Copyright 0 1995 by the Genetics Society of America
Activity, Expression and Function of a Second Drosophila
Protein Kinase A
Catalytic Subunit Gene
Alicia
Melkndez,
Willis
Li
and Daniel Kalderon
Department of Biological Sciences, Columbia University, New York, New York 10027 Manuscript received July 13, 1995
Accepted for publication September 13, 1995
ABSTRACT
The DC2 gene was isolated previously on the basis of sequence similarity to DCO, the major Drosophila protein kinase A (PKA) catalytic subunit gene. We show here that the 67-kD Drosophila DC2 protein behaves as a PKA catalytic subunit in vitro. DC2 is transcribed in mesodermal anlagen of early embryos. This expression depends on dorsal but on neither twist nor snail activity. DC2 transcriptional fusions mimic this embryonic expression and are also expressed in subsets of cells in the optic lamina, wing disc and leg discs of third instar larvae. A saturation screen of a small deficiency interval containing DC2 for recessive lethal mutations yielded no DC2 alleles. We therefore isolated new deficiencies to generate deficiency tranpheterozygotes that lacked DC2 activity. These animals were viable and fertile. The absence of DC2 did not affect the viability or phenotype of imaginal disc cells lacking DCO activity or embryonic hatching of animals with reduced DCO activity. Furthermore, transgenes expressing DC2 from a DCO promoter did not efficiently rescue a variety of DCO mutant phenotypes. These observations indicate that DC2 is not an essential gene and - is unlikely to be functionally redundant with DCO, which has multiple unique functions during development.
T
HE CAMP signal transduction pathway is a classic example of how extracellular signals can lead to intracellular biochemical changes. The traditional view of this pathway is that extracellular signaling molecules engage serpentine receptors, leading to the activation of G proteins and consequent modulation of the rate of synthesis of intracellular CAMP by adenylyl cyclase. The inactive CAMP-dependent protein kinase A (PKA) holoenzyme (R2C2) is activated by dissociation after binding of CAMP to regulatory subunit (R) and is re- sponsible for propagating the vast majority of intracellu- lar CAMP signals in eukaryotes by phosphorylation of target proteins on serine or threonine residues (re- viewed in FRANCIS and CORBIN 1994).In metazoa, PKA fulfills many different functions in different cell types. Although selective expression of receptors, PKA substrates and other downstream ef- fectors clearly dictates both the competence to respond to specific signals and the nature of the CAMP-mediated response, it is not clear whether the expression of par- ticular PKA isozymes also contributes to the sensitivity or quality of the response (discussed in DBSKELAND et al. 1993). There is an increased diversity of genes for PKA subunits in progressively more complex metazoa, but this is not necessarily indicative of function. For example, in Saccharomyces cereuisiae, any one of the three catalytic subunit genes ( TPKl-3) can undertake all of the known functions of PKA in this unicellular yeast (TODA et al. 1987b).
Comespondace to: Daniel Kalderon, Department of Biological Sci- ences, 1212 Amsterdam Ave., New York, NY 10027-7004.
Genetics 141: 1507-1520 (December, 1995)
The known biochemical properties of different PKA isozymes do provide some definitive ideas about how isozymes could contribute to functional specificity. Two biochemically distinct types of regulatory subunit are found in mammals and in Drosophila but not in Dic- tyostelium or S. cereuisiae (FOSTER et al. 1984; TODA et al. 1987a; LU et al. 1990; FRANCIS and CORBIN 1994). A
major difference between these proteins is the ability of type I1 isoforms to bind to members of a large family of A-kinase anchoring proteins that may be tethered to cytoskeletal structures or other signaling molecules
1508 A. Melendez, W. Li and D. Kalderon
and the sensitivity of activation to changes in CAMP concentration.
Further circumstantial evidence for the possible im- portance of isozyme diversity comes from the observa- tion that several PKA subunits are differentially ex- pressed in a variety of tissues in mammals and in Aplysia
(EPPLER et al. 1982; CADD and MCKNIGHT 1989). The major Drosophila PKA catalytic subunit, DCO, is widely expressed and has essential functions during oogenesis, embryogenesis and imaginal development (LANE and
KALDERON, 1993,1994; LI et al. 1995). Here we describe a second Drosophila PKA catalytic subunit, DC2, that is expressed only in limited subsets of cells during devel- opment. Although DC2 has some distinctive biochemi- cal properties that might allow it to act differently to DCO, we find that DC2 has no essential role during development. We also find no evidence for functions shared between DCO and DC2 and demonstrate that DC2 protein cannot substitute efficiently for DCO pro- tein when similarly expressed.
MATERIALS AND METHODS
Expression of Drosophila proteins in bacteria and insect cells: A KpnI site was inserted 5' of the initiation codon (GGTACCAAAAB) of DCO, DC2 and
RT
cDNAs, allowing excision of the complete coding region as a KpnI-XbuI frag- ment in each case. Bacterially synthesized DC2 and RI pro- teins were made using the T7 expression system and purified by sedimentation of inclusion bodies and SDSPAGE before immunization of rabbits to create antibodies as described pre- viously (STUDIER and MOFFATI 1986; LANE and KALDERON1993). Active RI protein was renatured after SDS PAGE puri- fication.
Recombinant baculovirus vectors were made by insertion of KpnI-Xbd cDNA fragments (1.9 kb for DC2, 2.2 kb for
Rl)
into the plasmid pEV55 (O'REILLYand MILLER 1988) followed by co-transfection of pEV55 derivatives with either wild-type baculoviral DNA or an engineered linear baculoviral DNA (PharMingen) into Spodopteru fngiperdu (Sf 21) insect cells using the cationic liposome mediated transfection kit form Invitrogen. Recombinant virus was identified by plaque mor- phology and purified by several rounds of plaque screening as previously described (MILLER 1986) A derivative of the DC2cDNA KpnI-XbuI fragment lacking sequences encoding amino acids 59-120 was also used.
Sf 21 cells were grown at 27" in TGlOO medium (GIBCO, Grand Island, NY) plus 10% fetal calf serum (GIBCO) and seeded at a density of 2.5 X
lo7
cells per 60-mm plate before infection with the desired recombinant virus. At 40-48 hr postinfection, cells were washed twice with cold phosphate- buffered saline (pH 6.2) and lysed in 50 mM Tris (pH 8.0), 150 mM NaCI, 1.0% Nonidet P40, 0.1% aprotinin, 1.0 mM dithiothreitol, 0.35 mM phenylmethylsulfonyl fluoride (PMSF), 10 pM benzamidine, 30 pg/ml leupeptin , 1 pg/ml bacitracin and 10 pg/ml cy-macroglobulin at 4" for 30 min. Insoluble material was removed by centrifugation at 10,000 X g for 15 min at 4". The extracts were assayed fresh or were stored at-80" after rapid freezing in liquid nitrogen.
Concentrations of specific proteins in bacterial extracts were determined by Coomassie staining of gels relative to bovine serum albumin (BSA) standards and were then used as
standards in Western blots to determine DC2 and RI protein concentrations in infected Sf 21 cell extracts.
Kinase assays: Protein kinase assays were performed on adult Drosophila protein extracts and infected Sf 21 cell ex- tracts. Drosophila protein extracts were made by homogeniz- ing 20 adult males (or 40 adult heads) in 200 pl of buffer A containing 10 mM sodium phosphate (pH 6.8), 1 mM EDTA, 0.5 mM EGTA, 2.5 mM 2-mercaptoethanol, 25 mM benzami- dine and 1 mM PMSF. Insoluble material was removed by centrifugation for 15 min at 10,000 X g a t 4". Up to 35 p1 of extract supernatant was assayed in a total volume of 50 pl of kinase buffer containing 50 mM Mops (pH 7.0), 10 mM MgC12, 0.25 mg/ml BSA, Kemptide (generally 100 or 300 pM), 100 pM ATP, [y'*P] ATP (5 Ci/mmol), with or without 10 pM CAMP at 30". Incorporation of label into substrate was measured by spotting onto Whatman P81 phosphocellulose paper every 15 sec for a minute of reaction. The filters were then washed three times in 75 mM phosphoric acid, dried and counted in scintillation fluid (Ultima Gold, Packard) . For inhibition studies, purified Drosophila
RI
in 50 mM Tris HC1 (pH 7.9), 0.5 M NaC1, 10% glycerol, 200 pg/ml PMSF and 25 mM benzamidine was incubated with DC2-containing ex- tracts or control extracts plus purified porcine catalytic sub- unit (Sigma) for 2 hr on ice before kinase assays.Whole mount tissue in situ hybridization: Embryos were incubated as whole-mount preparations with nonradioactive probes made from the DC2 cDNA as described in TAUTZ and PF'EIFLE (1989). Probes consisted of an 895-bp EcoRI fragment corresponding to nucleotides 2-897 from the 5' UTR and a 411-bp EcoRI-ScuI fragment corresponding to nucleotides 2389-2800 from the 3' UTR of the 3.0-kb DC2 cDNA ( K A L DERON and RUBIN 1988).
Fly stocks: The twist allele used was Df(2R)tuis6", the snail allele was sndTra5 and the double mutant stock was sndIGo5
Df(2R)tuiS6'. These and other mutations are described in LINDSLEY and ZIMM (1992), except that the listed cytology for
Df3L)th102 and Df(3L)thll7 is incorrect according to BIU- ZUELA et ul. (1994). Df(3L)A27, isolated in this study, carries a second mutation, readily separated from the 72A deficiency region by meiotic recombination, which is lethal when homo- zygous, giving rise to melanotic tumors in third instar larvae. Adults heterozygous for D f ( 3 L ) h l l or Df(3L)thl02 have a high frequency of duplications of the anterior scutellar bris- tles (-24%) that is increased in trans to Df(3L)A27 (-48%).
Isolation of potential DC2 alleles: For the F2 lethal screen, isogenized BGEl-151 P[w'] 71F males were fed 25 mM ethyl- methane sulfonate (EMS) in 1% sucrose, 10 mM Tris (pH 7.5) as described (GIUGLIATTI 1986) and mated to u; Df(3L)brmll/ TM3, Sbvirgin females. Male progeny (*/TM3, Sb) were indi- vidually mated to u; D f 3 L ) h l l / T M 3 , Sb females. Balanced stocks were derived from vials yielding only T M 3 progeny. These flies were then assayed for complementation of
Df(3L)stfl3, Df(3L)thll7and interse to establish complementa- tion groups.
Sequencing of DC2 coding region from mutant chromo-
somes: For one or cwo alleles from each potential DC2 com- plementation group, total RNA from adult heterozygotes was isolated and converted to cDNA clones containing the entire
DC2 coding region in a Bluescript vector (Stratagene) by re- verse transcription and PCR amplification of a l.6kb piece of DNA. For each allele, equal aliquots of rescued single- stranded DNA derived from seven independent cDNA clones were combined for sequencing using primers that hybridize at 300-bp intervals along the DC2 cDNA.
DC2: rZ Drosophila PKA Isozyme 1 m 1
a D C I Ab a R I Ab a DCOAb
.\I DC:! DC3 R1 R I . D c z *M:
1 S f 2 1
'
E. colr Sf 2 I'
'
S f 2 1'
"
"-
-
67kD-
m
5.5 kD- w
".
-
4HkD-u
I )
NkD-
w d
0
1 2 . 1 - 1 5 h 7
FIGL'RF. 1.-Western blot analysis of Sf 21 cell extracts. Mock-infected Sf 21 cell extracts show no DC2 protein (M;
lane l ) , DC2 and DC2t bacnlo\inlsinfectetI cell lvsates ex-
press high levels of 6i-kD DC2 (lane 2) and the truncated
:i.ikD D ( 2 t (lane 3 ). Drosophila RI purified from E coli (lane
4) ctrmigrated with RI expressed in baculo\inlsinfected Sf 21 cell extracts (lane .5) at 48 kD. In extracts of cells infected with I)C2 baculouirus (lane 7 ) . there was no increase in the expression of an apparent 40-kD Sf 21 DCO homologue rela- tive to control extracts (lane 6 ) .
that failed to complement llf(3l.)lnmll. In the second larger screen, 33 lines failed to complement Df(31a)Q2f5 and 4 also
failed to complement Df(3L)Imnl1. A l l 33 lines were character- ized further by Southern blot analysis using a DC2 cDNA probe and balancer chromosomes w i t h known RFLPs in the region.
Generation of DCOclones: Adult flies carrying DCOmmutant clones marked by loss of the ~rlloro gene were made as de- scribed prcviollsly (XL: and R l w s 1!193; LI r/ al. 199.5) but
using males that were J l~hspiO-jI/~]; Iq~'I251.;
p [ ~ ' .
hsnm FR1140A; l?f(3l4) ImnII/7*M3, .\br;~nd females that were J71f';l)(Xit2, 17 T - , lrsnm
FRT]
40A/ (<YO; Df(31.)A2i/ TM3, .OPT toeliminatc DC2activity in one quarter o f the progeny. Alterna- tively, atlditional DC2 or DCO transgenes werr provided in the cross [female] y f l / r . ~ / ) i O - j 7 / ) ] ; ITy'] 25F, IT?', Irsn~o
FRTJ 4 0 ~ X [male] J p[ T~IKO-D(:~ or T ~ ~ C O - D C O , 9 ' ] : D ( X P
Construction of P[TgDCO-DC21 transgene: A 4.2-kb BnmHI-
Kpnl liagmcnt of grnomic I)%\ Iron1 the IX'O region ex-
tending I'rom 2.8-kh upstream of the transcription start site
to the start codon, together with a 1 .%kb K/n~l-Xl)nl DC2cDNA fragment, were cloned upstream of the S\'.lO I antigen polyad-
enylation signal ( T I I L Y W X . PI al. 1988) in the transformation vector, pDSI3O (MISWX and RLWS l98i). DNA was in-jccted with helprr plasmid pUChspA2-3 into
t
y
'
"
'
'
embryos as dc-scribed (KI'RIS and SPRADI.IS~; 1982). Two independent trans formants were reco\wcd, and the X chromosome insertion was subsequentlv mohilized to yield five additional arttosomal insertions.
0.
. \ I ?, 17 T + , Irsnro FR714011/ QO.
RESULTS
DC2 encodes a protein kinase: Two Drosophila genes, DCI and l X 2 , encode proteins more similar to PKA than to any other protein kinase, including the cGMP- dependent kinases (KM.DERON and
Runrs
1988). How- ever, the sequences of DCl and DC2 differ from the prototypical mousePKA
catalytic subunit, C,, by more than any other known PKA catalvtic subunits, including those of S. c p w l i ~ i n p (TODA ~t nl. 1987b). We therefore tested the activities of DCI and DC2 proteins in 7dm.0 M O C L - C A M P
0 DC2 -CAMP
DC2+cAMP
0 15 30 45 60
T i m (sec)
FI(;~*KF. 'L.-Kin;lsc activity o f IN;!! protcin i n Sf 21 crll
extracts. Transfrr of phospI1;lte from y-'"I'-ATP t o Kenlpitlc is plotted as counts 1x-r Ininutc (elm) ;II successive I.3-sec time points for mock-infcctctl (0,
e)
and I)(;? baclhirus- infrctetl Sf21 cell extracts (0. W ) i n thc ;Ihscncr (0. 0 ) 01'presence ( 0 ,
m)
o f I O pbl ch\lI'. l M 2 containing extracts showed high killasc activity that \vas n o t significantly incrcasrtl bv addition ofc:WP. Mock-inlcctctl rx11';1cts had rnucll l o \ \ wkinase activity that was largely tlcprntlmt on thr addition o f CAMP. I)<:I-cont;~ining rxtracts heh;\vctl cx;~ctly as sl1own for mock-infected P ~ I K I C I S .
Full-length DCI protein (36
kD),
1)<2 protein ( ( i f kD) and Drosophila type I regulatory (RI) subunit (.SOkD)
were expressed at high Icvcls i n bacteria and i n Sf 21 insect cells infected with rccomhinant haculovinls (Fig- ure 1). Rot11 sources gave active
RI
protein, b u t DCl and DC'L proteins were only soluhlc whcn synthc~sized in insect cells. Protein kinase activity inSf
21 cell Iyatcs was measured using either histone H2R or a scven rcsi- due peptide. Kemptide, that serves as an escellc~nt s u b strate forPKA
(KIM' ~t nl. 19'77). Control extracts and DClcontaining extracts only sho\vcd signific;lnt kinase activity when CAMP ( 1 0 pIM) was addctl, prc~sl1m;Ihlydue to activation of the endogenous P R \ holocllzymc (Figure 2). Rv contrast, kinase activity i n I)O!con- taining extracts w a s extremely high ;Ind incrcascrl only
marginallv upon addition o f C A M I' (Figurc 2). \\'cstcrn
blots showed that the amount of t h c ;\pparc*nt Sf 21 homologue of the major 1~rosophil;l PK.A catalytic s u k unit, DCO, was not increased by expression of DC2 (Fig- ure 1). M'e therefore attrilmtc the IOkfold increase in kinase activity in extracts o f D(2cxprcssing cclls tli-
rectlv to the DC2 protc-in.
T h e specific activity of DC2 protein was
compared
with that of purified porcine
PIG\,
i n thc- prcscncc of control extract, over a rangc o f suhstratc (Kcmptitlc) concentrations. The apparent K,,, for DC2 \\.;IS - 1 0 -fold greater than that of mammalian PKA for Kcmptidc (Table I ) . However, the cstim;ltctl n1axim;d spccific ;IC-
1510 A. Melendez, W. Li and D. Kalderon
TABLE 1
Comparison of DC2, DCO and mammalian PKA specificity for substrate and inhibitor
K , (Kemptide) V,, ICm (PM) Protein ( P M ) (nmol/min/pg) (nM)
DCO
-
- 20DC2 375 1.7 200
DC2t 300 1.1 -
Mammalian
PKA 30 3.2 25
Protein kinase activities of purified mammalian PKA cata- lytic subunit supplemented with Sf 21 cell extract and of Sf 21 cell extracts after infection with baculovirus vectors for expression of DC2 and DC2t were measured at a variety of Kemptide concentrations to give an approximate K,,, the con- centration of Kemptide that gave half-maximal kinase activity, and an approximate V,,, the asymptotic maximum kinase activity. DC2t is a derivative of DC2 that lacks amino acids 59-
120. The concentration of PIU(5-24) peptide that inhibited kinase activity by 50% (ICSO) was then determined using Kemptide concentrations of 0.3 mM (DC2) or 0.1 mM (DCO and mammalian PKA). DCO activity was measured in adult head extracts, where it contributes all measurable PKAactivity
(W. LI, unpublished data).
size (67 kD), it appears that full-length DC2 protein is constitutively active. Because DC2 contains an addi- tional 150 amino acids at its amino terminus relative to DCO and the wellcharacterized yeast and mammalian 40-kD PKA isozymes, we suspected that this additional domain might be inhibitory as in the related protein kinases, PKC and myosin light-chain kinase (KFLMP et al.
1994). However, a DC2 derivative lacking amino acids 59-120 showed very similar activity and substrate affin- ity to the full-length protein (Table 1).
The peptide PKI (5-24), derived from a high affinity heat-stable mammalian inhibitor of PKA, has served as a specific probe for the active site of PKA from many organisms (WALSH et al. 1971; SCOTT et al. 1985; CHENG
et al. 1986 GLASS et al. 1992). We compared the inhibi- tory potency of PKI (5-24) toward mammalian PKA, Drosophila head extracts (where DCO contributes virtu- ally all PKA activity) and DC2-containing Sf 21 cell ex- tracts, in each case using Kemptide substrate at a con- centration close to the appropriate K ,
.
Although more PKI (5-24) was required to inhibit DC2 (IC50 -200 nM) than mammalian PKA catalytic subunit IC50 -20 nM), the inhibitory potency of PKI (5-24) toward DC2 was far greater than that reported for the nearest known relative of PKA, the cGMP-dependent protein kinase(GLASS et al. 1992).
DC2 activity can be regulated by
RI
and CAMP:Al-
though constitutive activity, a preference for Kemptide as a substrate and potent inhibition by PKI 15-24), are hallmarks of a PKA catalytic subunit, their defining feature is a high-affinity inhibition by regulatory subunit that can be reversed by the addition of CAMP. Drosoph- ila R1 protein purified from Eschm’chia coli was able to
inhibit the kinase activity of DC2containing Sf 21 ex- tracts almost as efficiently as it inhibited porcine PKA activity; 50% inhibition was achieved at roughly equimo- lar concentrations of DC2 and RI proteins (Figure 3, A and B). Addition of 10 p M CAMP completely reversed the inhibitory activity of
RI
at all concentrations tested (data not shown).DC2 could also be converted to a cAMPdependent enzyme by co-infection of Sf 21 cells with recombinant baculoviruses expressing DC2 and
RI
protein, respec- tively. Again, roughly equimolar concentrations of RIand DC2 converted about half of the DC2 kinase to a cAMP-dependent form (Figure 3C). Thus, DC2 has all the properties expected of a PKA catalytic subunit.
DC2 is expressed in mesodermal anlagen prior to gastrulation: It was previously shown that a 3.0-kb tran- script of the DC2 gene can be detected by Northern blot of RNAs from embryos, larvae, pupae and adults, whereas a 2.4kb transcript was seen only in pupae and adults. Comparison of genomic sequences with those of cDNAs representing each transcript showed that the 3.0- and 2.4kb RNA each has a unique 5’ exon that is spliced to a common acceptor (Figure 4). Thus, each transcript is under separate promoter control but en- codes the same protein. Embryonic RNA (3.0 kb) was most abundant at 2-4 hr after egg laying, and no mater- nal DC2 RNA was detected by Northern blot (not shown). These observations were corroborated by in
situ RNA localization in embryos using probes derived
from the 5’ and 3’ untranslated regions of the 3.0-kb
DC2 cDNA. These two probes gave identical results and showed no cross-reactivity with other genes in genomic Southern blots, even at low stringency. DC2 transcripts were first detected in ventral regions immediately be- fore cell cycle 14 (Figure 5A). DC2 transcripts spanned the entire length of the embryo but did not wrap around the poles and at the cellular blastoderm ex- tended laterally an average of -17-20 cell diameters from the ventral midline. This expression domain in- cludes all mesodermal anlagen and differs only margin- ally from that of the twist gene, which is essential for normal gastrulation and mesoderm specification (THISSE et al. 1987; LEPTIN and GRUNEWALD 1990; LEP-
TIN 1991). The continuous band of DC2 expression re- solved into seven broad stripes plus a cephalic region during cellularization (Figure 5C). These stripes disap peared at the beginning of gastrulation, leaving only a single region of expression in the region of the cephalic mesoderm (Figure 5D).
Early ventral expression of DC2 depends on dorsal but not twist or snail activity Embryos from mothers lacking dorsal function (Zn(2L)dp/Df(2L) 11 9) showed no DC2 RNA staining (data not shown), indicating that dorsal protein is required directly or indirectly for DC2
I)<:!?: A I)rosophil;~ I%\ Isozynw 1.51 1
20
-
o ! 1 I I I
0 IO0 200 300 400
[R11
nM
B
120-100 .(
-
C . I i0 IO0 300 200 400
[MI
nM
A
B
CD
R I : D C 2 0 0.75 2.3 5.3
FKX'RE S.-DC2 kinase activity can be inhibited by Drtr sophila type I regrhtory s u b ~ ~ n i r . Drosophila RI. purifird from k,*. coli, was prcincrtbatetl with ( A ) I)<;!? bacr~lovirr~s-in- fcctcd cell extracts or
(B)
mork-infcctc*rl extracts sr~pple- mented with pwified manlmalian PILI catalytic subunit for 2 hr on ice before kinase assays. DC!? protrin ( " x 0 1111) ant! mammalian I'KA ( " 1 0 nM) concentrations in tllcse rxt~:~cts wcrc cstimatctl by Western hlot and <:oornassir staining. (C;)Sf 21 cells w r c ro-infected with tliffercnt proportions of RI and DC!? recombinant hacllloviruses t o give C ~ I I ' P C I S with csti-
matctl RI:DC!? protein ratios ;IS shown. The kinase activity of cach cx1rac-1 w m mcasrtrctl in t h r prcscncc o r abscncc of
REKC;:\ U/ / I / . 1991; Kos\~..\s Pt / I / . 1 9 9 1 ; I.EII'IS 1991;
RIITIK r t / I / . 1993). Rv contrast. /IC2 R S X cxprcssion
\viis normal in almost ;dl prcgastrulation e m h T o s col- lecrctl from parents heterozygous for t h c
n u l l
troista l -
IeIc, .TN/I'"'~'' (2O<)/zI6 = W % ) , or thc t I o u t d c mutant CIlromosomc .snn""rf'/?r(2/~~tltJi\""
( 3 i i / M 3
=W h ) .
If there were an efrect o f 170; or snn m~tt;~tions. we w o u l d h;lw expcctcd 2.5% o f the embyos t o show abnormal o r no /IC2 expression.DC2 reporter genes are also expressed in developing
optic lobes and imaginal discs: X 3..5-kb /)C2 promotcr elcmcnt ilnmcdiatcly npstream of the transcription i n i - tiation site for t h c 3.0-kb /IC2 mRSA was fuscd to the
t,'.
roli /or% gene ;md tr;lnsformctl into the Drosophila germline. / n s i t u hybridizatiotl t o RNA using a InrZ D S A probc antl antihotly directed t o ~ a r d PR;Il;Ictositl;1s<~ both showed a ventral accumulation of transcripts bv-fore gastrulation ;IS for the endogcmous /)(,'2 R S A (Fig- urc 5E). ~-Galactosidasc protein persisted longer and could he seen clcarly i n the mesorlermal layer of the
cxtcntling germ band and in an anterior region proba- bly corresponding to cephalic mesoderm (Figure .5F). In third instar lal-\.;\e, the most prominent sites of ex-
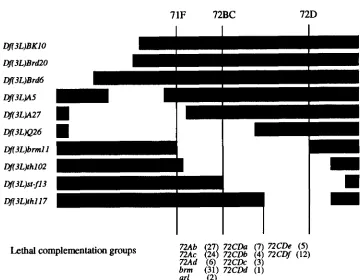
pression of the / K 2 reporter gene \vert in a crescent of optic lobe lamina cells surrounding the stalk of the optic nerve (Figure 6A). central regions of the leg imag- inal discs (Figure 6C ) and dorsal regions of the wing imaginal disc that develop into the notum (Figure 6D). Isolation of DC2 mutants: We mapped the /IC2 tran- scription unit to a 20-kb region that was absent in / > p ( 3 / . ) / m n l / , disrupted in Qf(3/#/1/02 (Figurc 4), hut intact in Qf(31Jst- f13 and /)/(3/#/1/17 (Figure i ) . The distal breakpoints of Qf(31,)lm1I / antl /?ji3/,).s/-f13 are separated
by
60 k h of DSA, including t h c /)(;2transcrip- tion unit (Figure 4). M'e used a stantlard EMS F'L lethal scheme to identify recessive lethals uncovered by Qp(3/.)/mnl I and found 11 complementation groups, fiw of which complemented /?f(3/.) .s/,fl3 and thcrefore lay within the 60-kh /)C'2-cont;\ining interval. Two of these complementation groups corresponded to the previously identified genes, hmhmn (T..WKYS P/ nl. 1992) and n@w (TxstK1.s PI nl. 1991). A /)(;2transgerleconsisting of thc X i k h DC2 promoter defined above. a full-length 1)C2 cDNA and an S\'40 poly(A) addition signal failed t o rescue any members of the three rc- maining complcmcnt;~tion groups even with respect to lethal phase (72Ab is cmblynic lethal; i!!:\c antl 72Ad arc l a n d lethal). Also, n o coding region changes i n / X 2 were found in PCR amplific;ltion products from representative alleles o f cach complementation group. Becat~se we isolated an average of 18 alleles per lethal
kk.
/?f(2!<)/7tIi'"" (464/484 = !I(;%), the stroilg .TtttIi/ :ll-1512 A. Slc*li.ntlr/.. 14'. 1.i ;In(! I). k d t l r r o n
TELOMERE CENTROMERE
Dfl3L) fhl02
I ...I
Dfl3L)bmll
I
...4
I t
Dfl3LY27l i i i f
Dc2 3.0 kb
i i R A 3 5
m K N h
...
-:
'ri'
:"::li'y'
rn
-.
i.".
iu
5ii
*
'"..
'
A
\
Dc22.4 kb
mRNA
3 5 ;
5 :
t i I\ i i :: a 1
-
kb& ~ R I I 12.411.81 3.0 7.4 I 2.9 I 10 I 5.8 I 8. I I 8.5 I12
mol 4.0 11 4.4 I 21 I 9.5 I 8.8 I 6.5 11.911 17.0
BumHI 2O I 2.01 7.0 1 3.0 I 7.8 1 1.51 3.4 I I 2.9 I 4.5 I 5.0 I
Sal1 27 I 3.5 I 5.8 I 4.6 I 6.8 I 4.0 1 25
Hind 111 1 3.5 I 8.6 11.61 5.5 I 9.0 1 2.4 I 5.4 I I 3.8 I 1.8 I 6.0 I 4.3
DC2 Promoter Fragment
17771
A
-C m O B H E 6 5 5
PROBE!? R II.
FI(;I.KI< 4.-Sk)lccular map o f t h c /IC2 gcnomic rrgion. Breakpoints were tnappcd I)? Sor~~hcrn blots ;IS i n Figwc 8 rc*lative
t o il ~h~O~lOSOnlil! Wa!k i n Iiun!)d;I phage. n c distil! I)rcilkpoint o f /?/(3/.)/h 102 lics !)ctwern t w o f h t t k f 1 sites (;IS indicated by
the dotted line: solid line rrprcwnts DSX t h a t is present). t h a t o f / ? / f Y , ) / m t I l lies l w t w r e n tlw int1ic;ltctl Shol inltl ItmttHI sites
illltl t h c proxinlitl I)rc*;tkpoint 01' I)/f3/.),427 lies hctwen the intlic;ttctl S d i m t l MntnHI sit(-s. l l w 3.0- ;In<! !!:l-kI> IIIKS;\S dilkr only i n t h c - 3' eson. Tlw Ir.;lnslation i n i t i a t i o n codon (XU(;) is i n the conlnlon second cson. I n w o n junctions IFCTC- tl~wrminctl
by comparison o f grnonlic ant1 cl)S;\ s q t w n c c s . Introns inwrrupt thc. pul)lishrtl /)(:2 cl)S;\ srqlwnce (K\I.DEKOX ; 1 n d K ~ . I ~ I s
I!)XX) d t e r nuclroritlc*s X I S , 1 4 W . I(i(il, 1 7 1 , 201 7, 2I!N and 2 2 i i . ' I - 1 1 ~ /X:2 promoter Irilgnwnl rstrntls l w n l ; ~ n EmKI sile t o a syntlwtic K p l sitc 123 bp ;1f1cr t l w :i' c m t l o f ' t h c 3.O-kh cDSA. Prol)cs ;\ (gcnomic i:'roKI i.4 kb), I\ ( / X 2 cI)S;\ / h t d ll-.Sml
581 bp) ilntl ( 1 f / N 2 3 . 0 kl) rl)SA) w r ( * rlscd i n Sotlthcrn l h t s o f Figure X.
complcmc.nt;ltion group, we considered it likcly a t this point t h a t the 1)C2 gcwc. \vas n o t csscntiill for viability. Rccallsc the distal hrcakpoint of' 1)fl3~.)hnnI~ lies im- mediately ;lclj;lccnt t o the 3' end of the 1)C2 gene. t w
attempted t o isolatc. a n w tlcficimcy that w o r l l t l corn-
recovered 22.5 whitc-cycd 1Iic.s from y-ny mlltagcnesis of 21 chromosome- carying a I ~ w ' ] transposon (M;k,' I -
151) at flF..jrlst distal t o thc 1)C2 locus. Onc of these mutagcnizctl cllroI~losoI~l(~s, l?ff31,),42i, complcmcntctl I?f(3I,)/mnl I h r t t not t h c morc distal tlcficicncy,
most of the 11C2 coding scqtlcncc (Figurcs 4 and 8).
Animals /m~/shctcroirygo~~s for Dfl31,)/\2i ant1 I?fl31,)- Imnl I lacked a l l scqwnc-cs o f the IX2 kinasr tlomain antl indectl c x p r c ~ ~ t l no I X 2 protcin ( d a t a not shown) but w'crc viahlc and fertile. I n particular, sections o f early cmh1y)s staincd w i t h / n k / antihotly showed t h a t
gastrrhtion \\";IS norm;d and no tlefc~ts w r c ohsc*twtl i n the cxtcrnal morphology o f ;rdrtlts o r i n sc-ctions o f plcmcnr Dff31,)/~11r I I for ;dl gcllcs o t h c r th;ln 1)0*2. \Ye
1?r(x)q2h, and Iv:ls ~ o r l l l ( ~ I)!
s o ~ l t ~ ~ c ~ n
I)lot t o tlclctcadlllt C ~ V S ;Inti optic- lohcs (A. (;.,\.\IIY)s, p<*rson;ll com-
mrtnication) that might relate t o the normal sites o f 1)C2 cxprcssion.
Reductions in DCO activity do not reveal latent DC2 functions: Rccausc 1)(,*2 a n t l I)(;(/ encode similar hio-
chcmicid fhctions, we investigated whathcr the* two ki- llilscs might share overlapping frlnctions during em- hlyogcwcsis. \4atcrn;d DCO expression persists at high levels for thc first Imlf o f cmhrygenesis and appears stlflicicnt for c.mh-yogc-nt~sis. ;IS animals zygotically n u l l
for I ) U / die ;IS normally patterned first instar 1an.ae
(L\SI;. m t l K.\I.I)KROS l!W3). 14'~ thcrefore tcastcd wl1cthc.r the partially pcwctrant mat(ma1 cfkct Icth;dit\. due t o hypomorphic I X ' O ;~llc*lcs w:rs enhanccd by lack of zygotic DC2. 'Thcrc was no significant altctxtion i n hatching rates hctwen anim;tls lacking 1)(:2 (Dff31,)A2i/ !)fl3I-)/m// I ) ; 1 n d controls ciu-rying onc fmctional copy o f ' the II(J.2 ~ C I W (l?fl31~)~\~/I?fl31,)/~171/~ I ) (Tddc %\).
Similarly. h;ltching of animals zygotically n u l l for I ) ( N WAS not rcctaccd
by
thc absence of 1)(,'2 rclativc t o the 1X2 Iwtcw)zygous controls (Tahlc 2R).DC2: A DrosoDhila PKA Isozvme 1513
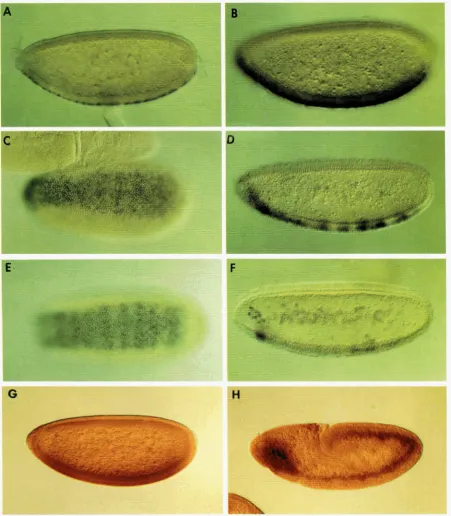
FIGURE 5.--DC2 l,,, exkLasion embryos. (A-F) wild-type embryos after in situ hyullciization with digv,l~,,,;,,,beled DC2 probes from 5' and 3' untranslated regions. G and H show embryos carrying a DC2-hcZ transgene stained with antibody to p-galactosidase. All embryos are oriented with anterior to the left and ventral down or (C and E) facing the observer. (A) DC2 is first expressed at the syncytial blastoderm stage, as early as cell cycle 13. (B) Maximum expression is seen at cellular blastoderm. At this stage, expression is restricted to a 17-20 cell width on the most ventral surface of the embryo (C). The pattern resolves into pair rule stripes (D and E) and largely disappears by the beginning of ventral furrow formation, when RNA is seen only in the anterior cephalic region ( F ) . DC2 promoter fusions show ventral accumulation of p-galactosidase before cellularization as for the endogenous DC2 RNA (G). Staining is seen in the mesodermal layer during germ band extension (H).
of adult legs, wings and other disc derivatives without P[TgDC&DC2] transgene does not efficiently rescue affecting cell viability (reviewed in KALDERON 1995). DCO mutant phenotypes: A transgene consisting of ge- Large DCO mutant clones could still be recovered in nomic DNA from
2.8
kb upstream of the transcription animals lacking DC2activity and produced avery similar start site to the termination codon of the DCO gene array of phenotypes regardless of DC2 genotype (not (TgDCO-DCO) produces DCO protein at 20% the level1514 A. MelCndez, W. Li and D. Kalderon
FIGURE 6.-Expression of DC2 reporter genes in third in- star larvae. Brain-eye disc complexes (anterior to the left) of third instar larvae carrying the DCZ-lac2 reporter gene were double-stained with antibody to /3galactosidase and a neu- ronal-specific antibody, anti-HRF'. (A) DC2-lacZ expression (brown) in the crescent-shaped lamina anlagen of the optic lobe surrounding the optic nerve (green in B). The DC2 re- porter gene was also expressed in central regions of leg discs (C) and dorsal regions of wing discs that give rise to part of the adult notum (D)
.
functions
(LANE
and KALDERON 1993). We thereforeused the same DCO promoter to drive DC2 expression by exchanging DCO and DC2 cDNA sequences (to give TgDCO-DC2) to test whether DC2 protein could substi- tute functionally for DCO protein. Western blots of
adults carrying the TgDCO-DC2 transgene showed ele- vated levels of full-length DC2 protein, roughly equiva- lent to the levels of DCO protein encoded by TgDCO- DCO (not shown). However, only the TgDCO-DCO transgene was able to rescue a variety of DCO functions efficiently.
A single TgDCO-DCO transgene
fully
rescues the viabil- ity of hemizygous weak DCO alleles (such as Oca"3) and partially rescues the viability of null DCO alleles (such asDCf')
.
By contrast, chromosomes carrying the TgDCO- DC2 transgene showed no rescue of DC@'3/DCf2 ani- mals and only a small increase in viability for the par- tially viable DCO allele combination, Df2L)Tw2/DC@'(Figure 9). Also, the female sterility and minimal egg laying associated with this genotype was unaffected by two copies of the TgDCO-DC2 transgene but fully res- cued by one copy of TgDCO-DCO. Similarly, the adult phenotnj'es induced by DCO null mutant clones were fully rescued by one copy of TgDCO-DCO but were unaf- fected by two copies of TgDCO-DC2 (not shown).
The expression of DC2 from the TgDCO-DC2 trans gene had no clear deleterious effects. Although several insertions caused a mild rough eye phenotype, abnor- mally constructed ommatidia have only been observed in males canying an X-linked insertion of the transgene.
DISCUSSION
We show that the Drosophila DC2 protein has the defining biochemical properties of a PKA catalytic sub- unit. It was constitutively active in the absence of regula- tory subunit and efficient inhibition by regulatory sub- unit was relieved by the addition of CAMP. DC2 also shares with mammalian PKA catalytic subunit, C,, a preference for Kemptide as substrate and susceptibility to inhibition by low concentratioqs of PKI.
The biochemical properties of DC2 can be rational- ized according to the roles of specific amino acids in the catalytic subunit that make contact with substrate, regulatory subunit and PKI. Both mutational studies
(GIBBS et al. 1988; TOMODA et al. 1993) and the crystal structure of mouse C, bound to PKI peptides
DC2: A Drosophila PKA Isozyme 1515
71F
72BC
72D
Df(3L)BKIO
Df(3L)BrdZO
m3L)Brd6
Df(3L145
-
Df(3LH226
I
Df(3LM27
Df(3L)bnnl I
I
Df(3L)rhlOZ
DJT3Lht-fl3
DJT3L)hl I7
I
I
I
Lethal
complementation
groups 7 2 ~ b 72Ac 1241 27 72cDa 72CDb (7) (4) 72CDe 72CDf (12) ( 5 )brm 72Ad ( 6 ) 72CDc (3) (31) 72CDd (1)
arl (2)
FIGURE 7.-Genetic map of 71F-72D region. The empty spaces represent DNA missing from each deficiency chromosome. The relative order of the distal breakpoints of D f 3 L ) h l Z , th102, s t f l 3 and t h l l 7 were determined by molecular mapping and are consistent with complementation studies with lethal point mutations in the region. The published cytology for Df(3L)brmll
and sty13 is shown. The listed cytology for Df(3L)thllZ rather than Df(3L)thlOZ is appropriate for Df(3L)thl02 according to our map and Df(3L))thll7is probably more complex than listed in LINDSLEY and ZIMM (1992). The proximal breakpoint of Df(3LjA27
has also been mapped molecularly, but other breakpoints are drawn only according to complementation properties. Thus,
Df(3L)A5 complements Df(3L)Brdb and Brdl8 but not Df(3L)BrdZO or Df(3L)BKlO, whereas Df(3L)Q26 and Df(3L)A27 fail to complement all four deficiencies. The five complementation groups between Df(3L)brmll and stf13 distal breakpoints have been described previously (BRIZUELA et al. 1994); the remaining six are designated 72CDa-f according to their approximate cytological location. The number of alleles isolated for each complementation group is in parentheses.
(TPK1-3), which are not as readily inhibited by PKI
(GLASS et al. 1992). Mutational studies have also identi- fied residues (H-87, [K-1891, W-196, L-198, [K-2131, [K-
2171)
that contribute specifically to interaction with regulatory subunit (GIBBS et al. 1988; LEVIN et al. 1988;ORELLANA et al. 1993). All of these residues are con- served in DC2, DCO and yeast TPK3, whereas at least two are altered in cGMP-dependent kinases and in Dro- sophila DC1 (Figure 10). Thus, although the major Dro- sophila PKA catalytic subunit, DCO, is identical to mouse C, in all of the residues thought to be critical for binding substrates and inhibitors, DC2 shares most of these residues and has similar but distinguishable biochemical properties.
Of the PKA catalytic subunits shown in Figure 10, only DC2 has a substantial amino-terminal region preceding the kinase domain. A PKA catalytic subunit in Dictyostel- ium is of similar size and structure to DC2, but the amino termini of these two proteins show no direct sequence similarity (MANN et al. 1992; ANJARD et al. 1993). Our in vitro experiments using a peptide substrate did not reveal a major difference between the activities of full-length DC2 protein and a derivative lacking half of the unique amino-terminal domain. It may be that this domain limits
the specificity of DC2 toward protein substrates by steric hindrance or that other proteins can bind this domain
in uiuo and thereby provide additional regulatory input. However, this domain does not contain an obvious pseu- dosubstrate site and does not appear to be the primary regulator of DC2 activity. The amino-terminus of DC2 might influence its subcellular distribution. The in- creased size of DC2 would be expected to reduce the efficiency of passive diffusion of monomeric catalytic sub- unit into the nucleus relative to the prototypical 40-kD catalytic subunits (PAINE and FELDHERR 1972; MEINKOTH et al. 1990; BACSKAI et al. 1993). Also, the extreme amino- terminus of DC2 lacks the consensus sequence for myris- toylation that is present on mouse C, and Drosophila DCO but absent from mouse Cp and the catalytic subunits of
S.
cerevisiae, Caenorhabditis elegans and Dictyostelium(CARR et al. 1982; TODA et al. 1987b; GROSS et al. 1990; M A N N and FIRTEL 1991).
1516 A. Meli.ndez, W. 1.i and D. Kaltlcron
A It
c
1)R
2 2 =?
2
4.
$ 2 2
g s
-
-41
s s $
< e
5 2 $ 2E
s a
s s s
u)u)
..."
- 4
k
7
2 ;
2E 2E -2"
41 c
2 2
9 9
2 2 2
9 9 - ? ? 3 .. -1 .. -?-1
e
7.2
-
7d)-5.x
-
7.4-
7.0-
3.0-
2.0-
K H s s s I{ I( I{ I{
s
K I Ip r t k A p r o l r A 1wlw 13 prolr ( * pnllo ('
FI(;t!KE 8.-Mapping the Df(3lJlnmll distal and Df(31JA27 proximal breakpoints. ( A - C ) Southern blot analysis of DNA
from flies carrying 1 ) / 3 1 ~ ) t m n l l in frnns to two different balancer chromosomes that show useful restriction fragment length polymorphisms. Restriction enzymes used were KmRI
(R),
XlroI (X), NnmHI ( R ) ,.%dl
(S) and I-lit~dlll (14). (A) llf~3l~)tmnll has an I h R I fragment of .i.X k b instead of the TM? 7.0 kh and the MKRU 7.5 kh (7.4 kh in Figure 4). (R) Ilf(?l@ml I has a 4.0-kh Xhol fragment instead of 9.9kh (TM3 and Figure 4) or 10.0 kh ( I M K R Y ) . ( C ) the IhmHI 7.0-kh fragment (XM3 and Figure 4:6 . 0 kh in M U S ) is missing in Df(31,)lmnlI. Thus. the Df(3L)tmnll breakpoint lies within the Xhol ! G k b fragment, not more
than 4.0 kh away from the 2.1-kh M o l fragment (Figure 4). DNA from fliescar~ingDf(3I,)A5or IIf(31JA2iin /rnm to 1~fl31~)lmn11 was tligcsted and hybridized t o a 1lC2 cDNA prohc ( C in Figure 4). D/(3l,)lnml I lacks all DC2 cDNA sequences so the DNA that hybridizes corrcsponds only t o the chromosome in trans t o Dfl3lJtmnll. (D) Df(31,)A27 lacks RnmHl fragment5 of 7.0 and 2.0 kh hut retains 3.0- and 5.4kh fragments. (E:) a Snfi fragment o f 3.5 kh and KmRI fragments of 10.3, 7.4 and 2.9 kh are mising; a new I h R I hand of 8.0 kh and a new Hind111 band o f 17 kh were detected. whereas the Hindlll hands of 9.0 and 5.5 kh are missing. \'\'e therefore placed the llff3l.)A2i breakpoint immediatelv distal to the I h t H I 3.0-kh fragment (Figure 4).
have been examined, with particularly high levels in nenfous tissue (SKOUIAKIS Pt nl. 1993;
LWE
and KALD- EKON 1995). By contrast, DC2 expression in embryos was purely zygotic and restricted to mesodermal pro- genitors for a short period of time around cellular Mas- toderm. Postembryonic DC2 expression, inferred from the use of reporter genes, was also highly restricted. In third instar larvae, prominent expression was confinedto a subset of optic lobe lamina cells and to a subpopula- tion of leg and wing imaginal disc cells. Furthermore, the sum total of DC2 protein expressed in adults was estimated to be no more than one tenth of the level of
DCO protein (data not shown).
T h e restricted expression of DC2 led u s t o question
it5 significance. In cells that express f X 2 , are there func-
tions that require the concerted action of DCOand DC2 or the function of DC2 alone? Would ectopic DC2 ex- pression interfere with
PKA
signaling pathways in cells that normally express only f X O ?DC2 is not an essential gene: T h e most striking aspect
of animals that completely lack DC2 activity is that they appear to develop normally, producing fertile adults of normal outward appearance. This observation was
unexpected given that DC2 encodes a biochemically active
PKA
catalytic subunit and that the more abun- dantly expressed catalytic subunit, D C O , has multiple functions during development & W E and ~ \ I , D E R O X1995; 1.1 Pt nl. 1995).
I t is clear that fX2activity is not essential for viability. Flies can be propagated efficiently for many generations in the complete absence of the coding region for the DC2 kinase domain. Also, we have searched extensively by nucleic acid hybridization and PCR for a gene closely related to DC2 but have consistently confirmed the DCO and D C I genes as the closest relatives of 1 X 2 i n the Drosophila genome.
T h e possibilities that DC2 performs a subtle nones- sential developmental function o r that it shares overlap ping functions with the biochemically related DCO pro- tein are not supported by any of our obsenations to
DC2: A Drosophila PKA Isozyme 1517
TABLE 2
Effect of DC2 activity on hatching of embryos
with reduced DCO function
% Expected hatching Zygotic chromosome I11
genotype Df(3L)A27/ Df(3L)AS/
D f ( 3 L ) h l l D f ( 3 L ) h l l
A. Maternal genotype chromosome I1
DCOE'"/Df(2L)Tw2 18" 92 88
DCOE'o/Df(2L)Tw2 25" 76 72
DCOAI3/Df(2L)Tw2 25" 80 88
% Expected hatching of DCO zygotic null larvae B. Zygotic genotype chromosome I11
Df( 3L)A 27 /D f (3 L)h l I 9.4
D f ( 3 L ) A S / D f ( 3 L ) h l l 9.2
Df(3L)A27/+ 67
Df(3L)A5/
+
75+ / D f ( 3 L ) h l l 44
In A, females heterozygous for P[y+] TM3 and DJ(3LlA27 or Df(3L)AS and additionally heterozy ous for the DCO alleles, Df(2L)Tw2 and DCO"" or DCO"', were crossed to
D f ( 3 L ) h l l/ P [ y ' ] males. The effect of chromosome ZIIgeno- type on hatching was measured by counting the number of
y-
[Df(3L)A27 or Df(3L)AS/Df(3L)brmll] and y + larvae that hatched. Hatching rates of y - larvae were multiplied by three and expressed as a percentage of y + larvae hatching rates. Overall hatching rates were 54% (line l ) , 72% (line 2) and 63% (line 3 ) . In B, crosses were made between yuP; DCf12/ p[y+]; Df(3L)A27 or AS/P[y'] TM3 females and yuP, DC@;'/ P [ y + ] ; D f ( 3 L ) h l l / P [ y + ] males and hatching larvae were counted. The number of y- larvae was compared to the num- ber expected from counting y + siblings and assuming 100% hatching efficiency for the latter. Each deficiency chromo- some, but particularly Df(3L)brmll, reduced hatching so that hatching is very low in deficiency t-lanpheterozygotes but no significant difference is seen between the presence [Df(3L)- A S / D f ( 3 L ) h l l ] and absence [Df(3L)A27/Df(3L)brmll] of a DC2 gene.to DC2. Because DCO is required for oogenesis, we could not examine the effect of DC2 mutations in the com- plete absence of DCO activity, so it remains possible that the two proteins share some functions during em-
bryogenesis.
Reductions of DCO activity in imaginal discs lead to the ectopic induction of hedgehogresponsive genes such as decapentaplegic ( d p p ) and consequent alterations in the growth and patterning of imaginal tissues, including partial leg and wing duplications. Elimination of DC2
alone did not affect the morphology of imaginal disc derivatives nor did the absence of DC2 affect the range of adult phenotypes induced by DCO null clones. In particular, clones encompassing >20 tarsal bristles were frequently recovered, indicating that cells were viable and able to grow in the absence of functional genes for both of the known Drosophila PKA catalytic subunits.
Z v I A B I r n
50
40
30
20
10
0 1
copies of
011 chromosome
TgDCO-DC2
1 1 0201 1 0 0
1 0 1001 0 1 0
"
MALES FEMALES
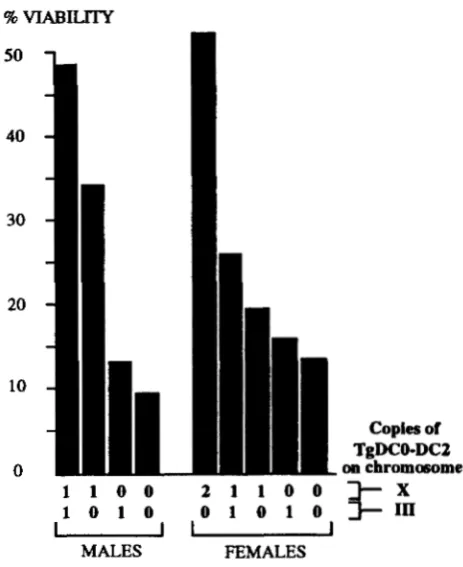
FIGURE 9. -Effect of DCO-DC2 transgene on viability of DCO hypomorphs. The percentage viability of animals heterozy- gous for the hypomorphic DCO allele, Df(2L)Tw2 and the null allele, D C V 2 was calculated by counting adults of this geno- type relative to siblings carrying the C y 0 balancer. Rescue by a DCO-DC2 transgene on the Xchromosome (X) or the third chromosome (IZa was assessed by comparing the effects of these chromosomes relative to a wild-type X chromosome or a yw" X chromosome and a wild-type third chromosome. Re- sults are expressed as averages from multiple experiments, including reciprocal crosses and show that a chromosome carrying a DCO-DC2 transgene consistently results in a small increase in viability relative to control chromosomes. Because the viability of this DCO genotype is affected considerably by genetic background, we cannot be sure from these studies that the DCO-DC2 transgene itself is increasing viability.
Functional overlap between DCO and DC2 proteins: DC2 was expressed from a DCO promoter fragment to ask whether DC2 protein could substitute for DCO and whether it is important to restrict DC2 expression. No
clear detrimental effects of expressing either DCO o r
DC2 from a DCO promoter were observed. By contrast, we know that ubiquitous expression of low levels (-5%
of endogenous PKA) of a constitutively active mouse
PKA catalytic subunit with minimal affinity for regula- tory subunit causes pupal lethality (W. LI and D. KALD- ERON, unpublished observations). T h e estimated 20%
1518 A. MelCndez, W. Li and D. Kalderon
10
F I A S G R T G + $ N @ I H D
u
r
‘
2351
236 239A
127 329 (331)-
170 230E D E
E D E
E D E
E O E
E O O
E m E l
mCa$
hC 7
DCO DCl Dc2 TPKl
hGKI
DGKI
189
K K K K K K K K
1% 198 213 217
W L K K
W L K K
G L B K
L K K
W L K K
O L K K
W m K m
W m K a
PKI AND SUBSTRATE BINDING REGULATORY SUBUNIT BINDING
FIGURE 10.-Amino acid residues critical for binding Kemptide, PKI and regulatory subunit. PKI, sequence from 10 to 24 is shown together with the amino acid residues numbered according to mouse PKA C, that are important for recognition of F-10 and the two arginine residues (underlined) preceding the normal phosphorylation site (circled, A in PKI; S in Kemptide). On the right are residues from mouse C, important for recognition of regulatory subunit. Below are the corresponding residues from human PKA y catalytic subunit (hC,); the Drosophila DCO, DC1 and DC2 proteins; a yeast PKA catalytic subunit (TPK1); human cGMPdependent protein kinase Ia (hGK1) and Drosophila cGMPdependent protein kinase 1 (DGK1). Boxed residues differ from mouse C,.
are viable, suggesting either that a similar compensatory mechanism is induced by DC2 or that the DC2 protein does not recognize potentially toxic substrates as effi- ciently as mammalian PKA, or both.
The functions of the endogenous DCO gene in the female germline, imaginal discs and in supporting via- bility through larval and pupal stages can be replaced by a DCO transgene that encodes roughly
20%
of the activity of the wild-type gene. DC2 protein expressed from the same DCO transgene promoter could not sub- stitute for any of these functions in the absence of DCOactivity and showed only a small enhancement of the viability of animals with reduced DCO activity. The latter effect could result from the phosphorylation of DCO substrates by DC2 or, alternatively, from displacement and hence activation of DCO from complexes with regu- latory subunit. The failure of DC2 to complement the imaginal disc phenotype of DCO mutations could be due to insufficient levels of DC2 expression. However, we estimate that the level of ectopic DC2 from the DCO- DC2 transgene is roughly 10% that of endogenous DCO, whereas null DCO mutant clones in imaginal discs are fully rescued by expression of activated mouse catalytic subunit at levels <5% of normal endogenous DCO (LI et al., 1995). We therefore favor the explanation that, at least in this instance, DC2 cannot recognize DCO substrates efficiently. This would be consistent with the differences in affinity for the peptide substrate, Kemp- tide, that we have observed in vitro.
PKA
isozymes in Drosophila: One objective of our studies has been to define the molecular components of Drosophila PKA isozymes. The 40-kD DCO protein is a typical PKA catalytic subunit. It is of similar size to mouse C, , >80% of amino acid residues are identical between the two sequences, including those crucial for recogni- tion of substrate, PKI and regulatory subunit, it accountsfor the majority of measurable PKA activity in flies and can be substituted, at least in imaginal discs, by mouse C,. The DCI and DC2 genes of Drosophila do not have such obvious counterparts in other organisms. The DC1 and DC2 proteins show only limited sequence similarity to DCO (<50% amino acid identity) compared with the similarities among the known yeast (>75% identity) and mammalian catalytic subunits (>79% identity). Our demonstration that DC2 has the biochemical properties of a catalytic subunit raises the possibility that mammals may also harbor PKA isozymes that are more divergent than currently recognized, perhaps including a specific homologue of DC2. Whether DCl also encodes a PKA catalytic subunit remains an open question. From se- quence comparisons, we would expect DC1 to recognize substrates including the
.
.RRXS.
. .
consensus but have reason to doubt that regulatory subunit would bind with high affinity (Figure 10). We were unable to detect ki- nase activity from soluble DCl expressed in Sf 21 insect cells, perhaps because Kemptide and histones were i n a p propriate substrates. Alternatively, DCl may require modification or association with an essential activator. Either of these explanations, if true, would set DC1 apart from any other known PKA isozyme.In yeast and mammalian tissue culture, a very low level of PKA activity can satisfy requirements for cell growth and viability (COFFINO et al. 1976; CAMERON et al. 1988; GIBBS et al. 1988). In Drosophila, an imaginal disc cell that lacks DC2 gene products and loses DCO