With 1 plate and 16 text-figures Printed in Great Britain
REFLEX ORGANIZATION IN THE SWIMMERET
SYSTEM OF THE LOBSTER
I. INTRASEGMENTAL REFLEXES
BY W. J. DAVIS
Division of Natural Sciences, University of California, Santa Cruz, California g$o6o,U.S.A.
(Received 18 February 1969)
INTRODUCTION
The four pairs of abdominal swimmerets of the lobster perform rhythmic, meta-chronous beating movements which are used in larval and adult locomotion, righting responses, reproductive and perhaps respiratory behaviour (Davis, 1968a). Each swimmeret undergoes an alternating powerstroke (retraction) and returnstroke (pro-traction). During the powerstroke the two rami of the swimmeret spring open, increasing the exposed surface area of the appendage, and curl toward the rear, adding to the effectiveness of the powerstroke and positioning the rami for the return-stroke. During the returnstroke, the rami close and uncurl in preparation for the powerstroke of the next movement cycle (Davis, 1968a).
The motor output patterns which underlie these movements consist essentially of alternating bursts of impulses in two sets of antagonistic motor neurones (Davis, 19686, 1969 a). This pattern of output occurs even in de-afferented preparations, showing that it does not depend upon timing cues provided by sensory feedback from the moving limb, but is instead endogenous to the CNS. Each swimmeret neverthe-less possesses elaborate receptor systems which are capable of providing sensory feed-back for the possible regulation of its movements. Indeed, elimination of these sensory systems disturbs the co-ordination of the swimmeret movements (Davis, 1968 c), in-dicating that the sensory feedback which they provide plays a significant role in shaping the motor output pattern.
The role of sensory feedback in regulating the movements of the swimmerets is the subject of the investigation reported in this series of papers. The present paper and the one which follows (Davis, 19696) describe the intrasegmental swimmeret reflexes and their functional role in swimmeret beating. Subsequent papers will deal with the intersegmental reflexes (Davis, 1970 a) and the interactions between reflexive and endogenous central mechanisms in the swimmeret system (Davis, 19706).
MATERIALS AND METHODS
Eastern lobsters (Homarus americanus) were used. The abdomen was detached and the main flexor and extensor musculature was removed by dissection from the dorsal side, thereby exposing the ventral nerve cord and segmental nerve roots. The left half
548 W. J. DAVIS
of the abdomen was secured in a chamber with the ventral surface down. The pre-paration was arranged so that the swimmerets were accessible from the side, and the unobstructed movement of each swimmeret in its natural arc was possible. The CNS was not damaged by the operation, and all sensory and motor connections between the abdominal ganglia and the swimmerets remained intact and fully functional.
During the experiments the preparations were submerged in oxygenated saline (Cole, 1941) which contained about 10 g./l. of granular dextrose. The saline was buffered with ' T E S ' (Sigma Chemical Co.) and maintained at io-i5°C. The reflex activity was especially sensitive to the pH of the saline, which was held at 7-2-7*4. With strict adherence to these procedures the preparations yielded useful data for several hours.
Glass pipette electrodes with fire-polished tips were attached by suction to various branches of the first abdominal nerve root to allow extracellular recording of efferent impulse traffic on its way to the swimmeret muscles. Swimmeret muscle potentials were recorded either extracellularly with suction electrodes on intracellularly with glass capillary micro-electrodes. Nerve and muscle potentials were amplified, dis-played on a Tektronix oscilloscope and filmed with a Grass C-4 camera.
Controlled movements of the swimmeret were accomplished by means of the device illustrated in Text-fig. 1. The swimmeret was coupled to one end of a lever whose movement was monitored with a photocell system and taken as an indication of the position of the swimmeret. Stepwise movements of the swimmeret were obtained by moving the lever by hand. Sinusoidal movements of the swimmeret were obtained by coupling the lever to an eccentric point on a rotating wheel, which was in turn coupled through a gear box to a variable speed electric motor.
RESULTS
Innervation of the swimmeret muscles
The swimmeret muscles, whose anatomy is described elsewhere (Davis, 19686), are supplied exclusively by the ipsilateral first nerve root of the ganglion of the cor-responding abdominal segment. The first root consists of two main bundles, anterior and posterior. At the base of the swimmeret the two bundles each divide into a dorsal and a ventral branch (Text-fig. 2). The anterior dorsal branch contains the efferent innervation of the main returnstroke (protractor) muscle (muscle 1-3; Davis, 1968 J), while the posterior dorsal branch supplies the main powerstroke (retractor) muscle (muscle 4-8; Davis, 19686). In the present experiments all of the nerve recordings were made from these two branches. The remaining muscles are contained within the swimmeret itself and are innervated mainly if not entirely by ventral branches of the posterior bundle of the first root.
Identification of efferent units
as well as motor axons. To ensure that the reflex discharge recorded from these sites was motor rather than sensory, the following criteria were used, either alone or in various combinations: (1) rhythmic, spontaneous bursts of impulse similar to the efferent patterns which underlie swimmeret beating were recorded from the neurones
Photocell
Text-fig. 1. Apparatus used for controlled movement of the swimmeret in its natural arc (see Materials and Methods). The swimmeret was coupled to one end of the moving lever. The posi-tion of the lever was monitored by passing light to a photocell through two pieces of polarized glass, one attached to the moving lever and one fixed in place over the photocell. The light trans-mitted to the photocell was directly proportional to the angle between the polarizing planes of the two pieces of glass, which was in turn proportional to the lever position.
in the absence of sensory feedback; (2) impulses in the neurones were correlated in a 1:1 fashion with potentials recorded from the swimmeret muscles; (3) the reflex responses of the neurones were abolished by severing the entire first root at its point of exit from the abdominal ganglion; and (4) stimulation of the sense organs which initiate the reflexes caused excitatory postsynaptic potentials and action potentials, both of which were recorded intracellularly from the somata of identified swimmeret motor neurones in the abdominal ganglia. The action potentials recorded from the
somata were correlated in a i : i fashion with reflex spikes recorded peripherally from the motor neurone. In addition to these tests, the major reflexes reported here have also been demonstrated by recordings made directly from the swimmeret muscles rather than from the motor neurones (e.g. Text-fig. 13).
Text-fig. 2. Innervation of the muscles of a single swimmeret on the left side of the abdomen. The view is from the midline, with dorsal on the top of the illustration and anterior on the right. Solitary numbers identify bundles of muscle fibres described elsewhere (Davis, 19686). Bundles 1-3 comprise the main retumstroke (protractor) muscle, while 4-8 make up the main powerstroke (retractor) muscle. 1 a and 16 designate the anterior and posterior bundles of the first nerve root, respectively, a.d., anterior dorsal branch; a.v., anterior ventral branch; p.d., posterior dorsal branch; p.v. 1 and p.v. 2, posterior ventral branches.
Impulses belonging to a single neurone were the same in amplitude and waveform. In these respects the variation between different motor neurones was usually large enough to allow unambiguous identification of single-unit activity.
Reflex responses to imposed retraction of the swimmeret
Experiments have been performed on the swimmerets of all segments of both sexes. The intrasegmental swimmeret reflexes were qualitatively identical in all segments, and no systematic differences were found between the strength of the reflexes in different segments.
Retumstroke {protractor) motor neurones. When a swimmeret was forcibly retracted
reflex response showed an initial high-frequency phasic component, but decayed within several seconds to a steady, tonic discharge which was maintained for as long as the swimmeret was retracted (Text-fig. 4).
5 sec
I
Text-fig. 3. Recording from the anterior dorsal branch of the first nerve root (upper trace), which supplies the main returnstroke muscle (Text-fig, a), during stepwise retraction of the swimmeret (upward deflexion of the lower trace), (a) and (6) are the beginning and end of the same record. Several seconds of activity between (a) and (6) are omitted.
20 18 16 14 12 10 8 6 4 2
3
i i i
0 4 8 12 16 20 24 28 32 36 40 44 48 52
Time (sec.)
Text-fig. 4. Time course of the response of two excitor neurones in the main retumstroke nerve during stepwise retraction of the swimmeret. The stimulus (lowest curve) and the responses (upper two curves) are plotted on the same time base. The record analysed here is shown in Text-fig. 3. The response of both neurones shows the typical initial phasic and long-term tonic components.
reduced (Text-fig. 5). As expected, the phase histogram of the response of this neurone to sinusoidal movement of the swimmeret was usually shifted approximately 1800 from that of the excitor neurones, i.e. its response approximately alternated with that of the excitors (e.g. Text-fig. 6). A similar reciprocity of activities occurred even in the absence of reflexive inputs, as shown by the following observations.
n=64,
low=1900°, r=+0-8465
n=166, low=67-30°,
+0-7263
300 360
Time (sec.) Fig. 5
Phase position of spikes in the imposed sinusoidal movement cycle (deg.)
Fig. 6
Text-fig. 5. Reflex response of three neurones in the main returnstroke nerve (upper three curves) during stepwise retraction of the swimmeret (lowest curve). The uppermost curve shows the response of the inhibitor axon, while the middle two curves show the typical responses of excitor neurones. The data were taken from the same preparation as illustrated in Text-fig. 7. Text-fig. 6. Phase histograms showing the positions of efferent returnstroke nerve spikes in the input cycle during sinusoidal movement of the swimmeret for many consecutive cycles of movement. o°-i8o° corresponds to protraction of the swimmeret, i8o°-36o° to retraction, (a), response of the peripheral inhibitor axon; (6), response of the two excitor axons. The data were taken from the same preparation as shown in Text-fig. 7. n = the number of observations, low = the phase position of most intense activity in the movement cycle, and r = the correla-tion coefficient which expresses the strength of impulse grouping in the input cycle (see Davis, 19696).
muscles deteriorated, and in all cases the swimmeret was immobilized. Thus, the pro-duction of the patterns was independent of the occurrence of sensory feedback, and undoubtedly reflected the activity of an endogenous oscillator in the CNS. During the production of such patterns, the same returnstroke neurone which showed reflex reciprocity with the excitors was active in bursts which alternated with bursts in the returnstroke excitor neurones (Text-fig. 7). Thus, the reciprocity between these units is also programmed into the central nervous motor score.
I 500 msec I
II.
I
Li.
U
Text-fig. 7. Recordings made from the main rerumstroke nerve of one preparation during sinu-soidal movement of the swimmeret (a) and during the spontaneous production of rhythmic motor patterns similar to those which underlie swimmeret beating (6). The tension response of the muscles had deteriorated and the swimmeret was fixed in the retracted position in (6); thus, the rhythmic motor discharge patterns were produced without the aid of sensory feed-back. Arrows point to impulses in the peripheral inhibitor axon. The records demonstrate reciprocity between the activities of excitor and peripheral inhibitor axons, during reflex (a) and purely central (6) driving of motor activity.
What is the function of the neurone which shows reciprocity with the excitors? The reciprocity alone suggests that the axon is a peripheral inhibitor. This interpreta-tion is corroborated by experiments on the swimmeret system of the crayfish, where recordings from the main returnstroke nerve also show the alternation of simultaneous bursts in several motor neurones with a burst of impulses in a single neurone. The former are correlated with depolarizing potentials in the main returnstroke muscle, while the latter are associated in a i : i fashion with hyperpolarizing potentials (P. Stein, unpublished data).
from active inhibition caused by sensory receptors responsive to retraction, or from the withdrawal of excitation from possible sensory receptors capable of signalling limb protraction. Additional experiments are needed to decide between these various interpretations.
I 1 sec
<•>
(b)
Text-fig. 8. Recordings from the posterior dorsal branch of the first nerve root (upper trace), •which supplies the main powerstroke muscle (Text-fig. 2), during stepwise retraction of the swimmeret (upward deflexion of the lower trace), (a) and (6) are the beginning and end of the same record. Several seconds of activity between (a) and (b) are omitted.
12 10 8 6 4 2 0 6 4 2 0 -o -opc
J
I I QV
V
pPOT o, p po W o l\m<h o\/b
w U J L ^ I UAJU Qy y H
Co 0 o C
I
Ytam
Retraction
1 i i i i i i i
p
\ 0 O o
Q Q 0 O
tocooWb
\ Protraction
I I I I
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 Time (sec.)
Text-fig. 9. Time course of the responses of two excitor neurones in the main powerstroke nerve during stepwise retraction of the swimmeret. The stimulus (lowest curve) and the responses (upper two curves) are plotted on the same time base. The record analysed here is shown in Text-fig. 8. The responses of both neurones show the typical initial phasic and long-tonic components.
Powerstroke (retractor) motor neurones. When the swimmeret was forcibly retracted,
tonic discharge. The tonic response persisted for as long as the swimmeret was held in the retracted position (Text-figs. 8, 9). The reflex action of the powerstroke excitor neurones therefore further reinforces the movement which initiates it. Unlike the resistance reflex of the returnstroke neurones, the feedback to the powerstroke muscle is positive with respect to the movement.
1iec
\ \ \ \ \
urn
Text-fig. 10. Reciprocity between the activities of excitor and peripheral inhibitor axons in the main powerstroke nerve during reflexive inputs, (a) Stepwise retraction of the swimmeret (upward deflexion of the lower trace). (6) Sinusoidal movement of the gwimmeret. Impulses in one neurone are associated with small, possibly hyperpolarizing potentials (arrows) from a muscle fibre which was sucked into the recording electrode. The polarity of these potentials may indicate that the axon which causes them is a peripheral inhibitor.
As in the case of the returnstroke response, multi-unit recordings from the power-stroke nerve often contained a single unit whose response to retraction of the swim-meret was opposite from that of the excitor neurones (Text-figs. 10, n ) . Impulses in this neurone were sometimes associated with small muscle potentials which were opposite in polarity from the muscle potentials affiliated with excitor neurone impulses. All of the available evidence thus suggests that this axon was a peripheral inhibitor, which also responds to reflexive inputs reciprocally with the excitor neurones supplying the same muscle.
Sensory basis of the reflex responses to retraction of the swimmeret
556
Photomicrographs of sections of the coxal region show that each strand possesses an apparent nerve supply, presumably sensory in function (PL i). The detailed structure and function of these organs has not been studied carefully, but the observa-tions described here suggest that they are proprioceptors analogous to those found in other crustacean appendages (e.g. Alexandrowicz & Whitear, 1957; Bush, 1964; Burke, 1954; Cohen, 1963; Pilgrim, i960; Taylor, 1967; Wiersma, 1959; Wyse & Maynard, 1965). These proprioceptors probably provide the sensory inputs for the retraction reflexes described above, although other sensory receptors, such as cuticular receptor cells in the membrane which articulates the swimmeret with the abdomen (Pabst & Kennedy, 1967), may also participate.
n=71,
I3\ low=178-2°, w
r=+0-7673
1 0 8 6 4 2
-a 0
g 60 120 180 240 300 360
n=132, low=37-9°, /•=+0-1977
60 120 180 240 300 360
Phase position of spikes in the imposed sinusoidal movement cycle (deg.)
Text-fig. 11. Phase histograms showing the positions of efferent powerstroke nerve spikes in the input cycle during sinusoidal movement of the swimmeret for many consecutive cycles of movement. The histograms are similar to those described in Text-fig 6. (a) The response of the peripheral inhibitor axon; (6) the response of excitor neurones. In (6) the correlation is weaker than usual for the excitor response, but nevertheless significant at the 5 % level. The data were taken from the same recording as shown in Text-fig. 10.
Reflex responses to stimulation of the sensory setae on the rami
The edges of the two rami of each swimmeret are fringed with filamentous setae which represent a major source of sensory feedback for the swimmeret reflexes. These reflexes can be elicited by displacing the setae with jets of water. Streams directed from the front have little or no effect, but streams directed from the rear consistently elicit the reflexes. The setae therefore appear to be stimulated normally by the flow of water against'them during the powerstroke.
before their entry into the basipodite of the swimmeret. Most of these axons are simultaneously excited by gently squeezing the rami distal to this point of convergence. This method of stimulation does not permit fine control of the stimulus parameters, but quantitative regulation of the stimulus was not required to demonstrate the reflexes.
Powerstroke {retractor) motor neurones. Activation of the sensory axons from the
setae by gently squeezing the rami with forceps elicited an intense burst of impulses in the excitor neurones which supply the main powerstroke muscle (Text-fig. 12). In contrast, the peripheral inhibitor axon to the powerstroke muscle responded to the same stimulus with a decrease in its discharge rate (Text-fig. 12). This reflex may also be considered a resistance reflex, since its action opposes the forward displace-ment of the swimmeret by water currents directed against the rami from the rear. During swimmeret beating, however, this reflex appears to provide positive feedback to the powerstroke muscle during the powerstroke, unlike the resistance reflex of the return-stroke excitor neurones in response to retraction.
I
1 secI
•»—f
i l l
1 ) 1
i\i Jill
'11
1
!
1
Text-fig. 12. Recordings from the main powerstroke nerve during gentle, rhythmic squeezing of the rami with forceps (large arrows). This stimulus excites the afferent pathways from the sensory setae which border the rami of the swimmeret (see text). In (a) the smallest unit responds oppositely from the large excitors. The same is true in (6) where the small nerve impulses are associated with muscle potentials which are presumably hyperpolarizing. These potentials suggest that the neurone is an inhibitor. This figure demonstrates the activation of powerstroke excitor axons by stimulation of the setae on the rami, and the reciprocity between excitor and inhibitor axon activity for this source of sensory input. The record in (6) was taken from the same preparation as those in Text-fig. 10.
Returnstroke {protractor) motor neurones. Stimulation of the afferent pathways from
Squeezing the rami vigorously often abolished even the most intense discharge in the retumstroke excitor neurones for several seconds. The duration of the inhibitory effect was considerably greater than the duration of the corresponding excitatory effect on the powerstroke excitor neurones (cf. Text-fig. 14 with Text-fig. 12).
There-I 5 sec. |
Text-fig. 13. Muscle potentials recorded from the main retumstroke muscle (muscle 1-3; Davis, 19686) with a suction electrode (upper trace) during retraction of the swimmeret (upward deflexion of the lower trace). While the gwimmeret was retracted the two rami were gently squeezed with forceps (upward arrow) and released (downward arrow). This stimulation of the sensory axons from the setae reduced the frequency of excitatory discharge to about one-quarter its original level.
22 20 18 16 •a 14
I 12
•S 10 & 8z
6
8. 4
I
2
ft 0
Rami squeezed
Retraction
, Protraction
j I
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28
T i m e (sec.)
Text-fig. 14. Time course of the response of the main retumstroke excitor neurones (upper curve) to retraction of the swimmeret (lowest curve) and simultaneous gentle squeezing of the rami (middle curve). The experiment was similar to that shown in Text-fig. 13, but was performed on a different preparation. The inhibiting effect of the stimulus persisted for several seconds longer than the stimulus duration.
excitors, i.e. squeezing the rami increased its activity. This unit may have been the peripheral inhibitor axon which supplies the muscle, but independent evidence of its inhibitory function was not obtained.
Distribution of the reflexes to the remaining swimmeret muscles
Systematic recording from motor neurones other than those which innervate the main powerstroke and returnstroke muscles has not been attempted during activation of the swimmeret reflexes. Forcibly retracting the swimmeret often caused the rami to spring open and curl toward the rear, however, as they do during a voluntary power-stroke (see Introduction). If the tension response of the muscles had not deteriorated, the opening and curling movements elicited by imposed retraction appeared as vigorous as the same movements during swimmeret beating. These results suggest the possibility that the contraction of the corresponding muscles in the normal movement cycle may be controlled largely by reflexes activated during the powerstroke.
DISCUSSION
Reflex compensation for central excitability gradients
In an earlier study of the swimmeret movements it was found that the power pro-duced by individual swimmerets is the same despite a probable rear-to-front decrease in the central 'excitability' of the segmental swimmeret oscillators. This result led to the hypothesis of a compensating reflexive amplification of the power output from the rear to the front of the abdomen (Davis, 1968 c). In the present work no systematic differences have been found between the strength of the intrasegmental swimmeret reflexes in different abdominal segments. A rigorous comparison of reflex strength of course requires good control over the strength of the stimulus used to elicit the reflexes. Although such control was lacking in the present experiments, the results nevertheless suggest that a segmental gradient in the strength of the intrasegmental reflexes can be reasonably excluded as the hypothetical rear-to-front amplifying mechanism. In the third paper of the present series, however, it will be shown that the intersegmental reflexes are capable of providing the proposed rear-to-front amplifica-tion (Davis, 1970 a).
Reflex activity during swimmeret beating
score (Davis, 1970&). The reflexive inputs to the main powerstroke and retumstroke muscles are organized to effect reciprocity of peripheral inhibitor axon activity with excitor neurone activity to the same muscle.
The above interpretation naturally requires the assumption that the reflexes can
Sensory setae on rami
Coxal proprioceptors
i h-Ti
Powentroke muscle w 0 . O -C c0 J
e 0 ci t Xr
Returnstroke muscle0 0 o
Swlmmeret motor •t; neurones Powerstroke muscle u _o .c c o *^ u X U i
r
Returnstroke muscle 2 Swimmeret motor neuronesFig. 15 Fig. 16
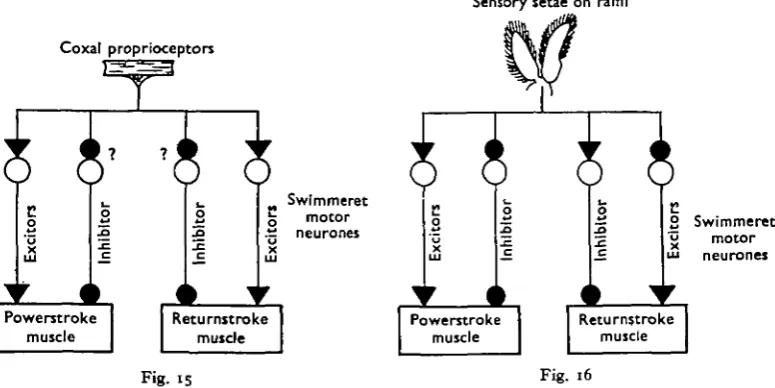
Text-fig. 15. Diagram symbolizing the reflex pathways activated by retraction of the swimmeret and probably controlled by the coxal proprioceptors. Black arrowheads denote excitatory influences, while black circles denote inhibitory ones. The major features illustrated are (i) the similarity of the effect of proprioceptive input on powerstroke and returnstroke motor neurones, and (2) the reciprocal effect of this input on excitor and peripheral inhibitor axons to the same muscle. The pathways designated by question marks could be replaced by different ones which are equally capable of accounting for the available data (see text).
Text-fig. 16. Diagram symbolizing the reflex pathways activated by stimulation of the sensory setae which border the rami of the swimmeret. Black arrowheads represent excitatory in-fluences, while black circles represent inhibitory ones. The major features illustrated are (i) the opposite effects of setae stimulation on the powerstroke and returnstroke motor neurones, and (a) the reciprocal effect of setae stimulation on excitor and peripheral inhibitor axons to the game muscle.
follow movements as rapid as those which occur normally during swimmeret beating. This is confirmed in the following paper (Davis, 1969 b). From the above hypothetical reconstruction of events it is evident that reflex initiation of the returnstroke by the powerstroke movement is possible, but a converse initiation of the powerstroke by the returnstroke does not occur. In fact, no intrasegmental reflexes have been found which are capable of initiating the powerstroke. This point assumes special signifi-cance in the context of the discussion in the next section.
Reflex contribution to the periodicity in the normal motor output
[image:14.451.34.422.133.327.2]As emphasized above, no intrasegmental reflex has been found which is capable of initiating the power stroke. Anterior-going intersegmental reflexes linking the power-strokes of different swimmerets have been identified (Davis, 1970 a), but these reflexes are incapable of initiating the powerstroke of the rear-most swimmeret. Moreover, a swimmeret performs normal movements even when input from more posterior seg-ments is blocked. Therefore it appears that in the intact lobster the powerstroke of each movement cycle must be initiated anew by a purely central nervous motor com-mand. Furthermore, all of the identified reflexive feedback to the powerstroke ex-citor neurones is positive, further reinforcing the powerstroke movement. Therefore it appears that in the intact lobster the cessation of the powerstroke discharge of each movement cycle is also controlled exclusively by the central motor score. The present results and the earlier work of others on the crayfish swimmeret system (Hughes & Wiersma, i960; Ikeda & Wiersma, 1964; Wiersma & Ikeda, 1964) have shown that the CNS can generate the complex, oscillatory pattern of motor output which underlies swimmeret beating, even in the absence of rhythmic timing cues from sensory feed-back. The present work has shown further that even when this sensory feedback is operative, the reflexes which it controls are in fact unable to directly initiate and/or sustain the cyclic output pattern.
Reflex amplification of the central motor command
The present study has shown that the intrasegmental swimmeret reflexes are organized so that they strengthen the powerstroke, and also reinforce the linkage between the powerstroke and the returnstroke within each cycle of movement. The reflexive inputs may also contribute to the reciprocity between excitor and peripheral inhibitor axon activities. The present work has shown, however, that each of these features of the motor output pattern is programmed into the central motor score independently of sensory feedback. Thus, the influence of the intrasegmental reflexes on the motor output pattern is quantitative rather than qualitative. In the event of an experimentally-induced conflict between the reflexive and endogenous central nervous mechanisms, the central command strongly dominates (Davis, 19706). The intra-segmental reflexes may therefore be viewed as relatively subservient amplifying mechanisms for independent neuronal oscillators in the CNS.
corresponding resistance to the movement. Such a mechanism for automatically adjusting the motor output to handle variable external loads may be present in the swimmeret system, but if so, the method of load compensation is clearly based upon different principles than the follow-up length servo of the vertebrate myotatic reflex.
SUMMARY
1. The intrasegmental feedback reflexes in the swimmeret system of the Iob8ter
Homarus americanus were activated while recording the responses from the swimmeret
nerves and muscles.
2. Two rnqin sources of sensory feedback were identified; proprioceptors in the coxal region of the swimmeret, and sensory setae on the edges of the two rami of each swimmeret. The reflexes activated by these inputs are described.
3. Reflexive feedback from the powerstroke movement to the powerstroke excita-tory motor neurones is positive, further reinforcing the movement. Intrasegmental reflexes capable of independently initiating or terminating the powerstroke activity are absent, however. Therefore the powerstroke movement of each cycle can begin and end only in response to a purely central nervous motor command. It follows that the intrasegmental swimmeret reflexes are incapable of contributing to the periodicity seen in the motor output pattern which underlies swimmeret beating.
4. In addition to strengthening the powerstroke, the intrasegmental reflexes strengthen the linkage between the powerstroke and the returnstroke within each movement cycle. The reflexes may also reinforce the reciprocity between excitor and inhibitor axon activity to the main powerstroke and returnstroke muscles.
5. It is shown, however, that these three features of the motor output pattern are programmed into the CNS independently of the sensory feedback. The intraseg-mental reflexes thus act as subservient amplifying devices for cyclic motor patterns which are produced independently by purely central nervous mechanisms.
This work was supported by NIH postdoctoral fellowship NB 24, 882 to the author and by NIH grant 5 Roi NB 01624 to Dr Melvin J. Cohen. I am grateful to Dr D. Kennedy, Dr D. M. Wilson, Dr P. Stein, Dr A. Selverston and Dr D. Hartline for discussion and criticism of the manuscript.
REFERENCES
AIEXAKDROWICZ, J. S. & WHITEAR, M. (1957). Receptor elements in the coxal region of Decapoda
Crustacea. J. mar. biol. Ass. UJC. 36, 603-28.
BURKE, W. (1954). An organ for proprioception and vibration sense in Cardnus maenas. J. exp. Biol. 31, 127-38.
BUSH, B. M. H. (1964). Proprioception by chordotonal organs in mero-carpopodite and carpo-propodite joints of Cardnus maemts legs. Comp. Biochem. Physiol. 14, 185-99.
BUSH, B. M. H. (1965). Leg reflexes from chordotonal organs in the crab, Cardnus maenas. Comp.
Biochem. Physiol. 15, 567-87.
BUSH, B. M. H. & ROBERTS, A. (1968). Resistance reflexes from a crab muscle receptor without impulses. Nature, Land. ai8, 1171-3.
COHEN, M. J. (1963). The crustacean myochordotonal organ as a proprioceptive system. Comp. Biochem.
Physiol. 8, 223-43.
COLE, W. H. (1941). A perfusing solution for the lobster {Homarus) and the effects of its constituent ions on the heart. J. gen. Physiol. 25, 1-6.
Journal of Experimental Biology, Vol. 51, No. 3
Plate 1
150 ,i
DAVIS, W. J. (19686). The neuromuscular basis of lobster swimmeret beating. J. exp. Zool. 168, 363-78. DAVIS, W. J. (1968c). Quantitative analysis of swimmeret beating in the lobster. J. exp. Biol. 48,
643-62.
DAVIS, W. J. (1969a). The neural control of swimmeret beating in the lobster. J. exp. Biol. 50, 99-117. DAVIS, W. J. (19696). Reflex organization in the swimmeret system of the lobster: II. Reflex dynamics,
J. exp. Biol. 51, 565-73.
DAVIS, W. J. (1970a). Reflex organization in the swimmeret system of the lobster: III. Intersegmental reflexes. (In preparation.)
DAVIS, W. J. (19706). Interaction between reflex and endogenous central nervous mechanisms in the swimmeret system of the lobster. (In preparation.)
FIELDS, H. L. (1966). Proprioceptive control of posture in the crayfish abdomen. J. exp. Biol. 44, 455-68.
GRANIT, R. (1966). Muscular Afferent! and Motor Control. Stockholm: Almqvist and Wiksell., HUGHES, G. M. & WIERSMA, C. A. G. (i960). The co-ordination of swimmeret movements in the
crayfish Procambarus clarfdi (Girard). J. exp. Biol. 37, 657-70.
IKEDA, K. & WIERSMA, C. A. G. (1964). Autogenic rhythmicity in the abdominal ganglia of the crayfish: The control of swimmeret movements. Comp. Biochem. Pkytiol. ia, 107—15.
PABST, H. & KENNEDY, D. (1967). Cutaneous mechanoreceptors influencing motor output in the cray-fish abdomen. Z. vergl. Physiol. 57, 100-208.
PILGRIM, R. L. C. (i960). Muscle receptor organs in some Decapod Crustacea. Comp. Biochem. Pkytiol. 1, 248-57.
TAYLOR, R. C. (1967). The anatomy and adequate stimulation of a chordotonal organ in the antennae of a hermit crab. Comp. Biochem. Physiol. 30, 709-18.
WIERSMA, C. A. G. (1959). Movement receptors in Decapod Crustacea. J. mar. biol. Ass. U.K. 38,
143-52-WIERSMA, C. A. G. & IKEDA, K. (1964). Intemeurons commanding swimmeret movements in the cray-fish, Procambarus clarkii (Girard). Comp. Biochem. Physiol. 13, 509-25.
WYSE, G. & MAYNARD, D. M. (1965). Joint receptors in the antennule of Panulirus argus Latreille.
J. exp. Biol. 43,
521-535-EXPLANATION OF PLATE
(a) Section through the coxal region of the swimmeret, showing the elastic strand A («.t.) at its point
of insertion on the tendons (f.) at the base of the main returnstroke muscle. *.»., probable sensory inner-vation which terminates on the strand as a raised hillock (h.). The section was made parallel to the frontal plane of the lobster, b, c and d, sections parallel to the one in a, made through the coxal region of the swimmeret to show the elastic strand designated B (e.s.). The sections in b and d were made respectively at the mid-point of the strand and at its point of insertion adjacent to muscle 9 (6. 9; Davis, 19686). The section in c was taken from a point between those in b and d. In c, 1.1. designates the probable sensory innervation of the strand, while c.b. denotes possible cell bodies which resemble those found embedded in the connective tissue strands of many crustacean proprioceptors (e.g. Cohen, 1963).