STUDIES ON THE PHYSIOLOGY OF
ARENICOLA MARINA L.
I. THE PACE-MAKER ROLE OF THE OESOPHAGUS, AND THE ACTION OF ADRENALINE AND ACETYLCHOLINE
BY G. P. WELLS
Department of Zoology, University College, London
(Received 19 September 1936)
(With One Plate and Twenty-six Text-figures) CONTENTS
PAGE I. Introduction 117 II. T h e activity of the isolated oesophagus . . . . 1 1 8 III. T h e invasion of the proboscis by oesophageal rhythms 121 (a) T h e nomenclature of the parts of the extrovert 121 (b) The isolated extrovert preparation 122 (c) Behaviour of the isolated extrovert in sea water 123 (d) B e h a v i o u r o f t h e i s o l a t e d e x t r o v e r t i n b o d y fluid . '. . . . . 1 2 4 ( e ) A d r e n a l i n e , a c e t y l c h o l i n e a n d t h e i s o l a t e d e x t r o v e r t . . . 1 2 7 ( / ) S i t e o f o r i g i n o f t h e a c t i v i t y o f t h e i s o l a t e d e x t r o v e r t . . . 1 3 0 (g) S i t e o f a c t i o n o f a d r e n a l i n e a n d a c e t y l c h o l i n e . . . 1 3 6 I V . T h e i n v a s i o n o f t h e c e n t r a l n e r v o u s s y s t e m a n d b o d y w a l l b y o e s o p h a g e a l r h y t h m s . 1 3 8 (a) T h e e x t r o v e r t : b o d y - w a l l p r e p a r a t i o n 1 3 8 ( 6 ) B e h a v i o u r o f t h e p r e p a r a t i o n i n s e a w a t e r 1 3 9 ( c ) T h e a c t i o n o f a d r e n a l i n e a n d a c e t y l c h o l i n e 1 4 2 V . O e s o p h a g e a l r h y t h m s i n t h e i n t a c t a n i m a l 1 4 3 (a) M e t h o d s 1 4 4 ( 6 ) T h e r e s t i n g r h y t h m ' 1 4 5 ( c ) T h e a c t i o n o f a d r e n a l i n e . . . 1 4 7 V I . D i s c u s s i o n . 1 5 0 S u m m a r y . . . 1 5 4 A c k n o w l e d g e m e n t s . . . 1 5 7 R e f e r e n c e s . . . . 1 5 7
I. INTRODUCTION
THE following communication contributes to our knowledge of two distinct aspects of annelid physiology.
(1) Since the work of Quatrefages (1844), it has been known that the polychaetes possess a system of nerves and ganglia, often very elaborate, in the walls of the
n 8 G. P. WELLS
eversible proboscis. Although the structure of the stomatogastric nervous system has been described in a number of different species, nothing has yet been published on its physiology. The few investigators who have studied the polychaete nervous system experimentally have confined their attention to the supra- and sub-oeso-phageal ganglia and the ventral nerve cord. The experiments here described were carried out on Arenicola marina, a species which, because of its abundance and large size, is well suited for experimental work, but in which, unhappily, the anatomy of the stomatogastric nervous system has not yet been described. Nevertheless, they throw a certain amount of light on the interrelations between the neuromuscular structures in the front part of the gut and those of the body wall of the anterior end. It is shown below that the oesophageal region of Arenicola exerts a remarkable pace-maker action, discharging impulses forwards, via the proboscis, into the central nervous system, and thus determining certain movements of the whole animal. This point is established by considering one by one a series of preparations of ascending complexity, starting with the isolated oesophagus and finishing with the intact worm.
(2) Recently, great advances have been made in our knowledge of the part played by humoral agents in the nervous integration of vertebrates. Comparative data on animals of other phyla are, however, very few. As regards the annelids, Gaskell (1919), reviewing his own work and that of earlier investigators, brought forward evidence to show that adrenaline exists in leeches and probably in certain other annelid worms, and studied the action of the drug on the body wall and blood vessels of the medicinal leech. Since that date, very little has been done on the problem of adrenaline in annelids. More recently, Bacq (1935) showed that choline esters and cholinesterase are present in polychaete worms; but, except for the well-known case of the eserinized dorsum of the leech, which contracts in response to minute amounts of acetylcholine (Fuhner, 1918) or of other choline esters (Chang & Gaddum, 1933), nothing is known about the action of this group of substances on the tissues of annelids, or about their possible physiological significance in that phylum. I therefore thought it worth while to study the action of adrenaline and acetylcholine on the different preparations employed during this research. It was originally intended to publish the experiments with drugs in a separate paper; but the results, especially with adrenaline, throw considerable light on the interrelationships of the various preparations, and it is therefore best to incorporate them with the main portion of the work.
II. THE ACTIVITY OF THE ISOLATED OESOPHAGUS
two by a transverse cut about half-way along its length, and the oral and aboral halves were separately studied. They gave somewhat different results.
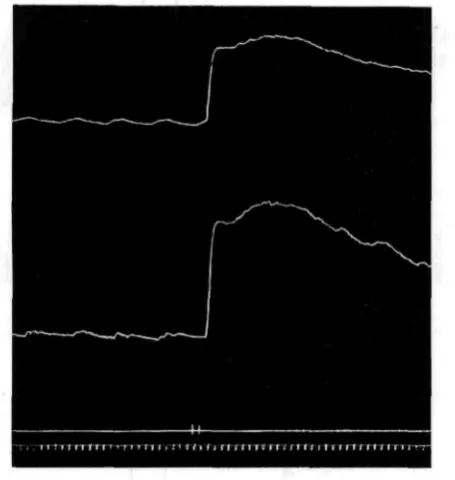
Fig. 1. The two halves of an isolated oesophagus. Oral half below; aboral above. At signal, adrenaline is added (i : 250,000).
In all records: Read from left to right; upstroke of lever records contraction of preparation; time signal
marks once every minute.
Fig. 2. The two halves of an isolated oesophagus. Oral half below; aboral above. At first signal, eserine is added (1 : 500,000); at second, about a minute later, acetylcholine (1 : 500,000).
The pieces of oesophagus thus dissected were tied at both ends and suspended in, sea water. Light isotonic levers were used in all the experiments to be described in this paper.
[image:3.451.112.340.287.527.2]ito G. P. WELLS
[image:4.451.108.363.140.437.2]The oesophagus preparations were mounted in finger bowls, to the bottoms of which disks of cork had been glued. A loop of thread tied to one end of each preparation was passed round a bent pin stuck into the cork. The action of adrenaline and acetylcholine was investigated by spraying known volumes of i : iooo solutions of the drugs (made up as described on p. 127) into the finger bowls with a hypo-dermic syringe, and subsequently computing the resulting concentrations.
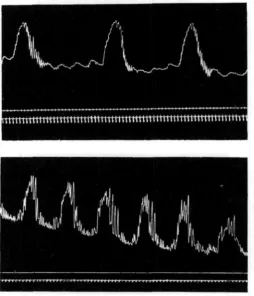
Fig. 3. Above: oral half of an isolated oesophagus, showing secondary rhythm superposed on the primary rhythm. Below, oral half of an isolated oesophagus, which gave - an unique behaviour pattern.
The behaviour of the aboral half of the oesophagus is straightforward. It shows, more or less distinctly, smooth contraction waves of low frequency, occurring in most cases about once every 5 or 6 min. (Figs, i, 2). Occasionally the interval may be as short as 3, or as long as 8 or 9 min., in exceptional preparations. In what follows, this rhythm will be termed the primary rhythm.
Both adrenaline and acetylcholine cause contracture. In the case of the latter drug, the primary rhythm can usually be seen to continue at the new level; but with the former the contracture is generally smooth, the rhythmic waves disappearing.
during a primary wave under the conditions of these experiments is of the order of one-seventieth of its total length.
The oral half of the oesophagus also shows the primary rhythm and it reacts in the same way to adrenaline and acetylcholine. The following complicating pheno-mena may however appear:
(1) The primary waves may not be smooth, but accompanied by a series of relatively rapid contractions of small amplitude.
(2) In about one-third of the preparations studied, a secondary rhythm was superposed on the primary rhythm of the oral end. This secondary rhythm consists of slow tonic contractions, of much greater amplitude than the primary rhythm and occurring at much longer intervals; they are accompanied by outbursts of relatively rapid and vigorous contraction (Fig. 3, upper half).
(3) In one only, of twenty-seven gullets studied, the oral half gave the very curious record of Fig. 3 (lower half). This is possibly an accelerated secondary rhythm.
These various complications, shown only by the oral end, are of interest as indicating a physiological differentiation between the two halves of the oesophagus. The main result of these experiments is, however, the demonstration of the slow primary rhythm, occurring in both halves of the oesophagus and therefore probably a generally diffused property of the oesophageal wall.
111. THE INVASION OF THE PROBOSCIS BY OESOPHAGEAL RHYTHMS The excised proboscis, with a short length of oesophagus attached, yields a vigorous muscle preparation, with a highly characteristic intermittent rhythm. The structure of the preparation is rather complicated, and it is necessary, before con-sidering its physiology, to understand the anatomical relations of its component parts.
(a) THE NOMENCLATURE OF THE PARTS OF THE EXTROVERT
The anatomy of Arenicola has. been described by Gamble & Ash worth (1898, 1900) and by Ashworth (1904). These accounts differ slightly in terminology; e.g. the term "buccal mass" is used in different senses. To avoid ambiguity, the sense in which the various terms will be used in this paper must therefore be specified. The terminology of Ashworth (1904) is followed, with the introduction of a new name.
122 G. P. W E L L S
shown in Plate Ib, which is a photograph of the extrovert split up along one side and pinned out with the lining upwards.
The parts of the extrovert are, from before backwards in the retracted condition: 1. The buccaltnass, characterized by the large, conical, anteriorly directed papillae. 2. The pharynx, with very numerous, fine papillae.
3. A narrow region in which the papillae are larger and more rounded than on the pharynx, which has not yet been named, and for which the name post-pharyngeal
ring is now proposed.
This ring marks the aboral limit of the part of the gut which is turned inside-out during extrusion. It is also an important physiological boundary, as we shall see later.
4. The oesophagus, whose lining is very smooth in front of the first diaphragm. In what follows, the term mouth will be used to denote the boundary between the buccal mass and body wall. The buccal mass and pharynx together constitute the proboscis.
The relations of the following structures must also be noted: the retractor
muscle, which forms a complete sheath round the aboral part of the extrovert, and is
inserted into the pharynx and into the body wall at the level of the first chaetigerous annulus; and the first diaphragm, having roughly the form of a flat cone perforated at its apex by the oesophagus,1 and bearing a pair of backwardly directed diverticula, the diaphragmatic pouches, immediately ventral to the oesophagus.
(b) THE ISOLATED EXTROVERT PREPARATION
The preparation is made as follows:
The front half of an Aremcola is pinned out, ventral surface uppermost. The body wall is divided by a cut along the mid-ventral line. This cut should just divide the first chaetigerous annulus, but go no further forwards. The flaps of body wall are pinned out sideways, that part which lies in front of the first chaetigerous annulus being turned forwards. The oesophagus is now turned forwards and the various membranes which suspend it are divided, up to the first diaphragm. At this stage the retractor muscle lies as a thick red sheath round the front part of the gut, concealing the proboscis; it must be divided right round, closfe to its origin from the body wall. When this is done properly, the buccal mass can be extended by gently pulling the oesophagus backwards, and the circum-oral nerve ring and the otocysts can be plainly seen. The gut is now ligatured twice, one thread being tied at about the level of the nerve ring and the other just aboral to the diaphragmatic pouches, and cut free.
The resulting preparation should be left for at least half an hour in sea water to recover from the dissection. It may conveniently be suspended in a bath of the pattern shown in Fig. 4, which allows the bathing fluid to be changed while the record is being taken.
1
The anatomy of the preparation is somewhat complex. It includes the buccal mass, the pharynx, the post-pharyngeal ring arid that part of the oesophagus which lies oral to the first diaphragm. It may also include the nerve ring and, owing to the way in which the animal commonly humps its mouth up during the operation, a certain amount of body wall from the region round the mouth. Moreover, as the retractor muscle is most conveniently divided along its origin from the body wall, and is not thereby separated from the first diaphragm, these two structures generally form a more or less complete sheath round the aboral half of the preparation.
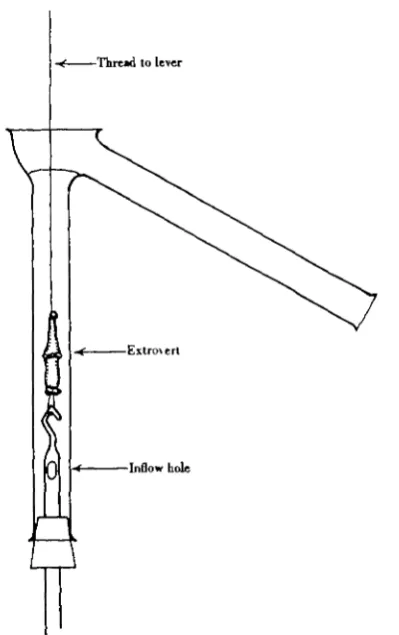
[image:7.451.130.330.188.506.2]—Thread to lever
Fig. 4. Bath used for experiments on the isolated extrovert, when the effect of changing the bathing fluid was to be studied. The new fluid flows in through a hole blown in the side of the glass hook which suspends the preparation, and sweeps the old over the side arm.
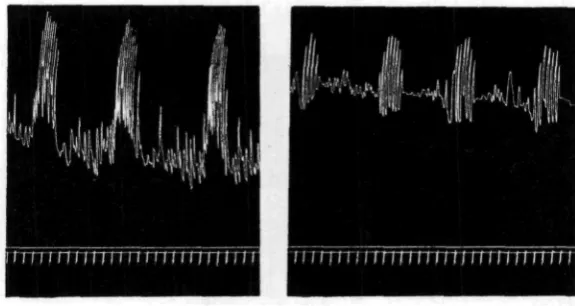
(c) BEHAVIOUR OF THE ISOLATED EXTROVERT IN SEA WATER
124 G. P. W E L L S
quiescent intervals. The four records of Fig. 5 are chosen to illustrate the range of variation as regards the latter factor that may be met with.
The individual strokes of the lever, during the rhythmic outbursts, are not simple contractions and relaxations of the longitudinal musculature, but are some-what more complex acts. If a vigorous preparation is watched, two complicating factors are usually evident: firstly, the preparation does not contract simultaneously along its whole length, the oral end contracting first, and secondly, the relaxation of
[image:8.451.43.412.145.458.2]••nn r t r r r r m r t i i i i t u i n i r
Fig. 5. Records of four isolated extrovert preparations, to show the range of variation encountered.
the longitudinal muscles (downstroke of the lever) is apparently accompanied by contraction of the circular fibres. In the present communication, the nature of the individual strokes will not be analysed in detail. Attention will be directed to the general activity pattern, with its characteristic alternation of active and resting phases.
(d) BEHAVIOUR OF THE ISOLATED EXTROVERT IN BODY FLUID
intermittent rhythm of the Arenicola extrovert. The point is illustrated by Fig. 6, which was obtained by the writer during an earlier investigation. It shows the behaviour of an atypical lobster heart, which, unlike most lobster hearts, gave grouped beats whenever it was perfused with a potassium-free solution. Except for the great difference of time scale, the picture is extraordinarily like that given by many of the lugworm preparations. Similar grouped beats have been recorded by Hogben (1925) in crustacean hearts in the presence of excess potassium, by Mines (1912) in the dogfish heart after arrest by lack of urea or by magnesium excess, and also by other authors.
[image:9.451.164.287.251.320.2]The suspicion therefore arises that the intermittent activity of the Arenicola extrovert is an artefact, and that sea water is an unsuitable chemical environment for the tissues of this animal. This point is of great importance, because sea water was used as bathing medium throughout this investigation. I have therefore made experiments in which the preparation was suspended in body fluid from Arenicola, to compare its action with that of sea water.
Fig. 6. Grouped beats of an atypical Homarus heart, in a potassium-free perfusion fluid. The time scale is different from that of the Arenicola records, the duration of the extract here shown being about 2 min. Compare Fig. 5.
It may first be pointed out that where the blood vessels of Arenicola penetrate among the other tissues, they always run in tubular prolongations of the coelom, so that it is the coelomic fluid, and not the blood, which forms the immediate chemical environment of the cells.
The experiments were done in July, when germ cells were thickly suspended in the body fluid. About sixty freshly collected lugworms were "bled" by opening the body cavity, and as each worm was opened the fluid was allowed to flow through muslin on to filter paper in a Buchner funnel. In this way it was freed as rapidly as possible from most of the suspended cells, to minimize the possibility of contami-nation by products of cellular breakdown. Such contamicontami-nation might, for instance, raise the potassium concentration significantly. The time which elapsed between the shedding of the fluid and its passage through the funnel was always less than 10 min. The pooled fluid, thus collected, was still slightly cloudy; it was therefore filtered a second time. This was done at about 6 o'clock, the "blood-letting" having begun about midday.
126 G. P. W E L L S
very unlikely that significant differences in electrolyte concentration exist between the two. The fluid was used about 4 hours after the final filtration,
Preparations suspended in body fluid showed an activity pattern essentially like that already described (Fig. 7). The following differences were noticed: first, the number of lever strokes in each rhythmic outburst is less in body fluid than in sea water and the duration of each individual contraction is longer, and second, on changing from sea water to body fluid there is a sharp rise of tone, which is reversed when the opposite change is made.
[image:10.451.104.392.227.380.2]This reversible tone rise is probably not due to a difference in electrolyte centrations between the body fluid and sea water. The effects of varying the con-centrations of K, Ca and Mg in the medium have been studied in some detail by the writer, and will be described in a separate paper. From the results of those experi-ments, it is likely that the composition of the body fluid, as regards inorganic salts,
Fig. 7. Two records of the same isolated extrovert preparation. On the left, in sea water. On the right, 30 min. after changing to Arenicola body fluid.
is very similar to that of sea water. Certain changes (e.g. Mg lack, K excess) produce reversible tone rises like that evoked by body fluid, but in these cases the rhythmic pattern is conspicuously modified. The same is true of adrenaline (see next section), which produces a tone rise but profoundly alters the behaviour pattern. Of the various substances studied by the writer, only acetylcholine resembles body fluid in producing a tone rise without disturbance of the characteristic intermittent rhythm. It would be unsafe to conclude on this basis alone that the body fluid contains acetylcholine, for the only chemical agents so far studied are adrenaline, eserine, acetylcholine and salts. It does, however, seem clear that none of these agents, except only acetylcholine, could be responsible for the differences observed between sea water and body fluid.
same physiological mechanism, and that the Arenicola extrovert is normally in a condition which appears as an occasional freak in other rhythmic muscles.
(e) ADRENALINE, ACETYLCHOLINE AND THE ISOLATED EXTROVERT
A large number of experirhents were done to determine the action of adrenaline and acetylcholine on the isolated extrovert. Because of its vigour and its highly specific behaviour pattern, the preparation is well suited for investigations of this kind. It is best to consider the results before passing to the next section, in which the site of origin of the excitations is localized.
Methods
The experiments were done in the bath illustrated in Fig. 4. The solutions were made up as follows.
Acetylcholine. A M/$ solution of sodium phosphate was acidified with HC1 until
just yellow-green to brom cresol green (pH 4-2). This gave a solution containing Af/5 acid sodium phosphate and Af/5 NaCl, having a pH at which acetylcholine is very stable. "Roche" acetylcholine was made up 1 : 1000 in this acid phosphate, and used, in most cases, on the same day.
Eserine. This drug was made up 1 : 1000 in the acid phosphate diluted tenfold
with distilled water. It was used in most cases within 48 hours of making up, being kept overnight in an ice chest, and showed no traces of discoloration.
Adrenaline. In most of the experiments on this substance, Parke Davis 1 : 1000
adrenaline solution was used. As this solution contains five times as much chlore-tone as adrenaline, control experiments were carried out with chlorechlore-tone in appro-priate concentrations, and also with(a dry preparation of adrenaline ("Adrenalina B.P." from British Drug Houses Ltd.), to make sure that the adrenaline was in fact responsible for the effects observed.
During an experiment, the 1 : 1000 solutions were kept in burettes on the bench, and were diluted in sea water (pH 8-2) to the required concentration immediately before application to the tissue. To guard against any possibility that the acetyl-choline was being inactivated by hydrolysis in the alkaline sea water, the experiments with that drug were checked over, using sea water acidified with HC1 to pH 67 as bathing fluid. The results were the same as with pH 8-2.
Action of adrenaline
128 G. P. WELLS
With low concentrations of adrenaline, the picture is not always so clear. A tone rise can be detected in concentrations down to i : 100,000,000. In most preparations a definite inhibitory action on
the normal behaviour pattern, appearing as a lengthening of the interval between the rhythmic outbursts, is exerted by adrenaline 1 :50,000,000 or 1 : 100,000,000 (Fig. 9). With concentrations of 1 :500,000,000 or lower there is usually no effect, but some-times a very slight and ephemeral tone rise, accompanied by slowing of the rhythmic outbursts, can be made out, even in adrena-line 1 : 2,000,000,000.
To summarize, the principal adrenaline actions are: (1) a rise of tone, (2) in low concentrations, a lengthening of the inter-val between the normal rhythmic outbursts,
and (3) in high concentrations, the evocation of a regular, continuous rhythm. The first and third effects are the most striking: and in investigating the action of
Fig. 8. Isolated extrovert preparation. Adrenaline i : i,000,000 at signal.
Fig. 9. Isolated extrovert preparation. Adrenaline 1 : 50,000,000 at signal.
adrenaline on the other preparations employed during this work I have only used those concentrations which evoke continuous rhythm in the isolated extrovert.
Action of acetylcholine and eserine
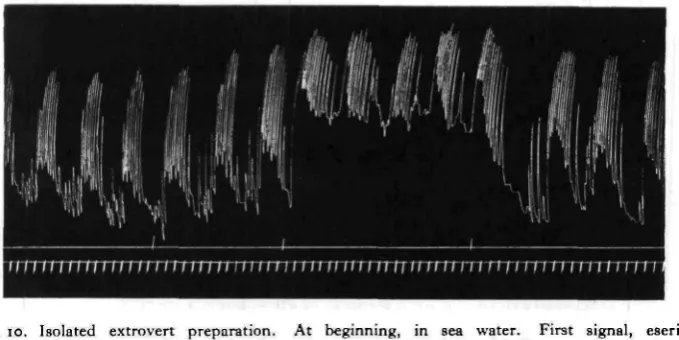
generally calls forth a very slight, gradual rise in tone, and, especially in a weak or irregular preparation, it may increase the vigour of the beats during the active phases; but the essential activity pattern is unaltered (Fig. 10).
Fig. io. Isolated extrovert preparation. At beginning, in sea water. First signal, eserine 1 : 500,000 in sea water. Second signal, eserine i : 500,000 and acetylcholine 1 : 1,000,000 in sea water. Third signal, sea water only.
Fig. 11. Isolated extrovert preparation. At beginning, in eserine 1 : 500,000 in sea water. First signal, eserine 1 : 500,000 and acetylcholine 1 : 1,000,000 in sea water. Second signal, eserine 1 : 500,000 and acetylcholine 1 : 100,000 in sea water.
[image:13.451.106.355.313.475.2]130 G. P . W E L L S
The extrovert is not particularly sensitive to acetylcholine. After exposure to eserine i : 500,000 for 2 or 3 hours, a clear tone rise is produced by acetylcholine
1 : 10,000,000 but not 1 : 100,000,000.
(/) SITE OF ORIGIN OF THE ACTIVITY OF THE ISOLATED EXTROVERT
In a series of experiments, the living extrovert was dissected in various ways in order to localize the site of origin of the excitations. The anatomy of the stomato-gastric system has not yet been worked on* in Arenicola, so one has no anatomical guide from which to work.
Restriction of the field of enquiry to the gut wall
As already pointed out, the anatomy of the preparation is complex, and, as dissected out for routine purposes, it includes the following two groups of structures in addition to the gut wall:
(1) The circunvoral nerve ring and a certain amount of body wall from the mouth region are commonly included.
(2) The aboral half of the preparation is enveloped by a more or less complete sheath consisting of the retractor muscle and most of the first diaphragm, with the diaphragmatic pouches.
The following experiments deal respectively with these inclusions:
(1) A series of preparations was made in which the oral cut and ligature were made behind the nerve ring. To be sure of this, the nerve ring, otocysts and pro-stomium were carefully exposed in each worm after the extrovert had been removed. The extroverts showed the typical behaviour pattern, and responded in the usual way to adrenaline.
(2) In four preparations the outer sheath was dissected off under a binocular microscope. Difficulty was only encountered in the median plane, where the gut wall is closely bound to the first diaphragm. However, the experiments on radial localization in the next section show that the region of this median partition cannot be the exclusive site of origin of the characteristic activity pattern. Over most of the surface of the gut, the outer sheath was satisfactorily removed. The resulting preparations gave the typical intermittent rhythmicity, and the typical response to adrenaline.
Clearly then, the field of enquiry is restricted to the following parts of the gut wall: the buccal mass, the pharynx, the post-pharyngeal ring, and that part of the oesophagus which lies in front of the first diaphragm.
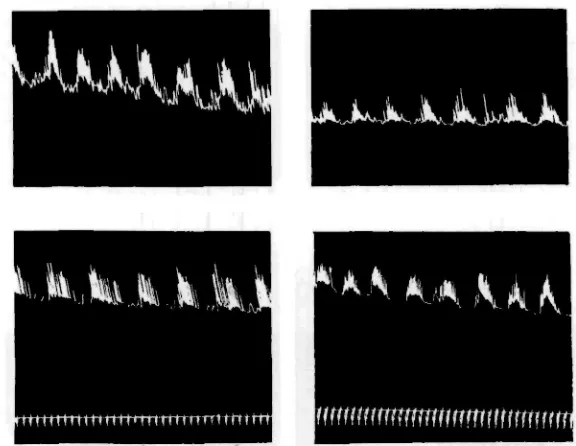
Radial localization
characteristic outbursts of rhythmic activity, although in this case, owing to the small size of the strips, the contractions are not as powerful as before (Fig. 12).
[image:15.451.57.345.121.344.2]Evidently we are not dealing with a special ganglion, or other strictly localized centre. The structure from which the excitations emanate is either a ring running round the gut or something diffuse, like a nerve plexus.
Fig. 12. Isolated extrovert divided into four longitudinal strips of approximately equal thickness. All show the typical behaviour pattern.
Longitudinal localization
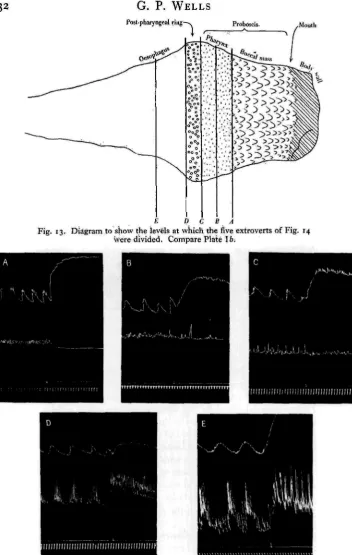
The experiments on longitudinal localization were done as follows. The extro-vert was split longitudinally and pinned out, inner side uppermost, on a disk of cork stuck to the bottom of a finger bowl. If such a preparation is examined under sea water with a low-powered binocular, the different regions of the gut can be readily distinguished (Plate Ib). The extrovert is now divided by a single transverse cut into two halves. The cut edges are pinned to the cork, the rest is freed, and the two ends are connected by threads to light isotonic levers. The level of the transverse cut was varied in the different experiments, the various levels employed being illustrated in Fig. 13. The effect of cutting at each level was investigated in at least three preparations, and in m6st cases in six.
The specifications of the various cuts are as follows:
A. Separates the buccal mass from the pharynx. B. Bisects the pharynx.
C. Runs across the pharynx, immediately oral to the post-pharyngeal ring.
G. P. WELLS
PosJ-pharyngeal r i n g - . Proboscis.
° 0
• • » •
o o
oo ' ' ,*
-' -' , -'
• ' : ' • ' '
• ' ' '
[image:16.451.71.424.34.590.2]E D C B A
Fig. 13. Diagram to show the levels at which the five extroverts of Fig. 14 were divided. Compare Plate 16;
E. Runs across the oesophagus, at a distance from the post-pharyngeal ring
equal to about twice the width of the ring.
F
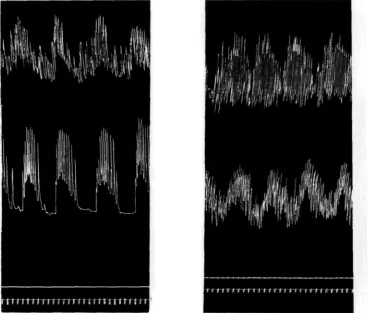
K^' Vk rTW°- e l r t r o v e r t 8' divided respectively along the lines C and E of Fig. 13. In each case the aDoralnalt writes above the oral. At first signal, eserine 1 : 500,000; at second, acetylcholine 1:250,000.
The results are shown in Figs. 14 and 15, the main points being these: (1) In every case the aboral half of the preparation shows tone waves occurring at intervals of several minutes, and clearly identical with the primary rhythm that
134 G. P. WELLS
we have already detected further back in the oesophagus (p. 120). In many cases these waves are accompanied by outbursts of rhythmic strokes of small amplitude, the behaviour being then essentially like that of the entire extrovert. Moreover, the aboral end of the preparation is invariably excited by adrenaline. Acetylcholine produces a sharp rise of tone, the waves continuing at the new level. Here again, the aboral half resembles the entire extrovert.
(2) In the case of cuts D and E, the oral half contains a certain amount of oeso-phageal tissue. It shows the typical behaviour pattern, and reactions like those of the whole extrovert to adrenaline and acetylcholine.
(3) In the case of cuts A, B and C, the oral half contains no oesophageal tissue. Its behaviour is now quite unlike that of the intact extrovert. Instead of rhythmic outbursts it shows a continuous, more or less regular activity, and, in striking
Fig. 16. Preparation for investigating the relation between the movements of the oesophagus and proboscis (compare Plate 16).
contrast to the whole extrovert, it is inhibited by high concentrations of adrenaline. With acetylcholine, there is no immediate response. Sometimes, but not always, an acceleration of the movements develops after several minutes, as in the record printed in Fig. 15.
(4) Preparations cut half-way between C and D, bisecting the post-pharyngeal ring, gave contradictory results. In some the oral end showed signs of the typical activity pattern; in others it behaved as if the cut were made at A, B or C.
Finally, if the proboscis is in continuity with oesophageal tissue, the oesophageal rhythm invades the proboscis, and we thus get the behaviour pattern of the entire extrovert.
Any remaining doubt that the last statement represents the true course of events is removed by the experiment illustrated in Figs. 16 and 17. An extrovert is split along one side and pinned out on cork. About half of the oesophagus is included in this experiment. Six pins are used, inserted at the points shown in Fig. 16. To the two ends of the preparation, threads are attached so that their movements can be separately recorded, though they are still continuous with each other. The pins
[image:19.451.106.350.188.409.2]irrtrrrnitttimttmmmiuTitumutimttti
Fig. 17. Record of an extrovert pinned out as in Fig. 16. The oesophagus writes above; the proboscis below. At signal, the extrovert was divided along the junction of the pharynx with the post-pharyngeal ring.
136 G. P. W E L L S
This makes it plain that the rhythmic outbursts of the isolated extrovert prepara-tion are due to the waves of the primary rhythm of the oesophagus, flowing forwards and reaching the more vigorous musculature of the proboscis.
A word of caution may here be added as to the intimate nature of the oesophageal contraction waves. The term "tone waves" has been used from time to time in describing the primary rhythm of the oesophagus because, in many cases, and especially with the aboral half of the oesophagus, the waves look perfectly smooth on my records. It is, however, not intended to imply that the oesophagus shows smooth, steady waves of excitation which are resolved, on reaching the proboscis, into groups of separate contractions. In many of the records of the oral half of the oesophagus the "tone waves" have numerous minute contractions superposed upon them; this can be seen for instance in the upper line of Fig. 17, where the appearance strongly suggests that each of the single strokes of the proboscis during the rhythmic outbursts corresponds to a minute contraction of the oesophagus. Unhappily, as already pointed out, the oesophageal contractions are of very small amplitude and my levers were not sufficiently sensitive to enable a definite decision to be reached on this point. That the activity outbursts of the proboscis emanate from the oeso-phagus is certain; whether the same is true of the single contractions during the outbursts is not yet clear.
(g) SITE OF ACTION OF ADRENALINE AND ACETYLCHOLINE
The preparation of Fig. 16 can be used to analyse further the action of drugs on the extrovert. If such a preparation be adrenalinized (Fig. 18), the oesophageal end only goes into contracture, while the proboscis exhibits a regular rhythm instead of the previous rhythmic outbursts. If now the preparation is severed along the post-pharyngeal ring, the movements of the proboscis cease.
In the particular experiment of Fig. 18, the oesophagus shows small tone waves during exposure to adrenaline, and at first glance the proboscis seems to be following the waves as it did before. This is, however, not the case. The undulation of the proboscis trace is now due to variation in amplitude, the frequency being pretty constant throughout; and the relation between the waves is now reciprocal, the amplitude of the contractions of the proboscis diminishing every time the tone of the oesophagus rises. The drug has profoundly altered the behaviour of the system. Clearly, then, the response of the entire extrovert to adrenaline, as seen for instance in Fig. 8, is complex and involves the following factors: (1) contracture of the oesophageal tissue only, and (2) rhythmical contractions of the musculature of the proboscis, which are due to excitations emanating from the oesophagus.
in nature from the primary rhythm, and is superposed upon it without inter-rupting it.
Fig. 18. Extrovert pinned out as in Fig. 16. Oesophagus above; proboscis below. First signal, adrenaline i : 250,000. Second signal, preparation severed.
Fig. 19. Extrovert pinned out as in Fig. 16. Oesophagus above; proboscis below. First signal, eserine 1 : 500,000. Second signal, acetylcholine 1 : 250,000.
i38 G. P. WELLS
acetylcholine, the oesophagus goes abruptly into contracture; the proboscis does not, but shows a slow, steady tone rise (compare Fig. 15 C), continuing meanwhile to follow the primary waves which the oesophagus exhibits at the new level.
It is interesting to note that the primary waves in the oesophagus resemble the contracture induced in the same organ by high concentrations of adrenaline, in that they are accompanied by rhythmic activity of the proboscis, whereas the secondary waves resemble acetylcholine contracture in having no effect which is transmitted to the proboscis, and in failing to interrupt the primary rhythm.
IV. THE INVASION OF THE CENTRAL NERVOUS SYSTEM AND BODY WALL BY OESOPHAGEAL RHYTHMS
The excitations of oesophageal origin which spread to the proboscis do not end there. By means of the preparation about to be described, their influence can be traced in the movements of the body wall of the worm.
(a) THE EXTROVERT : BODY-WALL PREPARATION
A lugworm is pinned out ventral surface upwards, and opened by a longitudinal incision running about 3 mm. to one side of the fine, mid-ventral groove that gives an outwardly visible indication of the
position of the nerve cord. This cut should just divide the first chaetigerous annulus, but go no further forwards. The flaps of body wall are then pinned out, the oeso-phagus is freed, and the retractor muscle is divided, as already described on p. 122. During the latter operation care is necessary at the point where the nerve cord perforates the retractor muscle; the cord must of course not be damaged. By means of two transverse cuts, a strip of body wall from the region of the second chaetigerous annulus is isolated from the rest, retaining, however, its con-nexion with the nerve cord (Fig. 20). The whole preparation is securely pinned to cork stuck to the bottom of a finger bowl, and threads are tied to the oesophagus, just aboral to the diaphragmatic pouches, and to the tip of the body-wall strip, for con-nexion to very light isotonic levers.
After the dissection, the preparation should be left for ^-i hour in sea water,
preferably aerated, before the experiment begins. Again, after connecting the threads to the recording levers, it may take another 20 min. or so to "get into its stride", as it is very sensitive to mechanical disturbance.
Oeiophagui
Probotcu
Nerve cord
(b) BEHAVIOUR OF THE PREPARATION IN SEA WATER
[image:23.451.223.379.128.472.2] [image:23.451.39.369.136.474.2]In a good preparation1, the extrovert shows outbursts of activity as described in the last part. The body-wall strip shows associated outbursts of rhythmic con* traction, occurring at the same time as the outbursts of the extrovert (Fig. 21). In between these outbursts, the body wall relapses into a slower rhythm of its own.
Fig. 21.
Fig. 21. Extrovert : body-wall preparation. Extro-vert above; body wall below. At X, the nerve cord is cut between the two strips.
Fig. 22. Extrovert : body-wall preparation, with longitudinal strip added. Circular muscle strip of body wall above; extrovert in the middle; longi-tudinal muscle strip of body wall below.
Fig. 22.
That the excitations responsible for the outbursts do in fact flow from the extrovert to the body wall is demonstrated by cutting the nerve cord between the two strips. After a sharp contraction of both strips, due to the stimulus of the cut, the activity of the extrovert continues as before, but the outbursts are no longer transmitted to the body wall behind the cut. The latter now steadily maintains its intrinsic rhythm.
1
[image:23.451.33.201.138.395.2]140 G. P. W E L L S
In a good preparation, the body wall follows the extrovert stroke for stroke during the rhythmic outbursts. The contraction of the two strips is, however, not synchronous, but reciprocal. One lever rises as the other falls. This fact suggests that we have to do with a reciprocal activity of the longitudinal and circular muscu-lature. The upstroke of the upper lever is due to contraction of the longitudinal muscles of the extrovert, and the appearance of a vigorous preparation strongly suggests, as already pointed out in an earlier section, that the downstroke of the lever involves, not only relaxation of the longitudinal muscles, but also contraction of the circular fibres. It is at this moment that the body-wall strip, which is of course a circular muscle strip, contracts. The individual strokes seem therefore to incor-porate two acts: first a contraction of the longitudinal muscles of the extrovert, and possibly also of the body wall of the first one or two segments, then a relaxation of the longitudinal muscles and a contraction of the circular fibres in both situations.
Evidence is adduced in a later section to show that the outbursts of activity in the dissected preparations correspond to outbursts of extrusion and retraction of the proboscis in the intact animal. As Just (1925) has pointed out, the acts of extrusion and retraction involve respectively contraction of the circular and longitudinal muscles of the body wall of the anterior end. In the preparation with which we are here concerned, then, the upstroke of the lower lever is an attempted extrusion, and the upstroke of the upper lever is an attempted retraction.
The analysis of the individual strokes has not yet been carried any further, Mention may, however, be made of a single experiment, which I have had no opportunity of repeating, but which gives a further indication of the great complexity of the neuromuscular mechanisms with which we are dealing.
A preparation was made as already described (Fig. 20) but, in addition, a longi-tudinal strip of body wall was dissected, which included the nerve cord and ran back through three or four body segments, starting behind the circular muscle strip. Thus three traces were simultaneously obtained. The extrovert and circular muscle strips were co-ordinated in the usual way. From the foregoing considera-tions I expected to see the longitudinal strip show correlated movements, syn-chronous with those of the extrovert. But what actually happened was this: during the rhythmic outbursts it progressively relaxed, and at the end of each outburst it contracted quite rapidly (Fig. 22).
Fig. 23. Extrovert : body-wall preparation. Extrovert above; body-wall strip below. At first signal, nerve cord divided; at second, adrenaline applied (1 : 250,000).
[image:25.451.50.362.272.586.2]143 G. P. WELLS
(c) THE ACTION OF ADRENALINE AND ACETYLCHOLINE
The action of these drugs was studied by adding known volumes of i : iooo solutions (p. 127) to the finger bowl containing the preparation, and subsequently computing the resulting concentrations.
If adrenaline is applied to a preparation whose nerve cord has already been cut between the extrovert and the body-wall strip, the following results are seen: the extrovert shows the usual response, its tone level rising sharply and its rhythm becoming continuous. The tone and movements of the body wall, on the other hand,
Fig. 25. Two extrovert : body-wall preparations. Extrovert above; body wall below. Left: prepara-tion whose nerve cord has been divided between the extrovert and the body-wall strip. At signal, eserine 1 : 500,000 and acetylcholine 1 : 125,000 are simultaneously applied. Right: preparation whose nerve cord is intact. At signal, eserine 1 : 500,000 and acetylcholine 1 : 250,000 are simultaneously applied.
are isolated from oesophageal influences, they both show a continuous rhythm which is inhibited by adrenaline. If, on the other hand, the normal conduction paths are intact, they are both invaded, not only by the primary rhythm of the oesophagus, but also by the continuous excitation induced in that organ by adrenaline. Physio-logically, then, the proboscis is stomodaeum. Whether the post-pharyngeal ring corresponds to the hinder limit of the morphological stomodaeum is, unhappily, not known.
Acetylcholine, applied to a preparation after the nerve cord has been severed, induces its usual effect in the extrovert—a sharp tone rise, the rhythmic outbursts continuing at the new level. No evident effect is produced in the body wall, except perhaps for a very slow, very slight rise of the base line (Fig. 25, left half). Appli-cation of acetylcholine to the preparation before section of the nerve cord has still no effect on the body muscles. The sharp tone rise of the extrovert is not transmitted to the body wall, which continues to follow the rhythmic outbursts of the extrovert as it did before (Fig. 25, right hah0). Once again, on comparing Fig. 25 with Figs. 15 and 19, a striking similarity between proboscis and body wall is seen.
V. OESOPHAGEAL RHYTHMS IN THE INTACT ANIMAL
An attempt has been made to find out what part the facts described in the preceding sections play in the physiology of the intact worm. We have seen in the dissected preparations that the oesophagus has a slow intrinsic primary rhythm, that every wave of this rhythm involves the discharge of a series of excitations which pass forwards, via .the proboscis, into the central nervous system, and that the musculature of the anterior end is thereby thrown, at regular intervals, into out-bursts of vigorous rhythmical contraction. During these outout-bursts there seems to be a well-marked reciprocal action of the longitudinal and circular muscles. Unless the whole phenomenon is artificial and due in some way to the experimental methods employed—a supposition that seems hard to believe in view of the facts (a) that the isolated extrovert gives essentially the same behaviour pattern in Arenicola body fluid as it does in sea water, and (b) that the isolated oesophagus, the isolated extro-vert and the extroextro-vert: body-wall preparation form an orderly series and show the same basal pattern with various degrees of elaboration—then it should be possible, by carefully watching the intact worm, to detect outbursts of some kind of muscular activity, corresponding to the outbursts of the various dissected preparations.
144 G. P. W E L L S
of minimal experimental interference. I have therefore made a series of observa-tions of this type in order to find out whether a periodicity of eating or burrowing movements could be detected.
(a) METHODS
In designing these experiments a compromise had to be struck between com-plete visibility of the worm and normality of conditions, i.e. the exclusion of factors, such as the absence of adequate contact stimuli, which might perhaps disturb the oesophageal rhythm. A preliminary attempt to watch the behaviour of worms allowed to burrow in a thin layer of sand between two vertical glass plates was unsuccessful. Even if the plates were so close together as to cause a considerable degree of compression of the animal, the sand surrounded the head and made close observation impossible. The method finally adopted was as follows. The worm was placed in a glass U-tube of internal diameter i cm., about 45 cm. in height and 17 cm. from limb to limb. Near the upper end of the U-tube a narrow cross-tube connected the two limbs; thus a gentle circulation of the sea water in the tube could be maintained by a capillary aeration jet inserted into one of them. The whole was immersed in a glass water bath to reduce temperature fluctuations. Under these conditions the worm rapidly settled down in the tube, lying usually head downwards near the lower end of one of the vertical limbs, and carrying out respiratory move-ments of the usual type. The observations were made after it had been left in the tube for at least an hour—overnight, in most cases—so that it could accustom itself to its situation. Altogether, twenty-eight worms were studied, each one being watched continuously for a period of from £ to 1 hour.
In making the observations, no account was taken of such movements as the respiratory waves, waving of the tail or opening and closing of the anus. Only the movements of the head end were noted in detail.
The following three types of anterior end activity were seen:
1. The head may be perfectly still, in which case it usually rests against one side (not always the lowest) of the glass tube, and often with the mouth curved down to touch the glass.
2. The proboscis may be rhythmically extruded and withdrawn, the associated movements of the body wall being more or less vigorous and the extrusions more or less complete.
In most of the experiments the behaviour of the worm was recorded simul-taneously in two ways. The movements were noted down from minute to minute on a written protocol, and were also registered by means of two morse keys, placed near the observer's left hand and connected with two signals writing on a drum. One key was tapped for each of the head movements not resulting in extrusion; this key was connected with the upper signal (Fig. 26). The other was tapped for each extrusion; this key activated both signals. This method of connecting the signals was adopted in conformity with the view already suggested that the head movements not resulting in extrusion are really only gentler forms of the extrusion act. The kymograph records should therefore be interpreted as follows: the upper trace indicates the timing of a series of muscular acts, all essentially of the same type, while the lower trace shows whether the acts were vigorous enough to result in extrusion of the proboscis.
Of the two kinds of record, the protocols are the most valuable, because, except for the distinction between extrusion and movements not resulting in extrusion, the kymograph records give no indication of the vigour of the various acts but only show their timing.
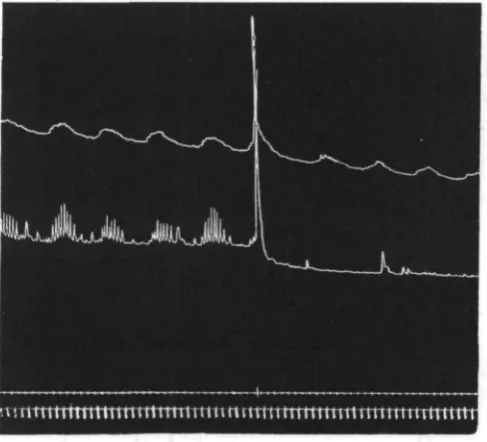
(b) THE RESTING RHYTHM
[image:29.451.78.378.330.508.2]In the great majority of records obtained, there is an evident periodicity in the activity of the anterior end. In a few cases, including the upper one of Fig. 26, there
Fig. 26. Two records obtained by observation of intact lugworms. Upper signal, movements of head. Middle signal, extrusions of proboscis. Lower signal, time (minutes).
146 G. P. WELLS
Table I
Time p.m. 5-42/43 5-43/44 5-44/45 5-45/46 5-46/47 5-47/40 548/49 5-49/50 5-5°/5' 5-51/52 5-52/53 5-53/54 5-54/55 5-55/56 5-56/57 5-57/58 5-58/59 5-59/6o 6.0/1 6.1/2 6.2/3 6.3/4 BehaviourHead moving from side to side
Five extrusions Seven extrusions One extrusion
Head swaying gently Head swaying gently Swaying gently, then quiet Very quiet
Becomes rather more restless. Head lengthens, then shortens
Very quiet, then three extrusions
Seven extrusions
One extrusion, then sways head
Head explores slightly, animal standing "on tip-toe", with anterior parapodia extended Quiet; then backs along tube, with gills waving Head sways slightly; still quiet
Head explores gently
Explores gently; then two extrusions
Nine extrusions Seven extrusions
Three extrusions, then quiet
Very quiet
Head swaying gently
Table I gives a very clear picture of the animal lying quiet, then becoming restless and giving an outburst of extrusions, presumably under the influence of an oesophageal primary wave, then becoming quiet again, and so on. In this particular case the outbursts come once every 8 min.
In other cases, such as the lower, record of Fig. 26, the periodicity is still clearly traceable, but the animals are less vigorous in their behaviour. Instead of giving outbursts of extrusion they give outbursts of swaying, of lengthening and shortening of the head, or of gulping, the proboscis being seldom or never extruded. Between the outbursts the head is quiet, usually with the mouth gently pressed against the side of the tube. Then rhythmical raising, swaying and stretching of the head begins, the movements increasing at first in vigour and extent and then decreasing again as the outburst passes off. In these cases, it is reasonable to suppose that the outbursts are still due to the primary rhythm waves of the oesophagus, the difference being that the excitations spread to fewer muscle fibres, and thus call forth less vigorous movements.
activity seems to fluctuate, but it is not possible definitely to locate the activity maxima and minima on the protocol.
The restlessness produced by the oesophageal outbursts is not always confined to the anterior end. Sometimes, for instance, as each activity wave begins, the worms suddenly creep backwards for several inches along the tube, waving their tails.
The duration of one activity cycle varies from worm to worm and even from time to time in the same worm. These variations are, however, paralleled in the records of the isolated extrovert. Different preparations vary somewhat in their outburst frequency, and they may occasionally show an abnormally long pause, or an abnormally prolonged outburst, in the middle of an otherwise regular record.
Confirmation of the view that the outbursts of anterior end restlessness are • indeed due to the primary rhythm of the oesophagus is obtained by comparing the
time relationships of the two cycles. The durations in minutes of all the cycles shown in my protocols of the intact worms (as measured from minimum to minimum) were determined and the mean and standard deviation were calculated, the result being 6-60 ± 2-13 min. From the material upon which the first section of this paper, dealing with the isolated oesophagus, is based, a similar computation was made, the result being 6-20+ 1-33 min. for oral ends and 6-22 + 0-99 for aboral ends. Con-sidering that we are dealing in the one case with a whole animal and in the other with an isolated organ in sea water, the agreement is sufficiently good.
I am inclined to the belief that the records obtained from the most vigorous animals, e.g. the upper one of Fig. 26, represent most closely the normal behaviour of the worms. As already pointed out, they live in burrows eating the surrounding mud, and my results suggest that this eating occurs in rhythmical outbursts with ah average frequency of one outburst in every 6 or 7 min. In any case, the occurrence of activity cycles of appropriate period in the great majority of the intact worms observed makes it practically certain that Arenicola has a "resting rhythm" of activity of some kind whose pace is set by the oesophagus.
(c) THE ACTION OF ADRENALINE
It was shown that the various dissected preparations can be thrown into con-tinuous rhythmical activity by adrenaline in concentrations from 1 : 1,000,000 upwards, provided that the normal conduction paths connecting the oesophagus, on which the drug acts, with the musculature of the proboscis and body wall are intact. If, therefore, it is true that the outbursts of extrusion (or swaying, etc.) described in the last section are identical with the rhythmic outbursts of the dissected pre-parations, then it should be possible to elicit continuous anterior end activity in the intact worm by injection of adrenaline.
A series of experiments was carried out to test this point, the conditions being as follows:
148 G. P. WELLS
Table II. Numbers of extrusions
-A 1 A z A 3 A4 A5 A 6 A 7 A 8 A g A 10 A 11 A 12 Total per 5 min. interval C i C 2 C3 C4 C 5 C 6 C 7 C 8 C g C 10 C I I C 12 Total per 5 min. interval 1 26 1 2 18 32 18 24 13 2 4 26 2 4 26 5° 293 1 4 3 23 — 5 —• 3 i 2g 2 0 33 149 2 15 2 0 15 29 19 2 1 1 1 2 1 23 24 2 0 34 252 1 2 7 18 3 9 — 15 26 2 1 27
1 2 9
3 1 0 17 18 2 0 16 19 1 1 2 1 19 23 2 2 30 226 — 7 16 13 18 — 24 25 2 4 29 156 4 1 0 ' 5 17 1 2 1 1 18 1 0 19 2 1 2 1 19 24 197 1 — 1 0 7 4 6 — 18 25 2 0 3°
1 2 1
5 1 5 nin. 6 intervals 7 Adrenalinized 8 1 2 16 1 0 1 2 17 2 2 15 19 16 19 2 0 186 7 1 0 1 1 13 17 14 19 17 16 14 19 26 183 Contro — — 4 1 14 — — 7 1 0 1 1 36 83 — — 8 1 4 — 1 5 — 1 9 19 57 3 5 12 1 2 16 16 2 2 1 2 17 17 18 2 1 171 8 9 animals 8 9 6 4 16 14 2 1 16 16 16 15 13 154 animals — — 5 1 2 3O 6 5 — 5 5 59 — — 2 — 1 0 — 29 1 — 15 1 58 1 8 8 9 13 13 19 13 1 1 16 16 18 H 5 — 2 — — 1 2 — 2 1 — 1 0 9 I 55 1 0 6 6 5 8 14 14 19 14 6 15 16 19 142 — — — — 4 — 16 — — 2 3 25 1 1 5 5 1 2 7 9 16 17 13 8 13 13 13 131 — — — — 3 4 — 6 2 3 18 1 2 6 5 7 6 7 12 1 2 18 15 15 1 0 " 3 — — — — 2 7 13 8 8 2 4 4 4 Total per animal i°5 124 138 163 167 193 196 197
2 0 0
2 1 4 218 2 7 8
2193 3 8 46 67 7 i 75 9 0 114 139
1 5 0 1 9 1 954
In the case of the adrenalinized animals, the injection fluid consisted of one part of Parke Davis adrenaline solution1 to ten parts of sea water. In the case of the controls, it consisted of one part of a 0-5 per cent solution of chloretone in 0-9 per cent NaCl to ten parts of sea water.
(2) After injection, the animal was put in a glass tube (length 26 cm., internal diameter 1 cm.) and immersed in a bath of sea water. Strips of mirror were so disposed that all sides of the worm could readily be seen. The tube was closed at the ends by corks perforated by narrow glass tubes, so that the animal could renew the water in the tube, to some extent at least, by its movements. It was found by Borden (1931) that the haemoglobin of Arenicola carries enough oxygen to supply the animal's needs for over an hour.
(3) When in the tube, the worm was watched continuously for 1 hour. This observation period began exactly 5 min. after the injection. This allowed ample time for putting the animal in the tube and cleaning the syringe. Whenever the animal extruded its proboscis, a Morse key was tapped; this operated a signal writing on
1
Table III. Numbers of " Quiet minutes " Ai A2 A3 A4 AS A6 A7 A8 A9
A i o A 11 A 12 Total per 5 min. interval C 1 C2
11
C6 Cy C8 C9C i o
C I I
C 1 2 Total per
5 min. interval
5 min. intervals
1 2 3 4 5 6 7 8 9 10 11 12
Adrenalinized animals
— — — — 1 — 2 2 4 1 1 — — — — — — — 1 — — 1 1 1 — — — — — — — 2 3 3 1 5 — — — — 1 — — 1 — 1 — 1
_ _ j !
I I I 2
— — — 1 2 — 4 6 8 8 5 1 0
Control animals
5 5 5 5 5 5 5. 5 5 5 5 5
4 4 5 4 5 5 5 5 5 5 5 5
2 3 5 5 5 5 5 5 3 5 5 5
3 — — — 2 — 1 3 5 5 5 5
— — — 1 4 4 4 5 5 5 5 5
5 3 1 3 — 3 3 2 1 3 2 3
— 1 1 3 5 5 — 5 5 5 5 3
5 5 5 5 5 4 2 — — — 1 2
1 — 1 — 1 2 1 4 5 5 5 3
— — — — 2 5 5 5 1 5 — —
— — — — — — 3 1 2 4 3 3 — — — — — — 2 4 4 2 2 2
25 21 23 2 6 3 4 38 3 6 4 4 41 4 9 43 41
Total per animal 11 4 4 2 1 1 5 1 1 44 60 57 53 29 38 29 38 28 23 16 16 421
a smoked drum. The records were subsequently counted and analysed. Complete and incomplete extrusions were signalled without distinction, but related movements not resulting in extrusion (gulping, swaying, etc.) were not recorded.
(4) The temperature in the bath rose about 2° during each experiment. It was never below io° and never above 150 C.
Records of twelve adrenalinized animals, and twelve controls, were obtained in this way. The main results are shown in Table I. The observation period has been divided into twelve 5 min. intervals, and the number of extrusions given by each individual during each interval is tabulated. The animals have been rearranged and renumbered in order, according to the total number of extrusions shown during the observation period. In addition, the total number of extrusions carried out by all the adrenalinized animals, and by all the control animals, during each 5 min. interval, is included in the table.
As Table I shows, the behaviour of the adrenalinized animals was remarkably uniform. They all exhibited pretty regular extrusions which gradually diminished in frequency. At the same time a falling off in amplitude was noted. In most cases
150 G. P. W E L L S
the animals began by giving very vigorous extrusions, which gradually became less and less complete. As one would expect, the behaviour of the controls was more variable. One animal gave no extrusions at all; another gave only three. Others began quietly, then became active. Most of the controls began with an outburst of activity, which passed off after a period of time varying from animal to animal. The grand total of extrusions, given by all the control animals, is less than half that given by those which received adrenaline.
A feature of the records which does not emerge in Table I is that the extrusions tended to come in groups in the control animals, while in the adrenalinized animals they were more evenly spaced. This is to be expected from the behaviour of the dissected preparations. In order to give an indication of this difference, Table II has been drawn up as follows. The original records include two traces, one showing the extrusions and one showing the time in minutes. If no extrusions were shown during the interval between two signals on the minute trace, that interval is termed a " quiet minute ". The distribution of " quiet minutes " is shown in Table II. Here the difference between the two sets of worms emerges even more sharply than in Table I. The grand total of "quiet minutes" exhibited by all the controls is nearly ten times that exhibited by the adrenalinized animals.
The least satisfactory controls—numbers C n and C 12 in the tables—were continuously active for 30 min. after injection. In these two animals there were abnormalities in the extrusion process possibly due to injury during the injection. Number C 11 gave a great number of minute movements of peculiar type which were logged as extrusion; the proboscis was held partly out, and there was a curious twitching extrusion and withdrawal of a few rows of buccal papillae at the tip. There were also bursts of normal extrusion. Number C 12, in addition to normal extrusion, repeatedly nicked out the buccal mass in an abrupt, jerky way. Persistent ab-normalities of the proboscis movements were only noted in three other animals, all from the adrenalinized series. The proboscis tended to stick in the extruded position during the latter half-hour of animal A 2 and the first 20 min. of animal A 7. Animal A 1 repeatedly showed a strong contraction, lasting for several minutes, of the circular muscles of the body segments anterior to the site of injection, and during most of the observation period, in addition to extrusions, it made numerous obvious attempts at extrusion which were not successful and therefore do not appear on the record.
It is clear from these data that the action of adrenaline on the whole worm resembles that on the dissected preparations previously described. The suggestion that the contractions of the dissected preparations, initiated by the oesophagus, correspond to extrusions is therefore confirmed.
VI. DISCUSSION
of the stomatogastric system, may be termed the somatic group. The main differences in behaviour of the two groups, in dissected preparations, are as follows. The oeso-phagus shows, among other phenomena, the slow rhythmic contractions here termed primary waves, and it is excited by high concentrations of adrenaline. The proboscis, or body wall, on the other hand, if isolated from influences of oesophageal origin, show more or less regular, continuous rhythmic activity which is inhibited by high concentrations of adrenaline. If the two systems are studied together, with the normal conduction paths between them intact, the surprising result emerges that the oesophagus exerts a pace-maker action on the somatic tissues. Every primary wave of the oesophagus determines an outburst of vigorous rhythmic contractions of the proboscis and body-wall musculature of the anterior end of the worm, and after the application of adrenaline in concentrations of from 1 : 1,000,000 up, the excitation induced in the oesophagus spreads as a continuous series of regular rhythmic contractions to the somatic tissues.
Studies on the intact worm make it practically certain that the contractions induced by the oesophagus in the somatic tissues of the various dissected preparations employed correspond either (a) to extrusions and retractions of the proboscis, or
(b) in certain cases, to acts similar in nature but less vigorous in execution.
In the last section, a suggestion was made as to the biological significance of these phenomena. Our knowledge of the mode of life of Arenicola marina has recently been revised, after an extensive series of field observations, by Thamdrup (1935). The worm spends most of its life lying quietly in an L-shaped burrow with its head at the blind end, eating the surrounding mud. As fast as the mud is eaten away, fresTi supplies slide down from above to replace it. The burrow, if favourably situated, is seldom altered or abandoned. The environment of Arenicola is therefore very sheltered and constant in its properties; and the brain and sense organs of the head are greatly reduced. In view of these facts it is not very remarkable that im-pulses from the digestive tract should play a dominating part in determining the animal's activity. The worm probably alternates under natural conditions between eating and rest, the period of the cycle being on the average 6 or 7 min. and the pace being set by the spontaneously active elements in the oesophageal wall.
This "resting rhythm" can of course be modified. If for instance an Arenicola is dug out and placed on the surface of the sand, it burrows continuously downwards until it reaches a depth where the sand has a certain critical firmness (von Budden-brock, 1912, 1913). This continuous burrowing might be an adrenaline action, since adrenaline in appropriate concentrations throws the dissected preparations into continuous rhythmic activity. Whether adrenaline exists at all in polychaetes is still somewhat doubtful. The evidence for the occurrence of adrenaline in annelids is of two kinds, histological and pharmacological. As regards the first, Poll & Sommer (1903) and Poll (1908) showed that various species of leeches had chromaffine cells in the ventral nerve cord, with a characteristic arrangement of three pairs of chro-maffine cells in each ganglion. This was confirmed by Gaskell (1914), who also found the cells, arranged in the same way, in Lumbricus; when however he turned to the polychaetes, he only found the cells in two out of seventeen species examined by
152 G. P. W E L L S
him. He did not investigate Arenicola. The pharmacological evidence (Biedl, 1910; Gaskell, 1919) rests only on experiments performed with extracts of the ventral nerve cord of the medicinal leech. It is nevertheless possible that adrenaline nor-mally plays a part, either by being secreted into the blood or at the endings of adrenergic fibres, in exciting the oesophageal pace-maker of Arenicola.
Whether any mechanism for inhibiting the oesophagus exists is not known. In all my observations on the intact worms, I did not observe a single case when an animal lay perfectly quiet, without extrusions or other rhythmic head movements, for more than a few minutes. It is, however, conceivable that the slowing action on the isolated extrovert of very low concentrations of adrenaline has biological signi-ficance. In-this context, the results obtained with acetylcholine are disappointing. The great importance of this substance in vertebrates, especially as an antagonist of adrenaline at the post-ganglionic nerve endings of the autonomic system, and the occurrence of choline esters, together with cholinesterase, in two species of polychaete worms (Bacq, 1935), suggest that it might have a considerable influence on oeso-phageal activity in Arenicola. Actually, however, no very striking effect on the rhythm was detected, the clearest results being a sharp tonic contracture of the oesophagus (and not of the somatic tissues) and, in the highest concentrations used, a slight slowing of the rhythm during the rhythmic outbursts. It appears unlikely from these results that cholinergic fibres modifying the oesophageal rhythm exist
in Arenicola.
The major result of these experiments is the demonstration that spontaneously active structures in the oesophageal wall—structures which, as shown by experi-ments on the transversely bisected oesophagus and on the isolated extrovert divided in various planes, are diffuse and probably of the nature of a nerve net—play an im-portant part in determining the behaviour pattern of a polychaete worm. Hitherto physiologists have neglected the annelid stomatogastric system and have treated the nervous system of the head as if it included only the supra-oesophageal ganglion, the circum-oral nerve ring, the sub-oesophageal ganglion, and a number of sensory and motor nerves radiating therefrom. For this reason, the few published investi-gations into the functions of the various head ganglia in polychaete worms contain inferences which are not valid.
For instance, the experiments here described throw doubt on a conclusion reached by Just (1925), who studied the physiology of the nervous system in
Areni-cola marina. One of the problems investigated by Just was the effect of deprivation
Fahigkeit zum Ausfuhren spontaner Bewegungen zu liegen. Ferner iibt es mit aller Wahrschemlichkeit einen regulierenden Einfluss auf den Muskeltonus und den Mechanismus der Bewegungen aus."
As regards the first of these points—the loss of spontaneity—the fault in the argument is evident. The operations of Just cut off, not only the brain, but also the oesophageal pace-maker, from the nerve cord and body-wall muscles. Just does not describe in detail what the spontaneous movements, executed by normal worms but lost after operation, were like; it is however clear that by interrupting the con-• duction path from the oesophageal wall, which normally causes the worm to exhibit
periodic outbursts of movement—outbursts which, as already pointed out, are not always confined to the anterior end but may involve locomotor movements of the whole animal—Just put out of action an important source of spontaneous move-ments other than the brain. It is of course perfectly possible that the brain initiates spontaneous movements; but there is at present no evidence that it does so.
The other conclusions of Just, about the functions of the brain, must also be regarded with suspicion. His statement that the brain exerts a regulating influence on muscle tone rests on the observation that after operation "die Tiere, wie meist untatig auf dem Boden liegend, sich in merklich verdicktem Zustande befanden. Die Langsmuskeln waren weit iiber das gewohnliche Mass hinaus contrahiert, und das ganze Tier war kiirzer, gedrungener und straffer ". It was shown above (Fig. 20) that the outbursts of activity emanating from the oesophagus are accompanied by a slow relaxation of the longitudinal muscles of certain segments at least of the body, which pass into partial tonic contracture after the end of each outburst. It is there-fore possible that the abnormal posture observed by Just after his operations was due in some way to oesophageal influences. Once again, brain and stomatogastric system must be separately put out of action before final conclusions can be drawn. As regards the remaining point, that the brain regulates the mechanisms of movement in the trunk, Just is probably correct. He found after operation (a) that "die Wellen, besonders bei der Atem- und Ruckwartsbewegung, folgten in weit schnellerem Rhythmus als gewohnlich aufeinander", and (b) that certain abnormali-ties appeared in the form of the waves. Recently, Dr Manton Copeland (private communication) has observed a conspicuous acceleration of the rhythmic respiratory waves of Nereis virens after removal of the supra-oesophageal ganglion without injury to the nerve ring or proboscis; so it is likely that in Just's experiments on
Aremcola the acceleration of the waves was due to the interruption of conduction
paths from the brain itself.