2(3): 292-312, 2012
SCIENCEDOMAIN international www.sciencedomain.org
Chronic Fluoxetine Administration during
Different Postnatal Development Stages Leads
to Stage Dependent Changes of Glial Fibrillary
Acidic Protein Expression in Rat Brain
Nathalie Bock
1*, Hannah Alter
1, Emre Koc
1, Veit Roessner
2,
Aribert Rothenberger
1and Till Manzke
3,41
Department of Child and Adolescent Psychiatry, Georg-August-University of Göttingen, von Siebold-Strasse 5, 37075 Göttingen, Germany.
2
Department of Child and Adolescent Psychiatry, Technische Universität Dresden, Dresden, Germany.
3
Department of Neuro- and Sensory Physiology, Georg-August-University of Göttingen, Humboldtallee 23, 37073 Göttingen, Germany.
4
DFG Research Center Molecular Physiology of the Brain, Göttingen, Germany.
Authors’ Contributions:
Nathalie Bock designed the study, performed the statistical analysis, wrote the protocol, and wrote the first draft of the manuscript. Hannah Alter and Emre Koc managed the analyses of the study. Veit Roessner managed the literature searches. Aribert Rothenberger was involved in writing of the manuscript. Till Manzke designed the study and wrote the protocol. All authors read and approved the final manuscript.
Received 26th January 2012 Accepted 22nd March 2012 Online Ready 14th April 2012
ABSTRACT
Aims: Depressive symptoms in children and adolescents are commonly treated with serotonin re-uptake inhibitors like fluoxetine. Astrocytes expressing different serotonin receptors (5-HTRs) and the serotonin transporter (5-HTT) are affected by fluoxetine administration. The study was conducted to revise whether fluoxetine treatment during postnatal brain development results in long-term changes of astroglia.
Methodology: Thus, immunohistochemistry and real-time PCR analyses were performed at different postnatal periods in rats to investigate short- and long-term changes following by a 14-day administration with fluoxetine (5 mg/kg/BW s.c. once daily).
Results: Fluoxetine-treatments from postnatal day (pd) 1-15, measured at pd 16, led to a significant reduction of GFAP gene (Gfap) in hippocampus and GFAP protein expression in the dentate gyrus and CA1 without changes at pd 90 compared to controls. Treatments from pd 21-35 resulted in a significant decrease of Gfap and protein (measured 24 hours after last injection) in striatum (putamen), frontal cortex and hippocampus. Contrary, if measured at day 90, the same treatment led to a significant increase in those regions. Later treatments from pd 50-64 did not result in significant changes in mRNA or in protein expression.
Conclusion: This study revealed a fluoxetine-sensitive period of brain astrocyte development (i.e. periadolescence) that led to structural effects, which can even be detected in adulthood. These results might be relevant for psychopharmacological treatment in children and require continuative clinical studies.
Keywords: Astroglia; fluoxetine; brain development; GFAP/Gfap.
1. INTRODUCTION
Although psychopharmacological treatment of depression in children and adolescent is strongly increased during the past, information about the influence of neuropharmacological drugs (i.e. antidepressants) on postnatal human brain development is rare. This is even though many studies already revealed neuronal changes induced by fluoxetine and other antidepressants in human adults and adult animals (Mongeau et al., 1997; Blier and de Montigny, 1999; Duman et al., 2001). Therefore, it is of great interest to understand whether antidepressants during postnatal development could lead to distinct changes or whether the higher plasticity in the juvenile brain has more possibilities to compensate induced changes in the long-term.
Besides neuronal alterations, changes in number and size of glia cells could be also of interest, since these are affected in different brain regions of humans with major depression. For example, reduction in the number of glia cells and in their packing density were reported in postmortem brains from subjects with mood disorders in different regions of the prefrontal cortex (PFC), the anterior cingulate cortex, and hippocampal subfields (Rajkowska et al., 1999; Rajkowska, 2000; Cotter et al., 2001; Stockmeier et al., 2004). In addition, investigations concerning GFAP, an astroglia cell marker, revealed reduced GFAP levels in patients with major depression (Webster et al., 2001; Fatemi et al., 2004; Webster et al., 2005).
Nevertheless, a causative influence of astrocytes on depression was described by Banasr and Duman, 2008. They observed depressive-like behavior after application of an astrocyte selective toxin into the rat prefrontal cortex.
In the traditional concept, the function of astrocytes was restricted to regulate and optimize the environment for neuronal function. From more recent studies it is, however, known that astrocytes also synthesize and release different neurotrophic factors such as brain-derived neurotrophic factor (BDNF), glial-derived neurotrophic factor (GDNF), nerve growth factor (NGF), and neurotrophins 3 and 4/5, which are vital for neuronal health (Friedman et al., 1998; Althaus and Richter-Landsberg, 2000). Furthermore, one of their principal functions is the uptake of neurotransmitters released from nerve terminals (Hansson and Rönnbäck, 1992; Zhang and Barres, 2010) They can also release neuroactive agents, including transmitters, eicosanoids, steroids, neuropeptides, and growth factors (Anderson and Swanson, 2000).
Fluoxetine exerts a direct effect on astrocytes, since they express several 5-HT receptor subtypes, for example, 5-HT1A, 5-HT2B and 5-HT2C receptors. If the mechanism of action of fluoxetine shall be explained, the latter mechanism cannot be ignored (Chen et al., 1995; Azmitia et al., 1996; Azmitia, 2001; Kong et al., 2002). Especially 5-HT1A receptors are abundantly expressed on hippocampal astroglia (Azmitia et al., 1996). Their activation is suggested to be a critical component in the mechanism of action of SSRIs (Santarelli et al., 2003). Moreover, in mammals astrocytes can take up serotonin by a sodium-dependent high affinity system, and the presence of MAO-A in brain astrocytes has been established by immunohistochemistry (Kimelberg and Katz, 1985; Fitzgerald et al., 1990).
Several studies (Wegerer et al., 1999; Moll et al., 2001; Bock et al., 2005; Roessner et al., 2010) revealed long lasting changes following psychopharmacological treatment depends on the time frame of drug administration during postnatal development. It is known that astrocytes undergo a certain two-step developmental expression. At the RNA level GFAP increases between birth and day 15, which is described as the period of astrocytic proliferation and then decreases until day 55 (period of astrocytic morphological differentiation) (Tardy et al., 1989). This findings are in line with developmental changes of the number of GFAP-positive astrocytes in the hippocampal formation described by (Kimoto et al., 2009).
Therefore, we designed this study to investigate the possible influence of chronic fluoxetine administration on postnatal development of astrocytes. Messenger RNA and protein expression of a common marker for astrocytes, the glial fibrillary acidic protein GFAP, were studied. GFAP is enriched in the cell body and the major processes of astrocytes (Bushong et al., 2002; Ogata and Kosaka, 2002) in the frontal cortex, hippocampus or striatum. These regions, which all receive strong serotonergic projections, were selected according to their physiological and clinical relevance (Lidov and Molliver, 1982; Rubenstein, 1998; Hensler, 2006).
2. MATERIALS AND METHODS
Sprague Dawley rats were obtained from a commercial breeder (Charles River, Sulzfeld, Germany) and used for further breeding in our own environmentally conditioned animal facility under standardized conditions. After mating, the dams were housed individually in cages with free access to food (ssniff R-M-H, 10 mm, Spezialdiäten GmbH, Soest, Germany) and water. Litter size was reduced to six male pups per dam. First group of rats (n = 24) received fluoxetine s.c. from day 1-15, control animals received saline. After weaning (day 21) the young rats were placed in separate cages, two per cage. The second group (n = 24) received fluoxetine s.c. from day 21-35 and a third group (n = 24) from day 50-64. In the control group, the same number of animals received saline instead of fluoxetine. The drugs were administered in the morning (always at the same time) via subcutaneous injection. The dose of fluoxetine was adjusted to 5 mg/kg body weight per day according to the suggestions of Wegerer et al., 1999. There was no evidence that fluoxetine administration affected body weight gain in the rats. Twelve animals per group were dispatched 24 hours after the last injection of fluoxetine or saline, and the remaining 12 animals per group at day 90. After 24 hours fluoxetine and its major active metabolite norfluoxetine are still in the brain in contrast to the later analyzed groups. Six animals each were used for immunohistochemical studies and 6 animals for real-time PCR analysis.
2.1 Real-time PCR
For real-time PCR analysis Sprague-Dawley rats were sacrificed by decapitation. After removal of the brain, tissue was directly frozen on dry-ice and then frozen at -80°C. The hippocampus, striatum (putamen), and the frontal cortex were cut from corresponding cryostat sections (n = 6 for each group; postnatal day 16 to 90). The total ribonucleic acid (RNA) of the homogenized brain tissue was isolated by using the Trizol® method according to manufacturer’s instructions (GibcoBRL). Total RNA concentration was determined by using the nanodrop ND-1000 spectrophotometer followed by its quality and integrity measurement by electrophoresis on RNA 6000 LabChip® kit (Agilent 2100 Bioanalyzer). The RNA was transcribed into the corresponding deoxyribonucleic acid (cDNA) by using the iScript cDNA Synthesis Kit (BioRad).
The following primer pairs were designed by using the Primer3 program (http://frodo.wi.mit.edu/cgi-bin/primer3/primer3_www.cgi): hypoxanthine guanine phosphoribosyl transferase (Hprt,135 bp; NCBI-Accession No.: NM_012583), F (5'-gtcaagcagtacagccccaaaatg-3´) / R (5'-gtcaagggcatatccaacaacaaac-3´) and glial fibrillary acidic protein (Gfap, 247 bp; NM_017009), F gcacgaggctaatgactatc-3´) / R (5'-atgtccagggctagcttaac-3´).
Gel electrophoresis revealed a single polymerase chain reaction (PCR) product, and the melting curve analysis showed a single peak for all amplification products. The PCR products were sequenced and blasted to confirm the correct identity of each amplicon. Ten-fold serial dilutions generated from cDNA of each sample were used as a reference for the standard curve calculation to determine primer efficiency. Duplicates of all real-time PCR reactions were performed in a 25 µl mixture containing 1/20 volume of the sample cDNA preparation from 250 ng total RNA, 400 nM of each primers, and 1X iQ-SYBR Green Supermix (BioRad,).
All real-time quantifications were performed with the iCycler iQ system (BioRad) and were analyzed by using a two-tailed paired t-test following normalization to a Hprt control. For methodical reasons duplicates of only three animals per group could be analyzed on one plate by real- time PCR. Therefore a two-factorial ANOVA followed by two-tailed poshoc t-test (Bonferroni) was used to compare corresponding plates. Differences were considered significant if p-values were ≤/0.05.
2.2 Immunohistochemistry
Sprague-Dawley rats were deeply anesthetized with isoflurane (1-Chloro-2,2,2-trifluoroethyl-difluoromethylether, Abbott, Wiesbaden, Germany). After thoracotomy, animals were transcardially perfused with 50 ml 0.9% sodium chloride followed by 200 ml 4% phosphate-buffered formaldehyde (10 ml x min-1). The brain was removed and post-fixed for 4 hours with the same fixative at 4°C. Tissues were cryoprotected in 10% sucrose for 2 hours followed by 30% sucrose in 0.1 M phosphate buffer overnight at 4°C and subsequently frozen at -25°C. A series of 30-µm-thick transverse brain sections was cut using a cryostat (Frigocut, Reichert-Jung, Germany). Before any immunohistochemical treatment was performed, free-floating sections were rinsed three times in PBS (pH 7.4) for 15 min each.
The intrinsic peroxidase activity was blocked with hydrogen peroxide-methanol (1:100) for 45 min at room temperature (RT) in the dark. After washing, sections were permeabilized with 0.2% Triton X-100 for 30 min and directly transferred into PBS containing 5% BSA for 1 h at RT to block non-specific binding sites. Sections were incubated at 4°C in rabbit anti-GFAP-antibody (DAKO) solution at a dilution of 1:5,000 overnight and subsequently washed three times for 10 min each. Secondary HRP-conjugated goat anti-rabbit antibodies (1: 1,000 dilution; Dianova) were applied for 2 h at RT and rinsed with PBS three times for 10 min each. After incubation, sections were extensively washed and pre-incubated with freshly prepared filtered diaminobenzidine (DAB)-solution [120 µl DAB (75 mg DAB dissolved in 1.5 ml 0.1 M phosphate buffer) diluted in 30 ml PBS] for 3 min at RT. The enzymatic reaction was started by adding 10 µl of 35% H2O2 to 10 ml DAB solution and stopped with PBS after 1 to 5 min. DAB-stained sections were washed in PBS, mounted onto gelatin-coated microscope-slides, dehydrated (2 x 50% ethanol, 2 x 80% ethanol, and 2 x 99.9% ethanol, 5 min each), cleared with four changes of xylene, coverslipped with mounting medium (DePeX from Serva, Heidelberg, Germany), and finally analyzed with the digital microscope Coolscope (Nikon, Melville, USA) equipped with a 2x and 20x objective. All images were taken under the same lightening and condenser conditions at 2,048 x 2,048 dpi and were imported into Adobe Photoshop CS5, were digitally adjusted if necessary for brightness and contrast, and were assembled into plates. Five sections per animal were analyzed for quantification of GFAP positive cells for each region. Cells were counted at Bregma level 5.64 mm for the frontal cortex, at 1.92 mm for the putamen, and at -3.24 mm for the hilus of the denate gyrus according to the 6th edition of the Rat Brain in Stereotaxic Coordinates (Paxinos and Watson 2007). The values were denoted as cells/10,000 µm2 and presented as mean ± SD. The size of the counting frame was 138,837 µm2.
3. RESULTS
3.1 Frontal Cortex
3.1.1 Real-time PCR
In the group that was fluoxetine treated from day 21 to 35 a significant 48% decrease of
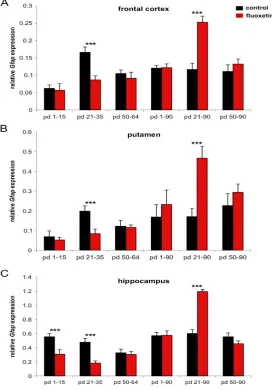
Gfap mRNA expression in the frontal cortex was measured 24 hours after the last injection, while no short-term effects were observed in the other groups (table 1; figure 1A).
For long-term effects, which were measured at day 90, we found an opposite effect in the same group (fluoxetine treatment from day 21 to 35) with a significant 31% increase of Gfap
mRNA in the fluoxetine treated animals as compared to control animals (table 1, figure 1A). In the two other groups, treated from day 1 to 15 or day 50 to 64, respectively, no significant differences were found between the fluoxetine treated animals and the control animals (table 1, figure 1A). An age effect was noted in the control animals i.e. the Gfap mRNA expression increased from day 16 to day 36 and decreased to adult levels at day 65 (figure 1A).
3.1.2 Immunohistochemistry
In the frontal cortex a significantly 20% reduction of GFAP-positive cells was found 24 hours after the last drug injection in the animals treated with fluoxetine from postnatal day 21 to 35. At day 90, a significant 58% increase in GFAP-immunopositive cells was found in this group. When treated earlier (day 1 to 15) or later (day 50 to 64) no significant changes were found 24 hours after last injection or at day 90 (table 2, figure 2). Concerning the development of GFAP-immunoreactive cells over time, an increase from day 16 to 36 was found, followed by a slight decrease until day 65 and no further changes until day 90 (figure 2).
3.2 Striatum
3.2.1 Real-time PCR
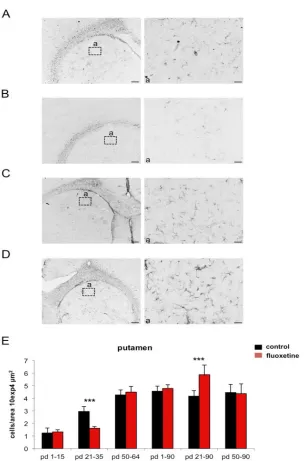
In the putamen of the striatum, the Gfap mRNA expression was significantly 58% decreased in animals, which were fluoxetine treated from day 21 to 35 if measured 24 hours after the last fluoxetine injection (compared to the control animals). Measured at day 90, the fluoxetine treated group revealed a significant 174% increase in Gfap mRNA expression. However, no effects were determined for the other groups (table 1, figure 1B). An age effect was analyzed in the control animals i.e. the Gfap mRNA expression increased from day 16 to adult levels at day 65 (figure 1B).
3.2.2 Immunohistochemistry
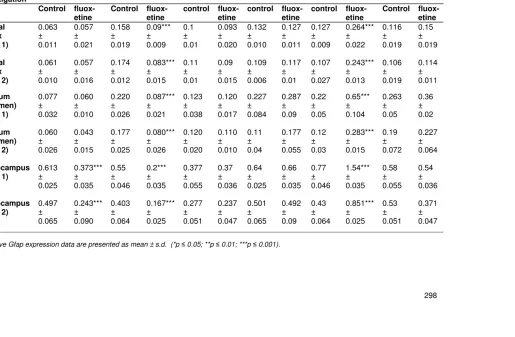
Table 1. Real-time PCR analysis during different development stage and 14-day fluoxetine treatment
Treatment * day of investigation 1-15 * 16 21-35 *36 50-64 *65 1-15 *90 21-35 *90 50-64 *90
Control fluox-etine
Control fluox-etine
control fluox-etine
control fluox-etine
control fluox-etine
Control fluox-etine Frontal cortex (plate 1) 0.063 ± 0.011 0.057 ± 0.021 0.158 ± 0.019 0.09*** ± 0.009 0.1 ± 0.01 0.093 ± 0.020 0.132 ± 0.010 0.127 ± 0.011 0.127 ± 0.009 0.264*** ± 0.022 0.116 ± 0.019 0.15 ± 0.019 Frontal cortex (plate 2) 0.061 ± 0.010 0.057 ± 0.016 0.174 ± 0.012 0.083*** ± 0.015 0.11 ± 0.01 0.09 ± 0.015 0.109 ± 0.006 0.117 ± 0.01 0.107 ± 0.027 0.243*** ± 0.013 0.106 ± 0.019 0.114 ± 0.011 Striatum (Putamen) (plate 1) 0.077 ± 0.032 0.060 ± 0.010 0.220 ± 0.026 0.087*** ± 0.021 0.123 ± 0.038 0.120 ± 0.017 0.227 ± 0.084 0.287 ± 0.09 0.22 ± 0.05 0.65*** ± 0.104 0.263 ± 0.05 0.36 ± 0.02 Striatum (Putamen) (plate 2) 0.060 ± 0.026 0.043 ± 0.015 0.177 ± 0.025 0.080*** ± 0.026 0.120 ± 0.020 0.110 ± 0.010 0.11 ± 0.04 0.177 ± 0.055 0.12 ± 0.03 0.283*** ± 0.015 0.19 ± 0.072 0.227 ± 0.064 Hippocampus (plate 1) 0.613 ± 0.025 0.373*** ± 0.035 0.55 ± 0.046 0.2*** ± 0.035 0.377 ± 0.055 0.37 ± 0.036 0.64 ± 0.025 0.66 ± 0.035 0.77 ± 0.046 1.54*** ± 0.035 0.58 ± 0.055 0.54 ± 0.036 Hippocampus (plate 2) 0.497 ± 0.065 0.243*** ± 0.090 0.403 ± 0.064 0.167*** ± 0.025 0.277 ± 0.051 0.237 ± 0.047 0.501 ± 0.065 0.492 ± 0.09 0.43 ± 0.064 0.851*** ± 0.025 0.53 ± 0.051 0.371 ± 0.047
3.3 Hippocampus
3.3.1 Real-time PCR
In the hippocampus, short-term effects were detected in the very early treated group (from day 1 to 15). Gfap mRNA expression was significantly 44% decreased compared to the control at day 16. A comparable 61% Gfap mRNA reduction was seen in the animals treated with fluoxetine from day 21 to 35 if measured at day 36 and compared to control animals (table 1, figure 1C). At day 90, there was no difference between fluoxetine treated animals from day 1-15 or day 50- 64, but we found a significant 99% increase in Gfap mRNA expression in the animals treated with fluoxetine from day 21- 35 compared to control animals (table 1, figure 1C).
Fig. 1. Developmental Gfap-mRNA expression under control versus fluoxetine treated conditions. (A) frontal cortex, (B) striatum (putamen), and (C) hippocampus.
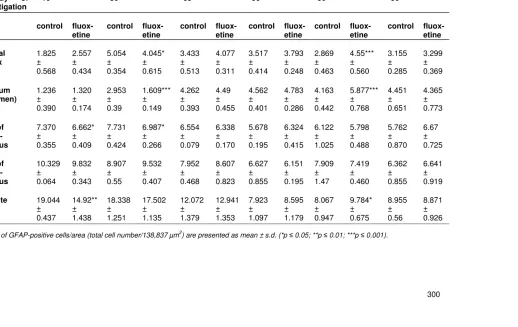
Table 2. Immunohistochemistry during different development stage and 14-day fluoxetine treatment
Treatment
* day of
investigation 1-15 * 16 21-35 *36 50-64 *65 1-15 *90 21-35 *90 50-64 *90
control fluox-etine
control fluox-etine
control fluox-etine
control fluox-etine
control fluox-etine
control fluox-etine Frontal cortex 1.825 ± 0.568 2.557 ± 0.434 5.054 ± 0.354 4.045* ± 0.615 3.433 ± 0.513 4.077 ± 0.311 3.517 ± 0.414 3.793 ± 0.248 2.869 ± 0.463 4.55*** ± 0.560 3.155 ± 0.285 3.299 ± 0.369 Striatum (Putamen) 1.236 ± 0.390 1.320 ± 0.174 2.953 ± 0.39 1.609*** ± 0.149 4.262 ± 0.393 4.49 ± 0.455 4.562 ± 0.401 4.783 ± 0.286 4.163 ± 0.442 5.877*** ± 0.768 4.451 ± 0.651 4.365 ± 0.773 CA1 of hippo-campus 7.370 ± 0.355 6.662* ± 0.409 7.731 ± 0.424 6.987* ± 0.266 6.554 ± 0.079 6.338 ± 0.170 5.678 ± 0.195 6.324 ± 0.415 6.122 ± 1.025 5.798 ± 0.488 5.762 ± 0.870 6.67 ± 0.725 CA3 of hippo-campus 10.329 ± 0.064 9.832 ± 0.343 8.907 ± 0.55 9.532 ± 0.407 7.952 ± 0.468 8.607 ± 0.823 6.627 ± 0.855 6.151 ± 0.195 7.909 ± 1.47 7.419 ± 0.460 6.362 ± 0.855 6.641 ± 0.919 Dentate gyrus 19.044 ± 0.437 14.92** ± 1.438 18.338 ± 1.251 17.502 ± 1.135 12.072 ± 1.379 12.941 ± 1.353 7.923 ± 1.097 8.595 ± 1.179 8.067 ± 0.947 9.784* ± 0.675 8.955 ± 0.56 8.871 ± 0.926
Fig. 2. Developmental GFAP expression after fluoxetine treatment in the frontal cortex. (A) GFAPexpression under control conditions at postnatal day (pd) 36. (B) GFAP expression after fluoxetine treatment (from pd 21 – pd 35) at pd 36. (C) GFAP expression under control conditions at postnatal day (pd) 90. (D) GFAPexpression after fluoxetine treatment (from pd 21 – pd 35) at pd 90. In (a) GFAP positive cells are
shown at a higher magnification. Scale bars = 200 µm for overview and 20 µm for higher magnification (a). (E) Bar diagram represents GFAP-labeled cells per area during different developmental stages under control versus fluoxetine treated
conditions.
Fig. 3. Developmental GFAP expression after fluoxetine treatment in the putamen. (A) GFAP expression under control conditions at postnatal day (pd) 36. (B) GFAP
expression after fluoxetine treatment (from pd 21 – pd 35) at pd 36. (C) GFAP expression under control conditions at postnatal day (pd) 90. (D) GFAP expression after fluoxetine treatment (from pd 21 – pd 35) at pd 90. In (a) GFAP positive cells are
shown at a higher magnification. Scale bars = 200 µm for overview and 20 µm for higher magnification (a). (E) Bar diagram represents GFAP-labeled cells per area
3.3.2 Immunohistochemistry
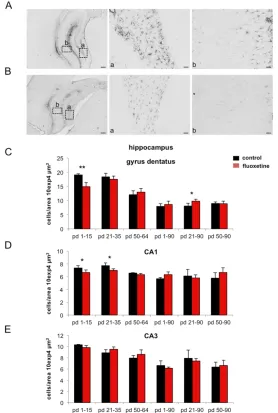
In the hippocampus GFAP-positive cells were counted in three different regions. In the CA1
region a significant 10% decrease of GFAP-immunoreactivity was found 24 day 16 to 65 in the CA1 region (figure 4D).
Fig. 4. Developmental GFAP expression after fluoxetine treatment in the
hippocampus. (A) GFAP expression under control conditions at postnatal day (pd) 16. (B) GFAP expression after fluoxetine treatment (from pd 1 – pd 15) at pd 16. Expression of GFAP in the dentate gyrus (a) or CA1 hippocampal region (b). Scale
bars = 200 µm for overview and 20 µm for higher magnification (a, b). (C - E) Bar diagrams represent GFAP-labeled cells per area during different development stages
under control versus fluoxetine treated conditions in the dentate gyrus (C), CA1 (D), and CA3 (E).
However, the CA3 region showed no differences in all six groups (table 2, figure 4E). The developmental profile was comparable to that seen in the CA1 region (figure 4E).
In the dentate gyrus there was a significant 22% reduction of GFAP-immunopositve cells 24 hours after the last injection in the fluoxetine treated groups, which were fluoxetine treated from day 1 to 15 and from day 21 to 35, while there was no obvious change in the last group treated from day 50 to 64. In addition, no long-term changes could be observed at day 90 (table 2, figure 4D).
The developmental profile showed a slight decrease in GFAP-immunopositive cells from hours after the last injection in the group fluoxetine treated from day 1 to 15 compared to the control group. No changes were detected after 24 hours in the other two groups. Long-term changes were obvious at day 90 in the group treated from day 21 to 35. A significant 21% increase of GFAP expressing cells was measured in the fluoxetine treated group if compared to the control group (table 2, figure 4 C).
The developmental profile was comparable to that of CA1 (table 2, figure 4 C).
4. DISCUSSION
This is the first study examine the potential influence of chronic fluoxetine administration on postnatal astroglial development.
Assuming that antidepressants do not only influence neuronal but also glial development, this study revealed short- and long-term changes of GFAP protein and Gfap mRNA expression after fluoxetine administered to rats during different postnatal developmental periods. GFAP is an established astroglial marker (Bignami et al., 1972; Bushong et al., 2002) and seems to be an adequate parameter, since an increasing number of studies showed direct connections between alterations of astrocytes, depression, and SSRI treatment (Lee et al., 2007; Cabras et al., 2010; Schipke et al., 2011).
So far, almost all published animal studies used adult rats. However, it needs to be considered that depression is also a developmental problem, which may be successfully treated in children with the clinically approved drug fluoxetine (Usala et al., 2008). Recently it was shown that prenatal fluoxetine treatment promotes gliogenesis during neuronal differentiation in mouse embryonic stem cells (Si et al., 2004).Therefore, in elucidating the treatment mechanism, it is of interest, whether SSRI treatment changes GFAP/Gfap
expression during postnatal brain development.
4.1 Very Early Postnatal Period (Day 1-15)
A significant reduction of GFAP mRNA 24 hours after treatment from day 1 to 15 could only be found in the hippocampus. At the protein level, no changes were detected in the CA3 region, while in CA1 and in the dentate gyrus we found a significant decrease of GFAP positive cells 24 hours after fluoxetine treatment during day 1 to 15. It has been shown that after two weeks of fluoxetine treatment, an increase of serotonin concentration occurs (Le Prince et al., 1990). This could be partially responsible for the obvious reduction of Gfap
Moreover, there are a lot of other molecular mechanisms possible, which could be mediated independently of serotonin e.g. stimulation of S-100B by fluoxetine via an unknown mechanism independent of serotonin was described recently (Tramonita et al., 2008).
The very early locally restricted short-term effect could be related to the normal developmental profile of GFAP expression. The hippocampus is the region where we found the earliest maximum of GFAP expression, which was reached shortly after birth, whereas in the frontal cortex and striatum the maximum of GFAP expression occurred later leading to a different addressability for serotonin. Long-term changes were not observed at all.
4.2 Periadolescent Period (Day 21-35)
This is a very important period, because during the periadolescence a wave of overproduction and elimination of synapses and receptors occurs. This developmental phenomenon has been observed in humans where the synaptic density in the frontal cortex decreases by approximately 40% between 7 and 15 years (Huttenlocher, 1979), primates (Lidow et al., 1991) and rats (Teicher et al., 2003).
Our main results are related to this “periadolescent period” during day 21 to 35.
We found a reduction of GFAP/Gfap expression when animals were investigated 24 hours after the end of fluoxetine treatment during day 21 to 35. The GFAP mRNA expression and the GFAP positive labeled cells were decreased compared to controls in frontal cortex, striatum, and hippocampus. This short- time response resembles effects of stress induced loss of GFAP (Leventopoulos et al., 2007; Ye et al., 2011), while the observed long term changes only found at this developmental stage resemble changes related to uncontrolled stress reactions which are not successful compensated and results therefore in an astroglial reaction. Barros et al., 2006 described that prenatal stress results in a significant increase in GFAP immunoreactive astrocytes with high levels of S-100B protein in frontal cortex, striatum and hippocampus at postnatal day 90. Those changes were similar to neurtotoxic drugs effects. Prenatal exposure of perfluorooctane sulfonate (PFOS) increased GFAP and S-100B, which were accompanied with elevation of pro-inflammatory cytokines (interleukin1ß and tumor necrosis factor (TNF)-alpha) and pro-inflammatory transcription factors (activation protein-1 (AP-1), nuclear-factor kappaB (NF-kappaB), and cAMP response-binding protein (CREB)) (Zeng et al., 2011). Although fluoxetine is able to increase the nitric oxide production via nuclear factor kappa B- mediated pathway (Ha et al., 2006), others found that the elevation of NF-kappa B activity was not an integral part of the apoptotic signaling cascade induced neuronal cell death caused by fluoxetine (Bartholomä et al., 2002).
relationship between neurogenesis and the exact influence of astroglia has still to be determined.
Moreover, the signal transducer and activator of transcription 3 (STAT3) is abundantly expressed in the postnatal brain (Gautron et al., 2006) and was found to be altered by SSRI (Sillaber et al., 2008). STAT3 in turn regulates GFAP expression after brain injuries (Xu et al., 2011). Interestingly, early activation of both STAT3 and NFkappaB might alter gene expression, which finally can lead to gliosis (Acarin et al., 2000).
Nevertheless, the most interesting part of our findings is the fact that the findings differ depending on time frame of drug exposure.
This period’s specific sensitivity to the drug might be supported by the studies of Wegerer et al., 1999, who found a significant increase of 5-HTT in the frontal cortex of adult rats after fluoxetine treatment (5mg/kg/BW p.o.) for two weeks from day 25 to 39, while treatment during later developmental stage (day 50 to 64) revealed no significant changes. The authors assumed that 5-HTT overexpression might be due to a higher density of serotonergic innervations. In conjunction with our results of the long-lasting changes of GFAP the increase of 5-HTT could be additionally due to an astrogliosis and/or to an increase of immature stem cells, which might be GFAP positive and which are able to
differentiate into neurons or glia (Steiner et al., 2004).
Not only fluoxetine exerts such a special “time frame of action” with long-lasting changes. Methylphenidate treatment from day 21 to 35 also reduces dopamine transporter density in the striatum of adult rats, while later treatment has no consequence (Roessner et al., 2010). Furthermore, (Moll et al., 2000), described the normal postnatal development of monoaminergic transporters in the frontal cortex and striatum and found that the most significant changes occurred from day 25 to 50.
A special time frame of fluoxetine induced behavioral changes in mice was described by (Ansorge et al., 2004; Ansorge et al., 2008). They found that postnatal fluoxetine treatment from day 4 to 21 is accompanied by abnormal emotional behavior in adult mice, while later treatment leads to no visible behavioral alterations. The development of rats and mice are not directly comparable, but it would be of great interest, if our findings are related to behavioral changes in adult rats, too.
4.3 Postadolescent Period (Day 50-65)
of structural plasticity, while antidepressants possibly act by compensating this dysfunction (Manji et al., 2003).
Other researchers described that chronic activation of the stress response during rat pregnancy resulted in a significant increase of GFAP-immunoreactive astrocytes in frontal cortex, striatum, and hippocampus in the pups (Barros et al., 2006), and theses changes are associated with behavioral and learning impairment (Weinstock, 2001).
These studies represent a different influence of chronic stress, which also depends on the timeframe of exposure as shown for the fluoxetine treatment in this study.
4.4 Normal Developmental Profile
of GFAP
Comparing only the GFAP immunoreactivity of the control animals of each investigated
group suggests a region specific developmental pattern. In the frontal cortex an increase of GFAP was seen from day 16 to 36 followed by a slight decrease at day 65, which remained the same until day 90. In the striatum (putamen) an increase of GFAP from day 16 to 65 occurred and remained until day 90. No significant developmental changes were found in the hippocampus CA1 and CA3 regions, while there was a decrease in the dentate gyrus from day 16 to 90. The developmental changes are in accordance with other studies, which described alterations in glia cells during postnatal development. During mouse brain maturation, Gfap mRNA showed a peak at postnatal day 3, a decrease until day 15 followed by a slight increase until day 55 (Riol et al., 1992) thus indicating a two-step developmental expression. It increases between birth and day 15 (period of astrocyte proliferation) and then decreases until day 55 (period of morphological astrocyte differentiation) (Tardy et al., 1989). With regard to the region specific investigations using immunohistochemistry, the number of GFAP positive astrocytes was increased in the hippocampal CA1 region in a one week old mouse compared to an 8 week old, but remained unchanged from week 2 to 8 (Ogata and Kosaka, 2002). Thus, our normal developmental profiles are in line with literature data and reflect validity of the data.
5. CONCLUSION
We suggest that a critical and drug-sensitive (i. e. periadolescence) period exists, which is closely related to very active glial-neuronal development and where psychopharmacological treatment results in long-lasting structural changes. This study showed that fluoxetine treatment in rats leads to a long-lasting increase of astroglia. Probably this critical period is highly sensitive to drugs since it is usually accompanied by overproduction of receptors and synapses followed by pruning or competitive elimination. This is not only observed in rats but also in primates and humans. For example, in humans the synaptic density in the frontal cortex decreases by approximately 40% between 7 and 15 years of age (Huttenlocher, 1979). Thus, psychopharmacological effects during this time frame (i.e. pre-adolescence) seem to be of great importance, since many children suffering from psychiatric disorders are treated with psychopharmaca within this sensitive age range.
factors involved in these adaptive processes and whether such changes will be accompanied by behavioral changes either to the better or the worse.
COMPETING INTERESTS
Authors have declared that no competing interests exist.
REFERENCES
Acarin, L., Gonzalez, B., Castellano, B. (2000). STAT3 and NFkappaB activation precedes glail reactivity in the excitotoxically injured young cortex but not in the corresponding distal thalamic nuclei. J Neuropathol Exp Neurol, 59, 151-163.
Allaman, I., Fiumelli, H., Magistretti, P., Martin, J. (2011). Fluoxetine regulates the expression of neurotrophic/growth factors and glucose metabolism in astrocytes. Psychopharmacology (Berl), 216, 75–84.
Althaus, H.H., Richter-Landsberg. C. (2000). Glial cells as targets and producers of neurotrophins. Int Rev Cytol, 197, 203-277.
Anderson, C.M., Swanson, R.A. (2000). Astrocyte glutamate transport: review of properties, regulation, and physiological functions. Glia, 32, 1-14.
Ansorge, M.S., Morelli, E., Gingrich, J.A. (2008). Inhibition of serotonin but not norepinephrine transport during development produces delayed, persistent perturbations of emotional behaviors in mice. J Neurosci, 28, 199-207.
Ansorge, M.S., Zhou, M., Lira, A., Hen, R., Gingrich, J.A. (2004). Early-life blockade of the 5-HT transporter alters emotional behavior in adult mice. Science, 306, 879-881. Azmitia, E.C. (2001). Modern views on an ancient chemical: serotonin effects on cell
proliferation, maturation, and apoptosis. Brain Res Bull, 56, 413-424.
Azmitia, E.C., Gannon, P.J., Kheck, N.M., Whitaker-Azmitia, P.M. (1996). Cellular localization of the 5-HT1A receptor in primate brain neurons and glial cells. Neuropsychopharmacology, 14, 35-46.
Bachis, A., Mallei, A., Cruz, M.I., Wellstein, A., Mocchetti, I. (2008). Chronic antidepressant treatments increase basic fibroblast growth factor and fibroblast growth factor-binding protein in neurons. Neuropharmacology, 55, 1114-1120.
Banasr, M., Duman, R.S. (2008). Glial loss in the prefrontal cortex is sufficient to induce depressive-like behaviors. Biol Psychiatry, 64, 863-870.
Barros, V.G., Duhalde-Vega, M., Caltana, L., Brusco, A., Antonelli, M.C. (2006). Astrocyte-neuron vulnerability to prenatal stress in the adult rat brain. J Neurosci Res, 83, 787-800.
Bartholomä, P., Erlandsson, N., Kaufmann, K., Rössler, O.G., Baumann, B., Wirth, T., Giehl, K.M., Thiel, G. (2002). Neuronal cell death induced by antidepressants: lack of correlation with Erg-1, NF-kappa B and extracellular signal-regulated protein kinase activation. Biochem Pharmacol, 63, 1507-1516.
Bignami, A., Eng, L.F., Dahl, D., Uyeda, C.T. (1972). Localization of the glial fibrillary acidic protein in astrocytes by immunofluorescence. Brain Res Bull, 43, 429–435.
Blier, P., de Montigny, C. (1999). Serotonin and drug-induced therapeutic responses in major depression, obsessive-compulsive and panic disorders. Neuropsychopharmacology, 21, 91S-98S.
the serotonin but not the noradrenaline transporter in the frontal cortex of rats. World J Biol Psychiatry, 6, 107-112.
Bushong, E.A., Martone, M.E., Jones, Y.Z., Ellisman, M.H. (2002). Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J Neurosci, 22, 183-192.
Cabras, S., Saba, F., Reali, C., Scorciapino, M.L., Sirigu, A., Talani, G., Biggio, G., Sogos, V. (2010). Antidepressant imipramine induces human astrocytes to differentiate into cells with neuronal phenotype. Int J Neuropsychopharmacol, 13, 603-615.
Chen, Y., Peng, L., Zhang, X., Stolzenburg, J.U., Hertz, L. (1995). Further evidence that fluoxetine interacts with a 5-HT2C receptor in glial cells. Brain Res Bull, 38, 153-159. Cotter, D., Mackay, D., Landau, S., Kerwin, R., Everall, I. (2001). Reduced glial cell density and neuronal size in the anterior cingulate cortex in major depressive disorder. Arch Gen Psychiatry, 58, 545-553.
Czeh, B., Simon, M., Schmelting, B., Hiemke, C., Fuchs, E. (2006). Astroglial plasticity in the hippocampus is affected by chronic psychosocial stress and concomitant fluoxetine treatment. Neuropsychopharmacology, 31, 1616-1626.
Duman, R.S., Nakagawa, S., Malberg, J. (2001). Regulation of adult neurogenesis by antidepressant treatment. Neuropsychopharmacology, 25, 836-844.
Fatemi, S.H., Laurence, J.A., Araghi-Niknam, M., Stary, J.M., Schulz, S.C., Lee, S., Gottesmann, I.I. (2004). Glial fibrillary acidic protein is reduced in cerebellum of subjects with major depression, but not schizophrenia. Schizophr Res, 69, 317-323. Fitzgerald, L.W., Kaplinsky, L., Kimelberg, H.K. (1990). Serotonin metabolism by monoamine
oxidase in rat primary astrocyte cultures. J Neurochem, 55, 2008-2014.
Friedman, W.J., Black, I.B., Kaplan, D.R. (1998). Distribution of the neurotrophins brain-derived neurotrophic factor, neurotrophin-3, and neurotrophin-4/5 in the postnatal rat brain: an immunocytochemical study. Neuroscience, 84, 101-114.
Gautron, L., De Smedt-Peyrusse, V., Laye, S. (2006). Characterization of STAT3-expressing cells in the postnatal brain. Brain Res, 1098, 26-32.
Ha, E., Jung, K.H., Cjoe, B.K., Shin, D.H., Yim, S.V., Baik, H.H. (2006). Fluoxetine increases the nitric oxide production via nuclear factor kappa B-mediated pathway in BV2 murine microglial cells. Neurosci Lett, 597, 185-189.
Hansson, E., Rönnbäck, L. (1992). Adrenergic receptor regulation of amino acid neurotransmitter uptake in astrocytes. Brain Res Bull, 29, 297-301.
Hensler, J.G. (2006). Serotonergic modulation of the limbic system. Neurosci Biobehav Rev, 30, 203-214.
Huttenlocher, P.R. (1979). Synaptic density in human frontal cortex - developmental changes and effects of aging. Brain Res, 163, 195-205.
Karpova, N.N., Lindholm, J., Pruunsild, P., Timmusk, T., Castren, E. (2009). Long-lasting behavioural and molecular alterations induced by early postnatal fluoxetine exposure are restored by chronic fluoxetine treatment in adult mice. Eur Neuropsychopharmacol, 19, 97-108.
Kimelberg, H.K., Katz, D.M. (1985). High-affinity uptake of serotonin into immunocytochemically identified astrocytes. Science, 228, 889-891.
Kimoto, H., Eto, R., Abe, M., Kato, H., Araki, T. (2009). Alterations of glial cells in the mouse hippocampus during postnatal development. Cell Molecular Neurobiology, 29, 1181-1189.
Le Prince, G., Copin, M.C., Hardin, H., Belin, M.F., Bouilloux, J.P., Tardy, M. (1990). Neuron-glia interactions: effect of serotonin on the astroglial expression of GFAP and of its encoding message. Brain Res Dev Brain Res, 51, 295-298.
Lee, Y., Gaskins, D., Anand, A., Shekhar, A. (2007). Glia mechanisms in mood regulation: a novel model of mood disorders. Psychopharmacology (Berl), 191, 55-65.
Leventopoulos, M., Ruedi-Bettschen, D., Knuesel, I., Feldon, J., Pryce, C.R., Opacka-Juffry, J. (2007). Long-term effects of early life deprivation on brain glia in Fischer rats. Brain Res, 1142, 119-126.
Lidov, H.G., Molliver, M.E. (1982). An immunohistochemical study of serotonin neuron development in the rat: ascending pathways and terminal fields. Brain Res Bull, 8, 389-430.
Lidow, M.S., Goldman-Rakic, P.S., Rakic, P. (1991). Synchronized overproduction of neurotransmitter receptors in diverse regions of the primate cerebral cortex. Proc Natl Acad Sci, U S A, 88, 10218–10221.
Luellen, B.A., Miller, D.B., Chisnell, A.C., Murphy, D.L., O'Callaghan, J.P., Andrews, A.M. (2003). Neuronal and astroglial responses to the serotonin and norepinephrine neurotoxin: 1-methyl-4-(2'-amino-phenyl)-1,2,3,6-tetrahydropyridine. Journal of Pharmacological and Experimental Therapy, 307, 923-931.
Malberg, J.E., Duman, R.S. (2003). Cell proliferation in adult hippocampus is decreased by inescapable stress: reversal by fluoxetine treatment. Neuropsychopharmacology, 28, 1562-1571.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S. (2000). Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci, 20, 9104-9110.
Manji, H.K., Quiroz, J.A., Sporn, J., Payne, J.L., Denicoff, K., N A.G., Zarate, C.A., Jr. Charney, D.S. (2003). Enhancing neuronal plasticity and cellular resilience to develop novel, improved therapeutics for difficult-to-treat depression. Biol Psychiatry, 53, 707-742.
Moll, G.H., Hause, S., Ruther, E., Rothenberger, A., Huether, G. (2001). Early methylphenidate administration to young rats causes a persistent reduction in the density of striatal dopamine transporters. J Child Adolesc Psychopharmacol, 11, 15-24.
Moll, G.H., Mehnert, C., Wicker, M., Bock, N., Rothenberger, A., Ruther, E., Huether, G. (2000). Age-associated changes in the densities of presynaptic monoamine transporters in different regions of the rat brain from early juvenile life to late adulthood. Brain Res Dev Brain Res, 119, 251-257.
Mongeau, R., Blier, P., de Montigny, C. (1997). The serotonergic and noradrenergic systems of the hippocampus: their interactions and the effects of antidepressant treatments. Brain Res Brain Res Rev, 23, 145-195.
Ogata, K., Kosaka, T. (2002). Structural and quantitative analysis of astrocytes in the mouse hippocampus. Neuroscience, 113, 221-233.
Rajkowska, G. (2000). Postmortem studies in mood disorders indicate altered numbers of neurons and glial cells. Biol Psychiatry, 48, 766-777.
Rajkowska, G., Miguel-Hidalgo, J.J., Wei, J., Dilley, G., Pittman, S.D., Meltzer, H.Y., Overholser, J.C., Roth, B.L., Stockmeier, C.A. (1999). Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol Psychiatry, 45, 1085-1098.
Roessner, V., Sagvolden, T., Dasbanerjee, T., Middleton, F.A., Faraone, S.V., Walaas, S.I., Becker, A., Rothenberger, A., Bock, N. (2010). Methylphenidate normalizes elevated dopamine transporter densities in an animal model of the ADHD combined type, but not to the same extent in one of the ADHD inattentive type. Neurosci Behav Physiol, 167, 1183-1191.
Rubenstein, J.L. (1998). Development of serotonergic neurons and their projections. Biol Psychiatry, 44, 145-150.
Santarelli, L., Saxe, M., Gross, C., Surget, A., Battaglia, F., Dulawa, S., Weisstaub, N., Lee, J., Duman, R., Arancio, O., Belzung, C., Hen, R. (2003). Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science, 301, 805-809.
Schipke, C.G., Heuser, I., Peters, O. (2011). Antidepressants act on glial cells: SSRIs and serotonin elicit astrocyte calcium signaling in the mouse prefrontal cortex. Psychiatr Res, 45, 242-248.
Si, X., Miguel-Hidalgo, J.J., O'Dwyer, G., Stockmeier, C.A., Rajkowska, G. (2004). Age-dependent reductions in the level of glial fibrillary acidic protein in the prefrontal cortex in major depression. Neuropsychopharmacology, 29, 2088-2096.
Sillaber, I., Panhuysen, M., Henniger, M.S., Ohl, F., Kuhne, C., Putz, B., Pohl, T., Deussing, J.M., Paez-Pereda, M., Holsboer, F. (2008). Profiling of behavioral changes and hippocampal gene expression in mice chronically treated with the SSRI paroxetine. Psychopharmacology (Berl), 200, 557-572.
Steiner, B., Kronenberg, G., Jessberger, S., Brandt, M.D., Reuter, K., Kempermann, G. (2004). Differential regulation of gliogenesis in the context of adult hippocampal neurogenesis in mice. Glia, 46, 41-52.
Stockmeier, C.A., Mahajan, G.J., Konick, L.C., Overholser, J.C., Jurjus, G.J., Meltzer, H.Y., Uylings, H.B., Friedman, L., Rajkowska, G. (2004). Cellular changes in the postmortem hippocampus in major depression. Biol Psychiatry, 56, 640-650.
Tardy, M., Fages, C., Riol, H., LePrince, G., Rataboul, P., Charriere-Bertrand, C., Nunez, J. (1989). Developmental expression of the glial fibrillary acidic protein mRNA in the central nervous system and in cultured astrocytes. J Neurochem, 52, 162-167. Teicher, M.H., Krenzel, E., Thompson, A.P., Andersen, S.L. (2003). Dopamine receptor
pruning during the peripubertal period is not attenuated by NMDA receptor antagonism in rat. Neurosci Lett, 339, 169-171.
Tramonita, A.C., Tramonita, F., Bobermin, L.D., Zanotto, C., Souza, F.D., Leite, M.C., Nardin, P., Gottfried, C., C.-A. G (2008). Serotonin of S100B, an astrocyte-derived neurotrophic protein, is stimulated by fluoxetine via a mechanism independent of serotonin. Progrtess in Neuro-Psychopharmacology & Biological Psychiatry, 32, 1580-1583.
Usala, T., Clavenna, A., Zuddas, A., Bonati, M. (2008). Randomised controlled trials of selective serotonin reuptake inhibitors in treating depression in children and adolescents: a systematic review and meta-analysis. Eur Neuropsychopharmacol, 18, 62-73.
Webster, M.J., O’Grady, J., Kleinmann, J.E., Weickert, C.S. (2005). Glial fibrillary acidic protein mRNA levels in the cingulate cortex of individuals with depression, bipolar disorder and schizophrenia. Neurosci Behav Physiol, 133, 453–461.
Wegerer, V., Moll, G.H., Bagli, M., Rothenberger, A., Ruther, E., Huether, G. (1999). Persistently increased density of serotonin transporters in the frontal cortex of rats treated with fluoxetine during early juvenile life. J Child Adolesc Psychopharmacol, 9, 13-24; discussion, 25-16.
Weinstock, M. (2001). Alterations induced by gestational stress in brain morphology and behaviour of the offspring. Prog Neurobiol, 65, 427-451.
Xu, Z., Xue, T., Zhang, Z., Wang, X., Xu, P., Zhang, J.K., Lei, X., Li, Y., Xie, Y., Wang, L., Fang, M., Chen, Y. (2011). Role of signal transducer and activator of transcription-3 in up-regulation of GFAP after epilepsy. Neurochem Res, 36, 2208-2215.
Ye, Y., Wang, G.J., AWang, H., Wang, X. (2011). Brain-derived factor (BDNF) infusion restored astrocytic plasticity in the hippocampus of a rat model of depression. Neurosci Lett, 503, 15-19.
Zeng, H.C., Zhang, L., Li, Y.Y., Wang, Y.J., Xia, W., Lin, Y., Wei, J., Xu, S.Q. (2011). Inflammation-like glial response in rat brain induced by prenatal PFOS exposure. Neurotoxology, 32, 130-139.
Zhang, Y., Barres, B.A. (2010). Astrocyte heterogeneity: an underappreciated topic in neurobiology. Curr Opin Neurobiol, 20, 588-594.