendinelli, P., Maroni, P., Matteucci, E., Luzzati, A., Perrucchini, G. and Desiderio, M. A.(2013). Hypoxia inducible factor-1 is activated by transcriptional co-activator with PDZ-binding motif (TAZ) versus WWdomain-containing oxidoreductase (WWOX) in hypoxic microenvironment of bone metastasis from breast cancer.Eur. J. Cancer.49, 2608-2618.
Biggar, K. K. and Storey, K. B.(2011). The emerging roles of microRNAs in the molecular responses of metabolic rate depression.J. Mol. Cell Biol.3, 167-175.
Biggar, K. K. and Storey, K. B.(2017). Exploration of low temperature microRNA function in an anoxia tolerant vertebrate ectotherm, the red eared slider turtle (Trachemys scripta elegans).J. Therm. Biol.68, 139-146.
Biggar, K. K., Dubuc, A. and Storey, K.(2009). MicroRNA regulation below zero: Differential expression of miRNA-21 and miRNA-16 during freezing in wood frogs. Cryobiology59, 317-321.
Biggar, K. K., Kornfeld, S. F., Maistrovski, Y. and Storey, K. B.(2012). MicroRNA regulation in extreme environments: differential expression of microRNAs in the intertidal snail Littorina littorea during extended periods of freezing and anoxia. Genomics. Proteomics Bioinformatics.10, 302-309.
Biggar, K. K., Wu, C.-W. and Storey, K. B.(2014). High-throughput amplification of mature microRNAs in uncharacterized animal models using polyadenylated RNA and stem–loop reverse transcription polymerase chain reaction.Anal. Biochem. 462, 32-34.
Bradford, D. F.(1983). Winterkill, oxygen relations, and energy metabolism of a submerged dormant amphibian,Rana muscosa.Ecology64, 1171-1183. Bustin, S. A., Benes, V., Garson, J. A., Hellemans, J., Huggett, J., Kubista, M.,
Mueller, R., Nolan, T., Pfaffl, M. W., Shipley, G. L. et al.(2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments.Clin. Chem.55, 611-622.
Christopher, A., Kaur, R., Kaur, G., Kaur, A., Gupta, V. and Bansal, P.(2016). MicroRNA therapeutics: Discovering novel targets and developing specific therapy.Perspect. Clin. Res.7, 68.
Cowan, K. J., Storey, K. B. and Storey, K. B. (2001). Protein kinase and phosphatase responses to anoxia in crayfish, Orconectes virilis: purification and characterization of cAMP-dependent protein kinase.Comp. Biochem. Physiol. Part B.130, 565-577.
Dawson, N. J. and Storey, K. B.(2011). Regulation of tail muscle arginine kinase by reversible phosphorylation in an anoxia-tolerant crayfish.J. Comp. Physiol. B. 181, 851-859.
De Lella Ezcurra, A. L., Bertolin, A. P., Kim, K., Katz, M. J., Gándara, L., Misra, T., Luschnig, S., Perrimon, N., Melani, M. and Wappner, P. (2016). miR-190 enhances HIF-dependent responses to hypoxia in drosophila by inhibiting the Prolyl-4-hydroxylase Fatiga.PLoS Genet.12, e1006073.
Gerber, V. E. M., Wijenayake, S. and Storey, K. B.(2016). Anti-apoptotic response during anoxia and recovery in a freeze-tolerant wood frog (Rana sylvatica).PeerJ. 4, e1834.
Gorr, T. A.(2004). Daphnia and Drosophila: two invertebrate models for O2 responsive and HIF-mediated regulation of genes and genomes.Int. Congr. Ser. 1275, 55-62.
Green, S. R. and Storey, K. B.(2016). Regulation of crayfish, Orconectes virilis, tail muscle lactate dehydrogenase (LDH) in response to anoxic conditions is associated with alterations in phosphorylation patterns. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol.202, 67-74.
Hadj-Moussa, H., Moggridge, J. A., Luu, B. E., Quintero-Galvis, J. F., Gaitá n-Espitia, J. D., Nespolo, R. F. and Storey, K. B.(2016). The hibernating South American marsupial, Dromiciops gliroides, displays torpor-sensitive microRNA expression patterns.Sci. Rep.6, 24627.
Hadj-Moussa, H., Logan, S. M., Seibel, B. A. and Storey, K. B.(2018). Potential role for microRNA in regulating hypoxia-induced metabolic suppression in jumbo squids.Biochim. Biophys. Acta Gene Regul. Mech.1861, 586-593.
Halder, G. and Johnson, R. L.(2011). Hippo signaling: growth control and beyond. Development138, 9-22.
Harvey, K. F., Pfleger, C. M. and Hariharan, I. K.(2003). The Drosophila Mst ortholog, hippo, restricts growth and cell proliferation and promotes apoptosis.Cell 114, 457-467.
Hochachka, P. W. and Lutz, P. L.(2001). Mechanism, origin, and evolution of anoxia tolerance in animals.Comp. Biochem. Physiol. B. Biochem. Mol. Biol.130, 435-459.
Huang, L. E., Gu, J., Schau, M. and Bunn, H. F.(1998). Regulation of hypoxia-inducible factor 1alpha is mediated by an O2-dependent degradation domain via the ubiquitin-proteasome pathway.Proc. Natl. Acad. Sci. USA95, 7987-7992. Huang, J., Wu, S., Barrera, J., Matthews, K. and Pan, D.(2005). The hippo
signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating yorkie, the drosophila homolog of YAP.Cell122, 421-434. Jaakkola, P., Mole, D. R., Tian, Y.-M., Wilson, M. I., Gielbert, J., Gaskell, S. J.,
Kriegsheim, A. v., Hebestreit, H. F., Mukherji, M., Schofield, C. J. et al.(2001). Targeting of HIF-alpha to the von hippel-lindau ubiquitylation complex by O2-regulated prolyl hydroxylation.Science292, 468-472.
Kallio, P. J., Wilson, W. J., O’Brien, S., Makino, Y. and Poellinger, L.(1999). Regulation of the hypoxia-inducible transcription factor 1alpha by the ubiquitin-proteasome pathway.J. Biol. Chem.274, 6519-6525.
Kornfeld, S. F., Biggar, K. K. and Storey, K. B.(2012). Differential expression of mature microRNAs involved in muscle maintenance of hibernating little brown bats, myotis lucifugus: a model of muscle atrophy resistance, genomics. Proteomics Bioinformatics.10, 295-301.
Krivoruchko, A. and Storey, K. B.(2010). Molecular mechanisms of turtle anoxia tolerance: A role for NF-κB.Gene450, 63-69.
Krivoruchko, A. and Storey, K. B.(2015). Turtle anoxia tolerance: Biochemistry and gene regulation.Biochim. Biophys. Acta Gen. Subj.1850, 1188-1196. Lant, B. and Storey, K. B.(2011). Glucose-6-phosphate dehydrogenase regulation
in anoxia tolerance of the freshwater crayfish orconectes virilis.Enzyme Res. 2011, 524906.
Li, A., Yang, Q. and Yang, K.(2015). miR-133a mediates the hypoxia-induced apoptosis by inhibiting TAGLN2 expression in cardiac myocytes.Mol. Cell. Biochem.400, 173-181.
Li, G., Li, J., Li, C., Qi, H., Dong, P., Zheng, J. and Yu, F.(2016). MicroRNA-125a-5p contributes to hepatic stellate cell activation through targeting FIH1.Cell. Physiol. Biochem.38, 1544-1552.
Loeb, G. B., Khan, A. A., Canner, D., Hiatt, J. B., Shendure, J., Darnell, R. B., Leslie, C. S. and Rudensky, A. Y.(2012). Transcriptome-wide miR-155 binding map reveals widespread noncanonical microRNA targeting. Mol. Cell. 48, 760-770.
Lund, E., Güttinger, S., Calado, A., Dahlberg, J. E. and Kutay, U.(2004). Nuclear Export of MicroRNA Precursors.Science303, 95-98.
Lyons, P. J., Crapoulet, N., Storey, K. B. and Morin, P. J.(2015). Identification and profiling of miRNAs in the freeze-avoiding gall moth Epiblema scudderiana via next-generation sequencing.Mol. Cell. Biochem.410, 155-163.
Ma, B., Chen, Y., Chen, L., Cheng, H., Mu, C., Li, J., Gao, R., Zhou, C., Cao, L., Liu, J. et al.(2015). Hypoxia regulates Hippo signalling through the SIAH2 ubiquitin E3 ligase.Nat. Cell Biol.17, 95-103.
Mahon, P. C., Hirota, K. and Semenza, G. L.(2001). FIH-1: a novel protein that interacts with HIF-1alpha and VHL to mediate repression of HIF-1 transcriptional activity.Genes Dev.15, 2675-2686.
Maistrovski, Y., Biggar, K. K. and Storey, K. B.(2012). HIF-1αregulation in mammalian hibernators: role of non-coding RNA in HIF-1αcontrol during torpor in ground squirrels and bats.J. Comp. Physiol. B.182, 849-859.
Mattice, A. M. S., MacLean, I. A., Childers, C. L. and Storey, K. B.(2018). Characterization of pyruvate kinase from the anoxia tolerant turtle,Trachemys scripta elegans: a potential role for enzyme methylation during metabolic rate depression.PeerJ6, e4918.
Maziere, P. and Enright, A. J.̀ (2007). Prediction of microRNA targets.Drug Discov. Today.12, 452-458.
Meng, Z., Moroishi, T. and Guan, K.-L.(2016). Mechanisms of Hippo pathway
regulation.Genes Dev.30, 1-17.
Journal
of
Experimental
Moreira, D. C., Oliveira, M. F., Liz-Guimarães, L., Diniz-Rojas, N., Campos, É. G. and Hermes-Lima, M.(2017). Current trends and research challenges regarding “preparation for oxidative stress.Front. Physiol.8, 702.
Morin, P. J., Mcmullen, D. C. and Storey, K. B.(2005). HIF-1αinvolvement in low temperature and anoxia survival by a freeze tolerant insect.Mol. Cell. Biochem. 280, 99-106.
Morin, P. J., Dubuc, A. and Storey, K. B. (2008). Differential expression of microRNA species in organs of hibernating ground squirrels: a role in translational suppression during torpor.Biochim. Biophys. Acta Gene Regul. Mech.1779, 628-633.
Muir, T. J., Costanzo, J. P. and Lee, R. E. (2007). Osmotic and metabolic responses to dehydration and urea-loading in a dormant, terrestrially hibernating frog.J. Comp. Physiol. B.177, 917-926.
Nakayama, K., Frew, I. J., Hagensen, M., Skals, M., Habelhah, H., Bhoumik, A., Kadoya, T., Erdjument-Bromage, H., Tempst, P., Frappell, P. B. et al.(2004). Siah2 regulates stability of prolyl-hydroxylases, controls HIF1αabundance, and modulates physiological responses to hypoxia.Cell117, 941-952.
Nolo, R., Morrison, C. M., Tao, C., Zhang, X. and Halder, G.(2006). The bantam MicroRNA is a target of the hippo tumor-suppressor pathway.Curr. Biol.16, 1895-1904.
Pantalacci, S., Tapon, N. and Léopold, P.(2003). The Salvador partner Hippo promotes apoptosis and cell-cycle exit in Drosophila.Nat. Cell Biol.5, 921-927. Pugh, C. W. and Ratcliffe, P. J.(2003). The von Hippel-Lindau tumor suppressor,
hypoxia-inducible factor-1 (HIF-1) degradation, and cancer pathogenesis.Semin. Cancer Biol.13, 83-89.
Reddy, B. V. V. G. and Irvine, K. D.(2008). The Fat and Warts signaling pathways: new insights into their regulation, mechanism and conservation.Development 135, 2827-2838.
Reiber, C. L. (1995). Physiological adaptations of crayfish to the hypoxic environment.Am. Zool.35, 1-11.
Salceda, S. and Caro, J.(1997). Hypoxia-inducible factor 1alpha (HIF-1alpha) protein is rapidly degraded by the ubiquitin-proteasome system under normoxic conditions. Its stabilization by hypoxia depends on redox-induced changes. J. Biol. Chem.272, 22642-22647.
Schmittgen, T. D. and Livak, K. J.(2008). Analyzing real-time PCR data by the comparative C(T) method.Nat. Protoc.3, 1101-1108.
Schueler, F. and Karstad, A.(1996). Interim report on the functions and status of kemptville creek, rideau valley conservation authority.Manotick.
Seibel, B. A., Luu, B. E., Tessier, S. N., Towanda, T. and Storey, K. B.(2018). Metabolic suppression in the pelagic crab, Pleuroncodes planipes, in oxygen minimum zones.Comp. Biochem. Physiol. Part B Biochem. Mol. Biol.224, 88-97. Semenza, G. L. (1998). Hypoxia-inducible factor 1: master regulator of O2
homeostasis.Curr. Opin. Genet. Dev.8, 588-594.
Storey, K. B. (2015). Regulation of hypometabolism: insights into epigenetic controls.J. Exp. Biol.218, 150-159.
Storey, K. B. and Storey, J. M.(1990). Metabolic rate depression and biochemical adaptation in anaerobiosis, hibernation and, source.Q. Rev. Biol.65, 145-174. Surridge, A. K., Lopez-Gomollon, S., Moxon, S., Maroja, L. S., Rathjen, T.,
Nadeau, N. J., Dalmay, T. and Jiggins, C. D.(2011). Characterisation and expression of microRNAs in developing wings of the neotropical butterfly Heliconius melpomene.BMC Genomics.12, 62.
Szereszewski, K. E. and Storey, K. B.(2018). Translational regulation in the anoxic turtle, Trachemys scripta elegans.Mol. Cell. Biochem.445, 13-23.
Thompson, B. J. and Cohen, S. M.(2006). The hippo pathway regulates the bantam microRNA to control cell proliferation and apoptosis in drosophila.Cell 126, 767-774.
Udan, R. S., Kango-Singh, M., Nolo, R., Tao, C. and Halder, G.(2003). Hippo promotes proliferation arrest and apoptosis in the Salvador/Warts pathway.Nat. Cell Biol.5, 914-920.
Vlachos, I. S., Zagganas, K., Paraskevopoulou, M. D., Georgakilas, G., Karagkouni, D., Vergoulis, T., Dalamagas, T. and Hatzigeorgiou, A. G. (2015). DIANA-miRPath v3.0: deciphering microRNA function with experimental support.Nucleic Acids Res.43, W460-466.
Wang, G. L., Jiang, B.-H., Rue, E. A. and Semenza, G. L.(1995). Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension (dioxin receptor/erythropoietin/hypoxia/transcription).Genetics 92, 5510-5514.
Wijenayake, S., Hawkins, L. J. and Storey, K. B.(2018). Dynamic regulation of six histone H3 lysine (K) methyltransferases in response to prolonged anoxia exposure in a freshwater turtle.Gene649, 50-57.
Wu, S., Huang, J., Dong, J. and Pan, D.(2003). hippo encodes a Ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts.Cell114, 445-456.
Xu, W. N., Bin Liu, W., Yang, W. W., Zhang, D. D. and Jiang, G. Z.(2014). Identification and differential expression of hepatopancreas microRNAs in red swamp crayfish fed with emodin diet.Fish Shellfish Immunol.39, 1-7. Zhang, J. and Storey, K. B.(2016). RBioplot: an easy-to-use R pipeline for
automated statistical analysis and data visualization in molecular biology and biochemistry.PeerJ.4, e2436.
Zhou, J., Xu, D., Xie, H., Tang, J., Liu, R., Li, J., Wang, S., Chen, X., Su, J., Zhou, X. et al.(2015). miR-33a functions as a tumor suppressor in melanoma by targeting HIF-1α.Cancer Biol. Ther.16, 846-855.
Ziello, J. E., Jovin, I. S. and Huang, Y.(2007). Hypoxia-Inducible Factor (HIF)-1 regulatory pathway and its potential for therapeutic intervention in malignancy and ischemia.Yale J. Biol. Med.80, 51-60.