warwick.ac.uk/lib-publications
A Thesis Submitted for the Degree of PhD at the University of Warwick
Permanent WRAP URL:
http://wrap.warwick.ac.uk/91144
Copyright and reuse:
This thesis is made available online and is protected by original copyright. Please scroll down to view the document itself.
Please refer to the repository record for this item for information to help you to cite it. Our policy information is available from the repository home page.
RESPONSES TO FOOD ODOURS MEASURED
BY BRAIN ELECTRICAL ACTIVITY
MAPPING (B.E.A.M)
IN TWO VOLUMES.
VOLUME ONE
MARTIN STUART KENDAL-REED
SUBMITTED FOR THE DEGREE OF PhD.
University of Warwick
D~partment of Psychology
TABLE OF CONTENTS VOLUME 1
Paee number
Title Page
Table of Contents
List of Figures, Tables and Plates Acknowledgements
Declaration Summary Preface
Introduction. The field of infant olfaction.
Chapter 1.
Chapter 2.
Chapter 3.
Chapter 4.
Chapter S.
Appendix 1.
Appendix 2. Appendix 3. References.
Research aims.
Anatomy/physiology and ontogeny of olfaction in Homo sapiens.
Review of the literature in the field of infant olfaction.
A model of infant cortical activity in response to 'biologically
significant' odours.
Description, results and discussion of experimental work.
Conclusions and future directions.
VOLUME 2
Additional References.
LIST OF FIGURES, TABLES AND PHOTOGRAPHIC PLATES
Fiaure, Table or Plate Number and Description.
Paae or Facina Paae Number
Table 1 - Pre-natal ontogeny of nasal chemoreceptors. Table 2 - Groupings in infant olfaction literature. Table 3 - Glossary of odours and chemicals.
Figure 1 - Diagram of systems inter-relationships. Plate 1 - Infant electrode headcap.
Plate 2 - Neuroscience Series III Brain Imager. Figure 2 - Diagram to illustrate 'head-space'.
Plate 3 - Mother & baby in laboratory during testing, Study 1. Plate 4 - Examples of Brain Imager topographic maps.
Plate 5 - Low Odour Room (LOR). Plate 6 - Odour delivery system.
Figure 3 - Low Odour Room (diagrammatic).
Figure 4 - Plan view diagram of odour delivery system. Plate 7 - Mother and baby in LOR, Study 2.
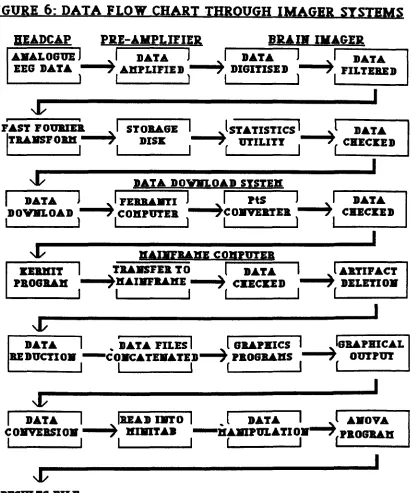
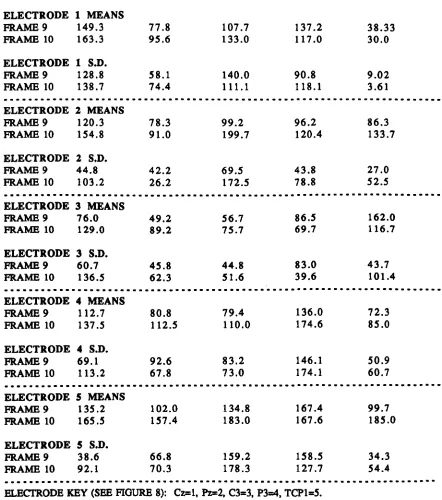
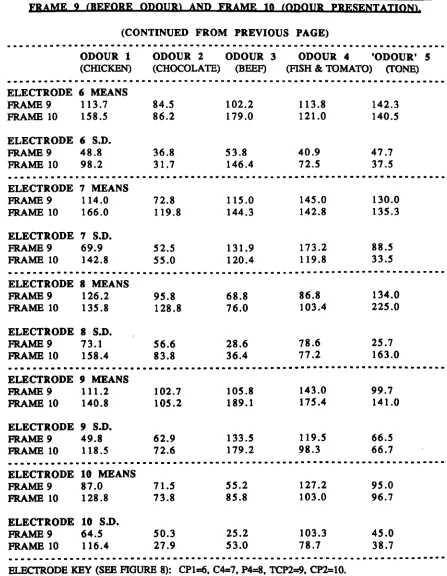
Figure 5 - Example output from graphics software. Figure 6 - Data flow through Brain Imager analysis. Figure 7 - Location map of headcap electrode positions. Figure 8 - Electrode subset used in Experimental Study 2. Figure 9 - Median standard deviations, electrode I, odour 1. Figure 10 - Median standard deviations, electrode 2, odour 1. Figure 11 - Median standard deviations, electrode 3, odour 1. Figure 12 - Median standard deviations, electrode 4, odour 1. Table/Graph 4 - Repeated-measures ANOV A, F-ratios. Table/Oraph 5 - Repeated-measures ANOV A, F-ratios (Tukey's Table/Oraph 6 - Means and standard deviations summary. Figure 13 - Cortical map (results summary).
Table 7 - Subject respiration frequency means, RP study. Table 8 - Subject inter-breath means, RP study.
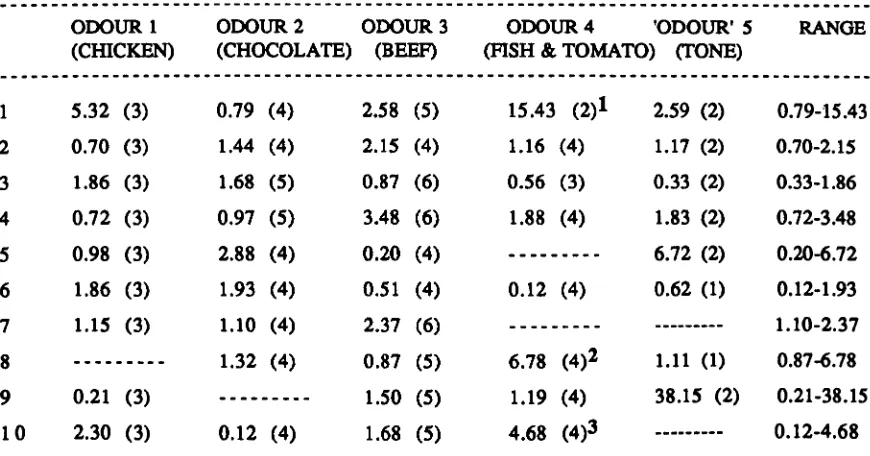
Table 9 - Response codes and operational definitions. Table 10 - Stimulus presentation summary (USA work). Figure 14 - Graphic representation of Imager data. Figure IS - Graphic representation of Imager data. Figure 16 - Graphic representation of response to tone.
vOLUME 1 Chapter I, page 21 Chapter 2, page 27 Chapter 2, pp. 29-30 Chapter 3, page 75 Chapter 4, page 82 Chapter 4, page 84 Chapter 4, page 87 Chapter 4, page 90 Chapter 4, page 93 Chapter 4, page 111 Cha,.. 4, page 112 Chapter 4, page 112 Chapter 4, page 113 Chapter 4, page 116 Chapter 4, page 120 Chapter 4, page 121 Chapter 4, page 123 Chapter 4, page 124 Chapter 4, page 127 Chapter 4, page 127 Chapter 4, page 128 Chapter 4, page 128 Chapter 4. page 132 test). Chapter 4. page 133
Chapter 4, pp. 134-35 Chapter 4, page 137 Chapter 4, page 145 Chapter 4, page 146
ACKNOWLEDGEMENTS
The research described in this thesis was generously funded by the Well come Trust, who I wish to thank for their financial support in the form of the Wellcome Trust Prize Studentship. I would like to gratefully acknowledge the constant support, encouragement and unstinting material assistance provided by my supervisor, Dr. Steve Van ToIler. His vast knowledge and expertise in the field of olfaction
was always available to me and willingly provided. The assistance
and cooperation of the following persons and institutions is also gratefully acknowledged (listed in alphabetical order):
Dr. G. Collis. Mr. K. Halstead. Dr. P. Kaplan.
Prof.
J.
Steiner.Dr.
J.
Werner.Coventry Health Authority.
Coventry Maternity Hospital (Ante-Natal Clinic Staff). Cow & Gate Limited.
Densa Limited.
Henley's Medical Supplies Limited. Neuroscience (UK) Limited.
Psychology Departments, University of Warwick, U.K., and University of Colorado at Boulder, USA., and their technical staff.
DECLARATION
The material contained in this thesis has never been previously submitted for any higher degree and represents the author's own
work. Some pilot experiments summarised at the beginning of
Chapter 4 were carried out at the University of Colorado. This work was preliminary and formed no part of studies submitted for the
degree of PhD. All remaining experimental work described in this
SUMMARY
This thesis addresses the area of human infant olfaction, which has hitherto been a somewhat neglected area in psychology. A review of the available literature showed that a number of different
experimental approaches to infant olfaction could be identified. It was concluded from this literature review that infants display a degree of olfactory competence in the weeks after birth. These previous findings are discussed in the light of a model concerned with odour significance. This model is systems-based and suggests an explanation for the apparent olfactory competence of human infants in the first weeks of life. It is argued that this may derive from pre-natal exposure to odorants and consequent acceleration of maturation in the foetal olfactory system.
The experiments reported in this thesis concentrated on the cortical reactions of infants to a small range food odours. These reactions were plotted by means of a technique involving electro-encephalography. This technique involves a computerised imaging system which summarises cortical potentials from twenty-eight locations on the scalp and is known as Brain Electrical Activity Mapping (BEAM). It is believed that this is the first time that this method has been used to examine responses to odour in human infants. This study also involved the use of a special low ambient odour testing environment. A parallel study used respiratory
plethysmography to test odour response. The major findings of the BEAM research are as follows:
1) The BEAM technique has been shown to be a practical
method in the psychophysical measurement of cortical responses to odour in the human infant.
2) Human infants at the age of three months show a pattern of cortical activity in response to a small range of food odours.
3) There is evidence that a limited area of the infant brain is responding to these odours.
It was argued that these findings lent some support to the model described above. However, similar findings to that of the BEAM work were not shown by the respiratory plethysmography study. This was explained by problems in data handling techniques.
It was concluded that the BEAM method could be used to further knowledge in the area of infant olfactory response. Possible
PREFACE
It should be noted that throughout this thesis, the terms
'infant' and 'baby' have been operationalised. 'Infant' refers to
children up to and including the first three months, and 'baby' is taken to refer to the first year of life. In line with current publishing practices to avoid sexist language, all references to one gender may
be taken to include both. All scientific units of measurement are
given in SI units. This thesis follows the presentation conventions of
the British Psychological Society (BPS) cited by: Sternberg, R.J. (1988)
The Psychologist's Companion (2nd. edition). England:
Cambridge University Press/BPS.
Each chapter is divided into sections and paragraphs. Sections
are generally identified by bold-faced, underlined headings (e.g. Method). Paragraphs are numbered according to the following
convention. The first number refers to the chapter. The second
number refers to a concept, area of discussion or topic. A new
number is generally given whenever one of these is introduced. The
Introduction to Thesis
"If I were a young researcher interested in making a career in science, especially in the area of neurobiology and the
mechanisms of the human brain: and if I was looking around for a field about which nobody knows much of anything and if I was ready, as all good researchers must be ready, to gamble
on a career in science, I would pick the problem of olfaction -and I would count on a professional lifetime of one surprise after another." Lewis Thomas, cited in Green (1988).
"In the domain of human perception, the "chemical senses" (taste and smell) are clearly the poor relations of research in neuroscience." Bruyer (1988).
"The omission of taste, touch and olfaction ... is .... an accurate reflection of the disproportionate concern which has been afforded to vision and audition in studies of infancy... Such an imbalance would detract from a comprehension of functioning at any stages of development, but may be particularly
distorting with regard to our understanding of the world of the infant. " Tiirkewitz (1979, cited in Schaal, 1988a)
"The status of the chemical senses at birth has become better understood; perhaps the real challenge now is to understand events at weaning, when the child encounters a wealth of new experience from these senses." Crook (1981).
Preamble
Much of the human infant sensorium has been investigated by
psychologists in the last hundred years. A considerable amount is
known about infant visual, auditory and somatosensory perception although, of course, much remains to be learned (Werner & Lipsitt,
1981). Despite these advances, a whole area of infant sensory
experience has been relatively neglected. Human infant olfaction
olfaction in general has been under-emphasised by science. As
Lewis Thomas described it: "a field about which nobody knows much
of anything". A good test of this contention is to consult most general
textbooks on sensory psychology published in the last 40 years. The
chances are that human olfactory perception will merit a sub-section of a chapter, or even as little as a paragraph. The empirical evidence is likely to be sparse, even inaccurate (though there are exceptions). The section usually closes with a dismissive statement that nothing much is known about the area, with the clear implication that it is not important anyway.
However, interest in the 'chemical senses' has started to grow
in the last few years. The formation of research organisations like
the European Chemoreception Research Organisation (ECRO), Association for Chemoreception Sciences (AChemS) and Japanese Association of Smell & Taste Scientists (JASTS) has helped to promote this growth of interest in a hitherto neglected field of human
experience. However, whilst the field of adult olfaction has seen a growth of interest, the area of human infant olfaction has lagged behind. The principal aim of this thesis has been to play a small part in improving knowledge about this area.
Aims of the thesis
The goals of this thesis were essentially four-fold and are
described below in the order in which the chapters appear. The first aim was to try and describe and integrate what little is known about
the biological development of human infant olfaction. This is to be
found in Chapter 1. The purpose of this chapter was to provide a context for the research programme by giving an understanding of some of the biological mechanisms involved in infant olfaction. Since the research involved addressing broadly 'psychobiological' events , description of the biological substrates provided a background for the understanding of these events.
The second aim was to examine the literature in the field of infant olfaction, delineate trends and schools of thought and trace the
development of thinking in the area. This material compnses
context of the research programme. By tracing the evolution of the various research trends, mainly in this present century, the rationale for the experimental work described in this thesis could be better established.
The third aim of this thesis was to describe a model which might account for the relative olfactory competence displayed by
infants. This competence has been demonstrated in many of the
studies described in Chapter 2. The model is based on 'systems thinking' and proposes that pre-natal olfactory influence may occur in the human infant. The model is biologically based and relates to the substrates described in Chapter 1.
The fourth and ultimate aim was to describe a number of experiments involving objective measurement of cortical and other psychophysical activity in young human infants presented with food
odours. This mainly methodological aim is encompassed in Chapter
4, which comprises the bulk of the thesis. The use of a relatively new technique, which became available only in the 1980's (Brain Electrical Activity Mapping, or BEAM), is evaluated in the testing of infants. The main aim was to examine whether the BEAM technique represented a usable tool for investigating infant olfactory response. This was confirmed, despite considerable technical problems.
The final chapter draws together the various strands described
above. The main conclusions of the experimental work are discussed
in the light of the original hypotheses discussed at the beginning of
Chapter 4. Future directions are examined and some likely courses
CHAPTER 1
ASPECTS OF ANATOMY, PHYSIOLOGY AND ONTOGENY OF OLFACTION IN HOMO SAPIENS
Introduction
1.1.1. This chapter aims to summarise some of the knowledge about the anatomy, physiology and neurological development of the human olfactory system. The reason for doing this is to set a context for the empirical research described elsewhere in this thesis. By attempting a synthesis of what is known, it is hoped that a clearer picture of the biological substrate to infant olfaction will emerge.
1.1.2. Human olfaction is an immense area, which is very diverse in nature. A full understanding of the olfactory system in Homo sapiens is likely to derive only from a multi-disciplinary, eclectic approach. This is because each level of the system contains elements relevant to several disciplines. It is not at all clear what the boundaries and degree of overlap of each speciality are. For example, the physical nature of the odorants is partly the domain of organic chemistry, whilst their olfactory transduction falls within the expertise of biochemists and molecular biologists. At a 'higher',
cortical level, odour perception may one day be explained by electrophysiologists, neuroanatomists and psychologists. This chapter attempts to combine a little of the knowledge of each discipline, taking the view that any detailed approach to olfaction requires evidence from living systems wherever possible.
and autoradiography that have yielded so much information from animals are never likely to be replicated in humans, not merely
because of the obvious ethical problems. Because of this, psychology
has a major part to play in understanding Structure-Activity relationships.
1.1.4. Biochemists have long used animal models to try and comprehend how structure in the olfactory system is related to
function. This can be done by manipulating the animal's ambient
odour environment, then sacrificing the animal to examine the often
subtle changes in the olfactory biochemistry. Clearly, such
experiments are not possible in humans. Hence, scientists have to
rely upon behavioural changes to understand the effects of odour. Psychology clearly has a major part to play here. With the use of psychophysical and psychometric techniques, insight into structure-activity relationships can be inferred.
1.1.5. Because much of the work in the area of olfactory
anatomy and physiology has been done with rodents and other small mammals it is always difficult to know to what degree cross-species generalisations are valid. The position taken in this thesis is that, although animal models provide a framework for understanding human olfaction, cross-species generalisations to humans need to be cautious. Because of this, animal work will only be referred to when there is an absolute lack of human evidence.
1.2.1. This chapter is subdivided into the following sections:
Section 1: gross anatomy, micro-anatomy and ultrastructure
of the human olfactory system.
Section 2: physiology of the human olfactory system.
Section 3: development of the human olfactory system.
and physiology that can be found in some recent works; notably Barr
& Kiernan (1983), Castellucci (1985), Macrides & Davis (1983), Moulton (1978), and Van ToIler, Dodd & Billing (1985). Specific discussion of comparative micro- and ultrastructure is gleaned,
unless otherwise identified, from the article by Moran et al (1982).
Histological data from infants comes mainly from the 1984 study by
N akashima, Kimmelman & Snow.
Section 1
Gross anatomy and micro-anatomy of the human Qlfactory system.
1.2.2. It should be made clear that relatively little anatomical
work has been done on the human infant olfactory system. This is probably due, in part, to the ethical problems of obtaining the
necessary autopsy material. These prefatory remarks should serve
as a caveat over the problem of comparing the adult system with
that of the infant. The major differences are probably those of scale
and size. A paper by Laitman & Crelin (1976) contains photographs
of autopsy material as well as airway casts of infants, children and adults. The major differences in size and shape can be clearly seen. The anatomical relationships of the structures are therefore
different. This has been noted by Cole (1982), amongst others.
Differences in shape and size mean that the flow patterns of air, and hence odorants, in the nasal cavity are probably quite different in
the human infant from those found in adults. Very recent evidence
suggests such differences between children and adults (Mennella &
Beauchamp, 1990a). These may have implications for
chemoreception in infants. Nonetheless, the scarcity of cadaver
, material and consequent lack of published work on the normal anatomy seriously hinders investigation of the human infant olfactory system. For this reason, this section will concentrate on evidence gleaned mainly from adult subjects, with comparative anatomical input. from infant studies where possible.
Gross Anatomy.
1.2.3. As with all sensory systems, structure subserves
airflow patterns in the human nasal cavity have been mapped, and it has been found that the anatomy of the nasal cavity serves to
perturb the mainly laminar flow of inspired air in the anterior portion (Proctor & Swift, 1977; cited in Cole, 1982). Because of this, turbulence is created, which is necessary to physiological functioning. This is stated by Cole (1982, page 173):
"The energy required to promote and maintain the disturbed state of respiratory air is not a waste of energy, these
characteristics of airflow are of physiological importance. If they did not occur, a boundary layer of air would insulate the main airstream from the mucosa".
1.2.4. Clearly, this would have implications for the diffusion of odorous molecules into the mucosa of the olfactory neuroepithelium, which is the necessary precondition for transduction and
chemoreception (Berglund & Lindvall, 1982, page 281). Stuiver
calculated that, in any case, only 2% of the molecules entering the nasal cavity actually make contact with the olfactory receptors
(Stuiver, 1958; cited in Gescheider, 1985). However, this investigator
may not have taken into account the turbulence of the airflow which would predispose to a greater number of receptors being stimulated
(Saito & Nashihata, 1981, mentioned below).
1.2.5. The human nasal cavity subserves several functions,
only one of which is concerned with olfaction. For example, according to Eccles (1982):
"The nose therefore acts as a filter, a humidifier and a heat exchanger... the nose functions as an air conditioning system" (page 192).
Hence the anatomy has evolved to cope with these functions, as well as olfaction. The heavy vascularisation of the nasal cavity and the presence of mucous-producing goblet cells are an example of
structure subserving function. According to Tos (1982), the
variations of airflow within the cavity. Tos further states that the density of goblet cells is lower in infant and children than in adults
(Tos, op.cit.). The reasons for this may be connected with the
differences in airflow patterns caused by the differences in anatomy, mentioned above, in younger subjects.
1.2.6. In the human infant, the same 'air conditioning'
functions are necessary; probably to a greater degree than in the
adult. The reason for this is the much smaller lumen of the nasal and bronchial passages which need to be kept patent at all costs to allow
un impeded respiration. This is why upper respiratory tract
infections in babies, especially those which cause oedema of the mucosa (croup, for example), may be more serious than in adults. Indeed, as stated by Cole (1982):
"Partial nasal obstruction has been suggested as a cause of infant cot deaths" (page 165).
This is probably because, as noted by Cole: "Infants are obligatory nose breathers" (op. cit., page 165). This author contends that it is not possible for infants to use the mouth for breathing until the age of 5 to 6 months (op. cit., page 164). If this is so, then the infant
olfactory system will, ipso facto, receive continual stimulation during
the first months of life. However, work on models of the adult nasal cavity suggests that, unless actual 'sniffing' is employed, very little odorant reaches the receptors (Stuiver, 1958; cited in Gescheider, 1985). Infants have clearly not yet learned this ability in their first
few months. Nonetheless, as no empirical work appears to have been
done on the airflow patterns in the infant nasal cavity, it is unknown approximately how much of any odorant is likely to reach the
receptors. Hence, it is not yet possible to either confirm or refute Stuiver's work. However, a contrary view is taken by Saito &
Nashihata, (1981; cited in Schaal, 1988a). These workers suggest that the higher airway resistance induced by the narrower airway passages in the infant nasal cavity may actually improve aeration of
the olfactory neuroepithelium. This was suggested as early as 1932,
generally low detection thresholds demonstrated by infants (Rovee, 1969; Balogh & Porter, 1986)
1.2.7. Turning to the olfactory region, the structure of this area is arranged so as to receive inspired odorants on the incoming
airflow, via the olfactory cleft. Hence it is necessary to know
something of the physical properties of the nasal cavity, in order to
understand how odorants arrive at the olfactory mucosa. The
'aerodynamic' properties of the nasal cavity have been investigated, as mentioned above. The airflow is a function of size, shape and
mucosal conditions as well as inspiratory velocity. In the adult, the
nasal mucosa consists of about 160 cm2 , with a nasal cavity volume
of approximately 20 ml. The cross-sectional area is about 130 mm2•
This results in a transit time of any given portion of inspired air of about 1120 of a second, during quiet breathing. This is partly due to the shape, and hence resistance of the anterior nasal cavity,
permitting an airflow velocity of up to 18 metres/second in the adult. All these figures (Cole, 1982) are for adults, as no information is readily available for infants. This is despite a study by Nakashima et al (1984) which studied late foetal material. However, the main point is that the inspired airflow, laminar at the external nares,
becomes turbulent within the nasal cavity. This ensures dispersion
and mixing of any odorants which reach the nasal cleft. In the adult human, this is located at the most superior portion of the nasal cavity
and is highly inaccessible to in vivo observation or biopsy. As stated
by Moran et aI, (1982) this is:
"A tiny 1.5 mm crevasse between closely apposed nasal bones (turbinates and septum) ... that lies within the recesses of the skull some 7 cm deep to the skull" (page 722).
In human infants, no figures are available. However, it is reasonable
to assume that the olfactory cleft is considerably smaller because of the overall smaller skull size, even though the infant skull is
proportionately larger than the adult. This may well have
1.2.8. A further difference between the adult and foetal/infant nasal cavity is the vomeronasal (Jacobson's) organ. This can be seen in the foetal and infant nasal cavity, but rarely in the adult according
to at least one worker (Humphrey, 1940). However, Johnson et al
claim that it can be found in a large proportion of adults (Johnson,
Josephson & Hawke, 1985; cited in Schaal, 1988a). The vomeronasal
organ has excited anatomists for a long time, because of its apparent resemblance to a similar organ found in other species, such as
marsupials, ungulates, reptiles, rodents and carnivores (Moulton,
1978). For example, it is believed that the nervi terminales serves to
innervate and perhaps subserve an olfactory function in the
vomeronasal organs in these species (Eccles, 1982). Certainly, these
nerves exist in Homo sapiens and run alongside the olfactory tracts.
However, since the vomeronasal organ is considered vestigial, or
even absent in adults, the function of the nervi terminales is unclear
(Moulton, 1978; Widdicombe & Wells, 1982).
1.2.9. The vomeronasal organ has been described in the human
foetus, as late as the seventh gestational month (Nakashima et aI,
1984) and even in neonates (Read, 1908 ; Peter, 1925; cited in Schaal,
1988a). These structures are found bilaterally on the anterior part
of the nasal septum. They are described as:
"tubular... [and] completely separate from the respiratory epithelium except at their opening into the nasal cavity anteriorly.... The vomeronasal organ has an oval lumen.... The cellular distribution of the vomeronasal epithelium is similar to that of the olfactory neuroepithelium" (Nakashima et aI, 1984, page 644).
The function of this structure, especially at a late stage in human
foetal development, is obscure. There is also controversy
surroundings its functions in the species listed above (Moulton, 1978). For this reason, it is not proposed to speculate on its
Micro-anatomy.
Olfactory mucosa: ultrastructure by cell types.
1.3.1. The olfactory mucosa, which contains the olfactory
receptor cells and their associated cell types, is confined to an area of about 2 cubic centimetres. There is disagreement in the literature about this figure. Some modem estimates give a much lower figure than earlier studies, which suggested an area of 10 cm2 (e.g. von Brunn, 1892; cited in Mouiton, 1978). However, even modem estimates show a considerable degree of variation. For example, Castellucci (1985) gives a figure of 5 cm2, whereas Jenner & Dodd (1988) suggest 120 cm2 . This is the figure for the mature adult; no figures were available from the foetal study by Nakashima et al (1984). The olfactory mucosa contains four types of cells (Morrison & Costanzo, 1990), which will be discussed in some detail, in order to make the point that there seem to be micro-anatomical differences between infants and adults. The four cell types are:
1) microvillar cells
2) ciliated olfactory receptor cells 3) sustentacular cells
4) basal cells and the lamina propria
1.3.2. Olfactory epithelium is a pseudostriated columnar type resting on highly cellular lamina propria. The mean thickness of this epithelium is approximately 70 microns. In the human infant, the epithelium is described only as:
"thick and highly cellular... [with] a zonal distribution of supporting [sustentacular], sensory receptor, and basal cells" (Morrison & Costanzo, 1990, page 643).
ai, 1984). It might be speculated that this zonal disruption could be the result of long-term exposure to atmospheric pollutants.
Furthermore, recent studies have shown that the olfactory
epithelium in adults may contain respiratory epithelium as well, in a complex mixture (Morrison & Costanzo, 1990; Paik, Lehman, Smith &
Seiden, 1990). The epithelial surface in both adults and foetuses is covered by a layer of mucous, through which odorants have to pass. The role of this mucous layer is crucial to olfaction and will be
discussed later.
1.4.1. The outermost cell type encountered in the olfactory epithelium is the recently-identified microvillar cell. These cells are flask-shaped, with a narrow neck at the cell apex, which is
equipped with short, straight microvilli that project into the mucous
layer. They are usually abundant but solitary, though pairs are
sometimes seen under electron microscopy. The cell population in
the average human is estimated at 600 000. Their function is
unknown, though they bear a resemblance to cell types known, from other species, to respond to odorants (Graziadei & Tucker, 1970; cited
in Moran et ai, 1982). Indeed, they apparently look like bipolar
sensory neurons, so it may not be unreasonable to ascribe a
chemoreceptive role to this cell class. However, it should be noted that not all authorities recognise this cell type as being discrete. Anholt (1989), for example, fails to mention it in his review paper, referring to only three cell types.
1.4.2. The actual sensory cells are the ciliated olfactory receptor cells. These cells are highly specialised, with several highly unusual features (Anholt, 1989), some of which will be
addressed below. Anholt speaks of
the:-"remarkable plasticity and unusual morphology... several unique biochemical and electrophysiological properties".
of olfactory receptor cells. The density of these cells is somewhat hard to estimate, because of the supposed turnover rate, but has
been given as 30 000 receptors/mm2 . This gives a likely mean
should be noted that Nakashima et al (1984) state that it is
unknown whether human olfactory receptor cells 'turn over' in the
same way as has been reported in other species. Other workers have
suggested different estimates of the number of olfactory receptor cells. Van Toller et al (1985) give an estimate of 10 000 000 olfactory receptors, though Castellucci (1985) gives a figure of ten
times that amount. These varying estimates serve to highlight the
problem of determining the number of olfactory receptor cells. However, the density of these cells appears to be high in the
neuroepithelium, with a figure of one receptor every 3 to 5 microns.
1.4.3. The morphology of the receptor cells is reasonably well-documented from adult studies, though there is little comparative work for infants. In summary, each receptor cell is a long, slender bipolar neuron, with a dendrite sent to the surface of the epithelium and an axon sent to the olfactory bulb. Receptor cells are about 42 microns long, not including the axon. The tip of the dendrite that projects into the mucous layer has a specialised structure; a
bulb-shaped modification called the olfactory vesicle. This structure is
about 1.5 microns in diameter and projects some 2 microns above the epithelial surface. It is this which terminates in the olfactory cilia.
1.4.4. The cilia attached to the receptor cells are believed to great~y increase the surface area of the cells, which presumably increases the receptive area available for odour molecule
transduction (Berglund & Lindvall, 1982; Anholt, 1989). Each cell
has between 10 and 30 of these cilia, which are presumed to be involved in the binding of odorous molecules and hence sensory transduction (Ohno et ai, 1981; cited in Moran et ai, 1982; Anholt, 1989). Further evidence for the role of these cilia in olfactory
transduction comes from animal work described by several workers
(Berglund & Lindvall, 1982; Cagan and co-workers, cited in Moran et
ai, 1982; Dionne, 1988). However, the exact mechanism of binding,
1.4.5. As much of the work on the role of the cilia comes from animal studies, it is probably premature to generalise to humans, and
particularly infants. Aside from the differences in distribution of
cells between human adults and foetuses, there is no evidence for any fundamental disparity in chemoreceptive mechanisms and
structures. There is histological evidence of ciliated dendrites from
foetuses as young as 14 weeks post-gestational age (Chuah & Zheng,
1987). The implication from this and the other studies is that the infant olfactory neuroepithelium is morphologically similar to the
adult type, but more sensitive to odorants. It could be speculated
that this is due to its comparatively 'pristine' state, in that infant epithelium is both highly plastic and has not yet been exposed to the destructive effects of airborne pollutants.
1.4.6. A further distinctive property of olfactory receptor cells
is that of a unique cytoplasmic protein called 'olfactory marker
protein', or OMP (Margolis, 1972, cited in Nakashima et ai, 1985;
Anholt, 1989). It was originally isolated in rodents, but has also been found in humans, both adult and in the later foetal stage (Nakashima et ai, 1985; Chuah & Zheng, 1987). It is found
throughout the olfactory receptor neuron and is reported to be useful in discriminating olfactory from respiratory epithelium, though its
function is unknown in either infants or adults (Nakashima et ai,
1985). Perhaps OMP has some role in dealing with the breakdown products of odorant molecules following transduction, along with cytochrome P-450, mentioned below.
1.5.1. The third cell type found in the olfactory epithelium consists of the supportive or sustentacular cells. These will not be
discussed in detail, though Moran et al (1982) describe them
as:"morphologically intriguing". These supporting cells lie between and around the receptor cells in the epithelium, showing a 'tight
junction' relationship. It may be that they perform a supportive and
nutritive function analogous to neuroglial cells in the brain.
However, 'gap' junctions, which would allow electrical coupling of
adjacent cells, have yet to be demonstrated. The sustentacular cells
do not secrete mucous, though this is not the case in the same class of
the sustentacular cells may be equipped for enzyme-based
degradation and axoplasmic transport of inspired odour molecules
and their metabolites (Moran et ai, 1982). This is an intriguing idea
which raises the possibility that repeated exposure to cytotoxic breakdown products acquired during lifetime inhalation of odorants may lead to the disruption of the neuroepithelium seen in the adult but not the foetus. This may be a microstructural analogue of
'presbyosmia', which is the age-associated increase in
odour-detection thresholds described by Van ToIler et al (1985). However,
a cellular transport mechanism is not the sole candidate for this process of odour molecule degradation, mainly because the sheer
diversity of molecules makes a unitary system less likely. An
intra-cellular enzyme (cytochrome P-450 mono-oxygenase) has been
suggested as a candidate for this process (Van ToIler et ai, 1985). For a review of the role of this enzyme, see Jenner & Dodd (1988). It is a powerful oxidiser of organic molecules, principally found in the liver, where it performs a detoxification role. It is also found in high
concentrations in the olfactory mucosa.
1.5.2. The fourth cell type is the basal cell. These are typical 'stem' cells that are the progenitors of the other cell types. The undifferentiated cell is small (4 to 6 microns, about the size of an erythrocyte) and located on the basement membrane, or lamina propria. This structure, which is highly cellular, is a framework of
collagenous connective tissue unlike a true submucosa. It contains
blood vessels, connective tissue cells, nerve bundles and Bowman's (secretory) glands. One of the main functions of the lamina propria seems to be guidance and support, especially during neurogenesis, for the afferent axons of ciliated olfactory receptor cells. It is within the lamina propria that these axons form large bundles and travel to the cribriform plate, en route to the olfactory bulb.
1.5.3. There is no evidence that this state of affairs is any different in the infant nasal cavity. It is not known, for example,
whether the basal cells are more active or numerous. However, if
human foetus and young infant. This would tend to be reduced over time as connections became established and neuronal die-back
occurred. This hypothesis awaits the necessary comparative
ultrastructural studies.
Section 2
Physiology of the human olfactory system
1.6.1. This chapter has already provided an overview of how odorous molecules are brought as far as the mucous layer overlaying the receptor cells. This section will deal with the physiology of the human olfactory system, with special reference to what is known about the infant system. It is not proposed to deal with this highly complex subject in great depth, as this has already been
accomplished in an excellent review by Anholt (1989). Like any other human sensory system, the olfactory sense is set up to collect, transduce, process and transfer incoming environmental information. This section will consider the molecular events surrounding
transduction, as well as briefly considering some of the various theories of odour transduction as applied to both adults and infants.
1.6.2. In order for any transduction to occur, odour molecules must first diffuse through the mucous layer to reach the cilia of the
receptor cells. It should be noted that various authorities have
calculated the possible number of odorants that the human system
may be sensitive to. According to Dionne (1988), there may be 100
000 different types, though Engen (1982) suggests 400 000. Although most perceived odours are complex mixtures, pure odorants exist and it is the physical features of the molecule that partly determine its ability to reach and stimulate the receptor (Van ToIler et ai, 1985, page 19).
1.6.3. One of the problems surrounding this event is encapsulated by Van ToIler (op.cit.):
"The first step in the molecular sensing mechanism is the
If the fine structure of the mucous is obscure, then one of the earliest
steps in odour transduction is unknown. However, Van ToIler et al
go on to suggest that the mucous comprises a solution of
glycoproteins (proteins with chains of sugar molecules attached to them) in an aqueous solution, resembling mucous found on other
epithelia in the human body. Water-soluble odorants will dissolve
readily in the mucous, as will highly volatile lipid-soluble molecules. In this way, the essentially biochemical attributes of an odorant molecule may determine its ability to reach and stimulate the
olfactory receptors. Hence the physical characteristics of an odorant
may contribute to its detection threshold. Furthermore, the
thickness and viscosity of the mucous layer may vary both spatially and temporally in adult humans, thus exhibiting a kind of cyclical
variation (Mair et ai, 1978; cited in Engen, 1982). This may further
affect the amount of odorant reaching the receptor cells. No evidence is available in the human infant for this view.
1.7.1. How and where odorants bind to the olfactory receptors
is apparently still an unanswered question (Bignetti et ai, 1988;
Anholt, 1989). This is especially the case when evidence from
Dionne (1988) is considered. This worker suggests that there may be no such thing as a special protein existing just to bind odorants. As Van ToIler et al (1985) put it:
"The first obvious question is whether or not the stimulant molecules bind to a receptor on the outside of the nerve cell or whether they pass into the cell and bind to an intracellular receptor" (page 23).
transduction will be replaced by a discussion of the broader area of olfaction theories. The reason for doing this is to provide a context for how infant olfactory perception may operate.
1.8.1. There are a number of theories of olfaction. A selection of the more well-known of these will be addressed. One, at least, dates back to Roman times. The poet Lucretius put it this way:
"You may readily recognise that those bodies which touch our bodies pleasantly are made of smooth round atoms, but
contrariwise all that seems to be bitter and rough are held in connection by atoms more hooked, and are therefore wont to tear open their way to our senses and to break the texture by their intrusion" (cited in Van ToIler et ai, 1985).
1.8.2. Several theories have extended Lucretius' idea of
specific shape being responsible for perceived odour. Amoore has
presented the most elaborate and closely-reasoned of these (Amoore, 1967; cited in Berglund & Lindvall, 1982). In summary, this theory states that odour molecules attach to molecular receptors by virtue of their shape. This has been called a 'lock-and-key' view, in that the configuration of the molecule is responsible for its ability to attach to the receptor site and thus induce a change in cell potential, which amounts to transduction.
1.8.3. The actual molecular events which occur as a result of
binding are more difficult to describe. Studies of other sensory
systems have shown that the critical events surrounding
transduction are biochemically mediated, as in the visual system (Carlson, 1981). There is no theoretical reason to assume a radically
different arrangement for the olfactory system (Van ToIler et ai,
1985). However, Dionne (1988) disagrees. It is apparent that the
solution to this conundrum awaits detailed biochemical evidence. Some of this is given by Anholt (1989), who provides a detailed discussion of the ionic events accompanying odorant binding.
study of odour transduction, even though no such highly specific
proteins have been found. Another theory suggests that it is the
spatiotemporal distribution of odour molecules within the mucosa that determines the perceived odour, which is analogous to
chromatography (Mozell, 1970, cited in Berglund & Lindvall, 1982).
However, there is nothing to preclude an eclectic mixture of these models, as Berglund & Lindvall (op.cit .. ) state.
1.8.5. Further theories include that of Wright (1954, cited in Berglund & Lindvall, 1982). This is the so-called 'vibrational
hypothesis' which states that the amount of resonance of an odorant molecule determines the degree of transduction by specialised
pigment molecules in the olfactory mucosa. This hypothesis has been
further argued by Briggs & Duncan (1962, cited in Berglund &
Lindvall, 1982), who claimed that these pigment molecules are
analogous to those found in the visual photoreceptors. Another
theory, which also relates to the physical characteristics of the odorant molecules is that of Laffort et al (1974, cited in Berglund &
Lindvall, 1982). The physico-chemical properties of the odour
molecules, combined with characteristics of receptor proteins, operate in conjunction to determine the receptor membrane
potentials. There is apparently some empirical evidence for this
hypothesis.
1.8.6. Lastly, Holley & Doving (1977, cited in Berglund &
Lindvall, 1982) suggest a complex model which relies upon the existence of odorant 'acceptors', which provide a spatiotemporal
patterning to encode odour quality. Recent work in the theoretical
and mathematical modelling of electrophysiological events in the
olfactory bulb, based on Chaos Theory (Skarda & Freeman, 1987)
have moved even closer to an eclectic approach. This research
represents an attempt to explain olfactory coding, discrimination and
odour memory. It attracted numerous criticisms (e.g. Brown, 1987;
Corner & Noest, 1987), so it is probably fair to say that Skarda &
Freeman's work is not yet proven. Nevertheless, the main value of
this work is that it provokes thought about how to conceive the
help us to consider the 'chaotic' nature of human EEG in response to odour.
1.8.7. In terms of the infant olfactory system, there is no evidence to support either one, or a combination of the above
theories. However, the major difference between the infant and
adult systems is likely to be the highly plastic nature of the former. There is considerable evidence that post-natal neuronal
interconnection is influenced by chemosensory experience, though this evidence derives from animal research, so caution is required in
generalising to the human system. This aspect will be considered in
the following section, which discusses the developmental aspects of the human olfactory system.
Section 3
Deyelopment of the human olfactory system
1.9.1. Much of the evidence in this section derives from embryological work, in particular that of Lemire, Loeser, Leech &
Alvord (1975). It should be emphasised that relatively little is
known about the embryological development of the human olfactory system. Muller & O'Rahilly (1989) succinctly put it thus:
"The embryology of the human brain is very poorly
documented and illustrated in contrast to that of experimental animals" (page 353).
These and other authors are usually not primarily concerned with
olfactory development per se, but the system is often considered
interesting to those studying the ontogeny of the brain and central nervous system. This is largely because of its relative accessibility in
experimental animals. Furthermore, the olfactory system is usually
considered to be one of the phylogenetically oldest, and can thus be
considered as a precursor to neocortical development. Some
anatomists have studied discrete areas of the olfactory system during development (for example; Humphrey, 1940), but not
attempted an integration of the whole area. Moreover, most authors
the human olfactory system. This is corroborated by Gottlieb (1973; cited in Schaal, 1988b) who, referring to the chemoreceptive senses, states: "there is little or no information on their development".
However, what is known in terms of anatomical and neurological development will be summarised. The aim of this is to provide a background for the model described in Chapter 3 and the research described in Chapter 4.
1.9.2. The first signs of differentiation in the embryological olfactory system are the olfactory placodes, which appear at about the fourth gestational week. This corresponds to stage XIII of Streeter's embryological coding system, which will be used in this section (Streeter, 1942, 1945, 1948: cited in Lemire et ai, 1975). Olfactory placodes are described by Costanzo & Graziadei (1986) as:
"Paired thickenings of the cranial ectoderm .... The nervous layer of the olfactory placodes gives rise to the receptors of the sensory epithelium; the non-nervous layer of the placodes
gives rise to the supporting cells and Bowman's glands" (page 234).
The first bipolar cells resembling olfactory nerves appear in the olfactory placode by stage XVI (about 35 gestational days) and the olfactory nerve proper is defined by stage XIX (about 50 days). The olfactory bulb begins to assume its adult macro- and micro-structure by stage XXIII, or nearly 60 gestational days. Differentiation is
completed by about 100 days.
1.9.3. Several other anatomical entities appear and then
apparently become vestigial during this period. These include the
olfactory ventricle, the accessory olfactory bulb and the vomeronasal
organ. It is not known what function these 'proto-organs' subserve,
or why they degenerate into a vestigial state during embryonic
development. However, it is known that the anterior nares are
plugged by epithelium (Schaffer, 1910, cited in Schaal, 1988a). Smith (1976) states that this occurs from about the eighth week of
gestation, until the twenty-fourth week, when they reopen. No
olfactory receptors from amniotic chemosensory stimulation during a
crucial period in neurogenesis. The proto-nasal cavity (or 'nasal pit')
is evident at stages XV and XVI, but develops into a separate nasal sac by stage XVII, about 41 post-ovulatory days (Muller & O'Rahilly, 1989).
1.9.4. The development of the micro- and ultrastructure of the
foetal olfactory system is less well documented. Post-natal
development is even more obscure, probably because of the paucity of cadaver material alluded to at the beginning of this chapter.
However, the olfactory receptor cells may be identifiable by the ninth to eleventh week of gestation (Schaal, 1988a, page 147). The cell types described in Section 2 can be demonstrated soon after the end of the first trimester of pregnancy, according to one study
(Chuah & Zheng, 1987). This may allow early receptor function to be inferred, as suggested by Pyatkina (1982, cited in Schaal, 1988a), as well as Gesteland et al (Gesteland, Yancey & Farbman, 1982; cited in
Schaal, op.cit.). The appearance of olfactory marker protein in older
foetuses, demonstrated by Chuah & Zheng (1987), may be a
biochemical indicator of receptor cell functioning. This is certainly
the case in foetal rats, though it may be premature to generalise to
humans. However, Shepherd (1988) states that the olfactory system
is one of the most precocious systems in early embryonic life. The implications of this precocious functioning are discussed at length in Chapter 3 of this thesis.
1.9.5. In humans, connections with the olfactory bulb and
hence the rest of the brain, appear somewhat later. Using olfactory
marker protein as a 'tracer' , it has been shown that the olfactory
bulb seems to become 'wired up' at about 32 weeks (Nakashima et ai,
"the differentiation of the human olfactory epithelium begins relatively early as compared to other mammals such as the rat in which the receptor and supporting cells can only be
identified in the second half of gestation" (Chuah & Zheng, 1987).
No great claims may yet be made for this finding, as the possibility of functioning receptors does not necessarily imply a fully functional,
cortically interconnected chemosensory system. Nonetheless, it may
have implications for prenatal olfactory experience in Homo sapiens
as discussed in Chapter 3 of this thesis.
TABLE 1; Pre-natal optogeny of nasal cbemoreceptors in bumans (adapted from Scbaal. 1988a)
---Gestational Ue
(post-ovulatory
week)
3.5
-
5 4.5-
6 4.5-
7 5 • 85.5 13 5.8
.
8 6·
6.5 6.5 7·
87.5
.
9.59.5 1 1
11 18.5 16
·
24 24·
term32
·
35Formation of the olfactory placode (2,4,12)
Formation of the nasal grooves (4,12)
Differentiation of the Olfactory nerves (4,12)
Formation of the vomeronasal grooves (2,4,12)
Presence of olfactory-like cells in the vomeronasal regions (13,14)
Terminal-like vomeronasal pathways differentiated (2,4,8,12,13)
Formation of the main olfactory bulbs (2)
Formation of the accessory Olfactory bulbs (8,13)
Characteristic structure of the main olfactory bulbs delineated (2)
Opthalmic & maxillary divisions of the trigeminal nerve
differentiated (5,6,9,10)
Increase of the size of the mitral cells in the main olfactory bulbs (8)
Presence of ciliated olfactory receptors suggested to be ready for
reception (1,15)
Mitral cell layer clearly delineated in the main olfactory bulbs (8)
Presence of epithelial nasal plugs in the external nares (16)
Resolution of epithelial nasal plugs in the external nares (16)
Olfactory marker protein present in the olfactory neuroepithelium, the
olfactory nerve and the bulbar glomerular layer (3)
---. /
References: 1. Arey (1930), 2. Bossy (1980), 3. Chuah & Zheng (1987), 4. Gasser (1977),
S. Gasser & Hendrickx (1969), 6. Hogg (1941), 7. Hooker (1952), 8. Humphreyl.(1940), 10.
Humphrey (1978), 11. O'Rahilly (1967), 12 O'Rahilly tt al (1981), 13. Pearson (1941),
[image:32.559.79.508.239.584.2]1.9.7. The post-natal neurological development of the human infant olfactory system is difficult to assess directly. As Schaal (1988a) puts it:
"What we know about the ontogenetic interrelations between structural and functional correlates of human olfaction remains disconcertingly poor" (page 148).
Histological studies of the postnatal anatomy are very rare indeed. Steiner (1979, cited in Schaal, 1988a) mentions occasional cases of gross eNS malformation which responded post-natally to odorants,
but no micro-anatomical details are given. However, inferences from
animal studies suggest a predisposition to plasticity which may well also be valid in the human. As Shepherd (1988) puts it:
"lts early-developing functional properties display a
considerable plasticity whose effects can last well into adult life" (page 240).
1.9.8. The mechanisms underlying this remarkable neuronal
ability to adapt structurally to sensory experience are now being
uncovered. For example, molecular biology has suggested that
certain proteins found in cortical neurons may underpin this ability (Aoki & Siekevitz, 1988). These authors provide a clear analogy of the process of neuronal plasticity:
"The developing brain can be likened to a highway system that evolves with use; less travelled roads may be abandoned,
popular roads broadened and new ones added where they are needed" (page 34).
1.9.9. It is not known whether the effects of the degree of enrichment of the olfactory environment influence the degree of plasticity, in the manner of classical studies of the feline visual
system. Deprivation and enrichment studies have been carried out
post-natal development in rodents are numerous and tend to confirm the uniquely high degree of plasticity of the system (e.g. Wilson,
Sullivan & Leon, 1987; Royet, Jourdan & Ploye, 1989a; Royet, Jourdan, Ploye & Souchier, 1989b). Indeed, Costanzo & Graziadei (1986) go so far as to state:
"The capacity for neurogenesis and reconnection of pathways in the olfactory system is unique in the mammalian nervous
system" (page 146).
1.9.10. It is tempting to generalise such studies to the human, but there are several problems, as outlined in the introduction to this
chapter. The first of these is that the necessary morphometric,
anatomical and ultrastructural data on the developing infant brain as a whole is lacking, though many general principles of neurogenesis have been propounded (Nowakowski, 1987). This view is taken by
Greenough and his colleagues (Greenough, Black & Wallace, 1987):
"Although research has demonstrated substantial effects of experience on brain connections, we do not yet understand just how the infant's brain is specialised to organise and incorporate experience, or the ways in which an infant may program its own experience" (page 539).
This is particularly so in the case of the human olfactory system. As explained elsewhere, much is known about this area from animal work. However, it is unlikely that the necessary in vivo human studies will be done to elucidate the olfactory system in infants or
adults. Because of this, plasticity in the postnatal human olfactory
system may only be inferred rather than demonstrated. However, as discussed in Chapter 3 of this thesis, there are reasonable grounds for assuming, and perhaps one day demonstrating, a high degree of plasticity in the human foetal olfactory system.
1.9.11. The second problem concerns what is actually meant by 'plasticity' and whether it is a unitary phenomenon at all in the
human infant. It has been suggested that there are at least two
One is called 'experience-expectant' plasticity, and relates to the organisation of the nervous system being set up to deal with sensory experiences common to all organisms. The other is called
'experience-dependent' plasticity, and appears to be the more applicable in the human case. This type assumes a high degree of flexibility in the nervous system, because certain sensory
experiences will tend to appear at unpredictable times during foetal
development. An example might be the arrival in the amniotic fluid
of a large quantity of a particular odorous molecule. Hence the developing nervous system has to be able to incorporate some
information at any time during maturation. Whether this is indeed
the case in the humans is speculative at the moment. There may even be more than two types of plasticity in a highly complex organism like the human foetus, though there is no empirical
evidence either way. Indeed, as Alberts (1981, cited in Schaal,
1988a) states:
"Little is known about the development of olfactory function as well as about the function of olfaction during development" (page 145).
However, in Chapter 3 of this thesis, a model is described which may go some way to answering this question.
1.9.12. To summarise this chapter, whilst much is known about the anatomy, physiology and ontogeny of the mammalian olfactory system, relatively little is known about these areas in the infant
human. This is probably a reflection of the concentrated interest in
other sensory modalities, which have received considerable attention (Werner & Lipsitt, 1981). Inevitably, olfaction and other sensory
systems have been relatively under-researched. This scientific
neglect has hitherto led to a biased view of the infant sensorium; with perhaps an inappropriate weight given to the importance of
vIsIon. This bias towards other sensory modalities continues to the
present day. As an example of this, in a recent review article
"the omission of taste, touch and olfaction... is ... an accurate reflection of the disproportionate concern which has been afforded to vision and audition in studies of infancy.... Such an imbalance would detract from a comprehension of fUnctioning at any stages of development, but may be particularly
distorting with regard to our understanding of the world of the infant" Ttirkewitz (1979, cited in Schaal, 1988a).
However, with increasing interest in the pre-natal infant world, this situation may improve. If, as argued by Schaal (1988a&b) and others, chemosensory experience and learning might begin in utero, then it is to be hoped that greater research resources will be directed to the study of infant olfaction.
1.9.13. It is important to understand this area in order to avoid the sort of distorted picture Ttirkewitz described. Although it is clear that older infants are visually dominant, this may not be the case for the immediate post-natal period. It seems possible that, in the first few weeks, or even months after birth, the infant requires a communications channel to the mother. This is, of course,
non-linguistic. In the period before the visual system has developed suffiCiently to permit high-capacity information input to the infant, the olfactory system may function as such a communications channel. The relative post-natal precociousness and probable high degree of plasticity in this system may allow some level of information
CHAPTER 2
A REYIEW OF THE LITERATURE IN THE FIELD OF 'INFANT OLFACTION'
Iptroduction
2.1.0. The aim of this chapter is to describe and evaluate the previous scientific research in infant olfaction. The purpose of this is twofold. Firstly, to chart progress in the field and identify key
contributors to the field. The second purpose is to provide a context
for the studies described in Chapter 4. The broad strands of the research literature will be examined, as far as possible in
chronological or related order and an attempt made to integrate the
findings. The pertinent literature is broadly summarised in Table 2.
The aim of this Table is to suggest groupings whereby the inter-relationships of the literature body can be made clearer and
therefore this chapter will have sub-headings which refer to Table 2. It should be emphasised that the groupings are not mutually
exclusive and that, for reasons of clarity, not all the studies discussed in this chapter are included in the Table. This especially refers to clinical papers.
2.1.1. The main conclusions of this chapter are that definite groupings in the literature are identifiable, despite the relatively
small number of papers in this area. The relatively meagre size of
the literature corpus in human infant olfaction, compared with the