Copyright 0 1994 by the Genetics Society of America
Second Site Mutations Specifically Suppress the
Fix- Phenotype of
Rhizobium meliloti ndvF Mutations on Alfalfa: Identification
of a
Conditional ndvF-Dependent Mucoid Colony Phenotype
Ivan J. Oresnik, Trevor C. Charles
and
Turlough
M. Finan
Department of Biology, McMaster Uniuersity, West, Hamilton, Ontario L8S 4K1, Canada Manuscript received August 9, 1993
Accepted for publication December 8, 1993
ABSTRACT
Rhizobium meliloti mutants carrying nduF insertion or deletion mutations induce nodules on alfalfa which contain very few infected cells and fail to fix N, (Fix-). We have characterized five independent second site mutations (designated s f . ) which completely suppress the Fix- phenotype of nduF mutants on Medicago sativa but not on another R . meliloti host Melilotus alba. Genetic mapping and phenotypic analysis revealed that the suppressor mutations sfx-1, sfx-4 and sfx-5 mapped to a single locus which was distinct from another locus defined by the sfx-2 and sfx-3mutations. Tn5-mobmediated conjugal mapping experiments showed that the sfx-1 locus was located clockwise from trp-33 on the R . meliloti chromosome and a detailed cotransduction map of this region was generated. To clone the sfx-1 locus, we prepared a cosmid library from total DNA obtained from an sfx-1, nduF deletion strain. From this library, a cosmid pTH56, which converted Fix- nduF mutants to Fix’, was isolated. Southern blot analysis provided direct physical evidence that the insert DNA in plasmid pTH56 was contiguous with the sfx-1 region. On low osmolarity glutamate-yeast extract-mannitol-salts medium (GYM) agar medium, nduF insertion and de- letion mutants were found to have a mucoid colony phenotype, as opposed to the dry colony phenotype of the wild-type strain. This phenotype was shown to be dependent on the exoB and expE genes required for synthesis of exopolysaccharide I1 in R. meliloti but not to be dependent on genes required exclusively for the synthesis of the succinoglycan or exopolysaccharide I. Transduction of either sfx-1 or sfx-2 or transfer of the cosmid pTH56 into the nduF mutants restored them to a wild-type dry colony phenotype. The mucoid phenotype is not responsible for the Fix- phenotype of nduF mutants as the Fix-, nduF exp double mutants can be complemented to Fix’ by introducing plasmids which carry only the wild-type nduF genes.
T
HE development of N,-fixing root nodules on le- guminous plants results from a complex series of interactions between the plant and bacteria. Genetic analysis has revealed the presence of nodulation ( n o d ) genes in the bacteria which are expressed in the pres- ence of flavonoid compounds produced by the plant roots (LONG 1989). The Nod proteins synthesize an lipo- oligosaccharide signal that is excreted and stimulates meristematic activity in the root cortical cells which eventually give rise to the nodule (LEROUGE et al. 1991; TRUCHET et al. 1991; SPAINK et al. 1991). At the same time the bacteria invade the nodule via tubelike structures called infection threads. Recent evidence suggests that the pathway of infection thread growth may be deter- mined by the interaction of the lipwligosaccharide sig- nal molecule and the plant cells (VAN BRUSSEL et al. 1992). Later in nodule development the bacteria are released from the infection thread into the cytoplasm of the plant cell. At this time the nitrogen fixation genes ( n i f and f i x ) are expressed and the bacteria differen- tiate to their N,-fixing form called bacteroids.We are studying the symbiosis between the bacterium Rhizobium meliloti and the alfalfa (Medicago sativa) plant. Among R . meliloti mutants which form nodules
Genetics 136: 1233-1243 (April, 1994)
1234 I. J. Oresnik, T. C. Charles and T. M. Finan
pressed to Fix' by second-site mutations which lead to the increased synthesis of the EPS I1 exopolysaccharide (GLAZEBROOK and WALKER 1989; ZAUN et al. 1989). Ad- ditionally the Fix- phenotype of exo mutants can also be suppressed by either naturally occurring or mutant sup- pressor alleles of loci involved in lipopolysaccharide syn- thesis (PUTNOKY et al. 1990; WILLIAMS et al. 1990).
Many of the exo and eps genes are located on the sec- ond (pEXO) of two symbiotic megaplasmids of R. me- liloti (FINAN et a l . 1986; HWES et al. 1986). Employing linked transposon insertions we generated a detailed ge- netic map of the 1600 kilobase (kb) pEXO megaplasmid (CHARLES and FINAN 1990). We constructed large de- fined deletions in the plasmid and by screening strains carrylng these deletions, we identified a symbiotic locus which was designated ndvF. Strains carrying ndvF mu- tations had a nodule development phenotype ( n d v ) similar to the ndvA, ndvB and exo mutants of R. meliloti (CHARLES and FINAN 1991; CHARLES et al. 1991). The nduF locus was shown to be distinct from the previously char- acterized ndvA, ndvB and exo genes. However, no free- living phenotype was observed in nduF insertion or de- letion mutations. Insertion mutagenesis localized ndvF to a 5-kb region and the isolation of TnphoA insertions which expressed alkaline phosphatase activitywithin this region suggests that ndvF encoded proteins are either exported or span the cytoplasmic membrane (CHARLES
et nl. 1991).
Here we report on the genetic characterization and manipulation of mutations which suppress the Fix- phe- notype of nduF mutants but do not suppress the Fix- phenotype of nduA or exo mutations. The s f x alleles are shown to map to at least two different loci and we de- scribe the cloning of the sfx-l allele on the cosmid clone
pTH56. We show that the ndvF mutants exhibit a low osmolarity dependent mucoid phenotype which is de- pendent on the exp genes and that sfx mutations s u p press this mucoid phenotype.
MATERIALS AND METHODS
Bacterial strains, plasmids, transposons, nomenclature, me- dia and growth conditions: The major strains, plasmids and transposons employed in this work are listed in Table 1. All R. meliloti are derivatives of strain SU47. Many of the strain constructions are described in RESULTS, and others are outlined below.
Strain Rm8002 was originally identified as an alkaline phos- phatase deficient mutant and was isolated following ethyl methanesulfonate mutagenesis of Rm1021 (LONG et al. 1988). The mutant was subsequently found to be deficient in utili- zation of several different organic phosphate sources and is thus assumed to carry a mutation in a regulatory gene such as the Escherichia coli p h o B gene (cited in ZHAN et al. 1991).
Strain RmG256 was made by transducing RmF378 contain- ing a5117::Tn5 (but not sfx-I) into RmG167 which contains R5122::Tn5-132 and screening the resultant colonies for both markers. Together these insertions flank the sfx-1 locus (see Figure I ) .
R. meliloti strains RmF378 and RmF379 were constructed by transducing Nm' from strains RmF338 and RmF339 respec-
tively into Rm1021. To screen for the presence of the sfx-1
mutation, A5408::Tn5-233 and AF114::Tn5-233 were trans- duced from Rm5408 and RmFl14 respectively, into both RmF378 and RmF379 by selecting for Gm'Sp' transfer. Upon inoculation of alfalfa plants, RmF378 transductants formed Fix- nodules whereas RmF379 transductants formed Fix+ nod- ules. Thus unlike RmF378, strain RmF379 carried the sfx-1
mutation.
R. meliloti strain RmC479, carrying R5025::Tn5, sfx-2 was constructed by replacing the AF114::Tn5-233 deletion in RmF346 with the corresponding wild-type region. That was accomplished by transducing R5025::Tn5 (Nm'), which lies internal to AF114, from Fh5348 into RmF346. Strain RmG497 carrying fl5033::Tn5-233 and sfx-2 was constructed by trans- ducing R5033::Tn5-233 (Gm'Sp') which is linked to a5025 from Rm5356 into RmG479 and screening for Nm" transduc- tants. (Rm1021 strains carrying R5025::Tn5 or R5033::Tn5- 233 have a wild-type symbiotic phenotype.)
R. meliloti strain RmG591 which contains the sfx-1 allele in an otherwise wild-type background was derived from RmF379. First the Tn5 insertion a5118 was replaced with Tn5-233 to give strain RmF994 (05118::Tn5 sfx-1). Nm' was then trans- duced from Rm5439 [pck-I::TnV (Nm')] into RmF994 and a Nm' GmsSps transductant was purified. As the distance from pck to the sfx-I mutation should be approximately 160 kb ( i . e . , the packaging capacity of the transducing phage) it was very unlikely that a Nm' Gm'SSp" transductant would lose the sfx-1
allele. One such colony was purified and transduced with a phage lysate grown on the wild-type Rm1021. Transductants which grew on succinate minimal medium were then selected. One such transductant was purified and designated RmC591. To simplify descriptions, we refer to the large deletions in Rm5408 and RmF114 as A5408 and AF114 respectively. Simi- larly, the 12-kb deletion which removes the ndvF locus is re- ferred to as AG439.
Complex medium was LB; minimal medium modified M9; antibiotic concentrations for R. meliloti and E. coli; growth conditions; all were as previously described (CHARLES and Fr-
NAN 1991; FINAN et al. 1984). Low osmolarity glutamate-yeast
extract-mannitol-salts (GYM) medium was prepared as de- scribed by DYLAN et al. (1990a) except that l mM sodium phos- phate was substituted for 1 mM potassium phosphate. GYM thus consisted of 1 mM glutamate (monosodium salt), 0.02%
(w/v) yeast extract (Difco), 2.5 mM mannitol, 1 mM NaHPO, (pH 6.8), 0.5 mM CaCl,, 0.5 mM MgSO,. For solid medium, GYM was used at 0.5 X concentration.
Genetic manipulations and DNA methodology: Bacterial matings, 4M12 transductions and transposon replacements, DNA isolation, restriction enzyme digestion and Southern hy- bridization were carried out as previously described (CHARLES and FINAN 1991; FINAN et al. 1984). Phage 4M12 lysates used to isolate linked transposon insertions were grown on previ- ously prepared random Tn5, Tn5-132 and Tn5-233 insertion banks (CHARLES and FINAN 1990). O n average, 0.7% of the in- sertions in these banks are linked in transduction to any site on the R. meliloti genome (FINAN et al. 1984).
R . meliloti ndvF Suppressor Alleles
TABLE I
1235
Bacterial strains, plasmids, phage and t r a n s p o s ~ ~
Strain, plasmid
o r transposon Relevant characteristics Source, reference or construction
Rhizobium meliloti Rm1021 Rm8002 Rm5000 Rm5348 Rm5356 Rm5439 Rm5408 Rm7013 Rm7031 Rm7055 RmFll4 RmF263 RmF338 RmF339 RmF346 RmF378 RmF379 RmF427 RmF993 RmF994 RmG167 RmG203 RmG204 RmG256 RmG425 RmG439 RmG490 RmG491 RmG479 RmG497 RmG5 14 RmG588 RmG591 RmG631 RmG635 RmG637 Escherchia coli MT607 MT616 DH5a RM1021 derivatives
Plasmids and phage pRK2013 pRK600 pRK607 pRK7813 pTH36 pTH56 pTH57 pGMI102 (DM12 Transposons
T n 5 Tn5-11 Tn5-132 Tn5-233 Tn5-mob Tn V SU47 str-21 SU47 str-21 Pho- SU47 rif-5
Rm1021 05025::Tn5 Rm1021 R5033::Tn5-233
pck-1 ::TnV
A5408 (R5033-5007::Tn5-233) exoBI3::Tn5
exoA3I::Tn5 exoF55::Tn5
AF114 (05033-5064::Tn5-233) A05033-5007::Tn5-233, sfx-I
A05033-5007::Tn5-233, R5117::Tn5, sfx-I
A05033-5007::Tn5-233, R5118::Tn5, sfx-1
AR5033-5064::Tn5-233, sfx-2 05117::Tn5
05118::Tn5, sfx-I
AR5033-5007::Tn5-233, R5122::Tn5-132 05117::Tn5-233
05118::Tn5-233, sfx-1
R5122::Tn5-132
A05033-5007::Tn5-233, sfx-4
AR5033-5064:Tn5-233, sfx-5 R5117::Tn5 05122::Tn5-132 A05033-5064::Tn5-233, sfx-3 AG439 (AndvF HindII1::Nm (13 kh)) ndvF-I. 7R Sp'
ndvF-5.?R Sp' 05025::Tn5, sfx-2 05033::Tn5-233, sfx-2
AG439 ndvF, sfx-2, R5025::Tn5 nduB
sfx-I, AG439( ndvF)
R5118::Tn5, ndvB::Tn5-233, sfx-I
R5118::Tn5, ndvB::Tn5-233, sfx-2
MM294A recA56 MT607 (pRK600)
endAl hsdRl7 supE44 thi-1 recAl gyrA96 relAl sfx-l
A(argF-lacZYA)U169 480dlacZAM15
ColEl replicon with RK2 transfer region, Nm-Km' pRK2013 npt::TnP, Cm', Nm-Km'
pRK2013::Tn5-233; Nm-Km', Gm', Sp'
RK2 derivative carrying pUC9 polylinker and cos site, Tc' 7.3 BamHI fragment containing nduF in pRK7813 pRK7813 clone carrying sfx-1
pRK7813 clone carrying sfx-1
Nms derivative of RF'4
R. meliloti bacteriophage
Nm', Sm'
oriT of pRK2 cloned into Tn5-233, Gm'-Sp' 0 t'
Gm', Sp'
T n 5 containing mob site from RK2 T n 5 containing pSClOl oriV, Nm'
MEADE et al. (1982) LONG et al. (1988) FINAN et a!!. (1984)
CHARLES and FINAN (1990) CHARLES and FINAN (1990) FINAN et al. (1988) CHARLES and FINAN (1991) FINAN et al. (1985)
LEIGH et al. (1985) LEIGH et al. (1985) CHARLES and FINAN (1991) Fix+ nodule isolate from Rm5408 This work
This work
Fix' nodule isolate from RmFl14 Q(RmF338)
-
Rm1021, Nm' Gms-Sps Q(RmF339) + Rm1021, Nm' Gms-SpsQ(Tn5-132 Bank)
-
RmF339, Gm'-Sp' Ot' Nm' Fix- Tn5-233 replacement of 05117::Tn5 in RmF378 Tn5-233 replacement of 05118 in RmF379 Q(RmF427)-
Rm1021, Ot'Fix' nodule isolate from Rm5408 Fix' nodule isolate from RmFll4 Q(RmF378) + RmG167, Nm', Ot' Fix' nodule isolate from RmFl14 CHARLES et al. (1991)
CHARLES et al. (1991) CHARLES et al. (1991) Q(Rm5348) + RmF346, Nm'
(D(Rm5356) + RmG479, Gm'-Sp', Nms
This work J. GLAZEBROOK
Q(Rm1021)
-
RmG560, succinate', Nm', Gm'-Sps Q(RmG424) + RmF346, Nm'(P(RmG558) + RmF379, Nm', Gm'-Sp' (P(RmG558) + RmG479, Nm', Gm'-Sp'
FINAN et al. (1986) FINAN et al. (1985)
B.R.L. Inc.
FIGURSKI and HELINSKI (1 979) FINAN et al. (1986)
DEVOS et al. (1986)
JONES and GUTTEMON (1987)
CHARLES et al. (1991) This work
This work
MEADE et al. (1982) FINAN et al. (1984) BERG and BERG (1987) FINAN et al. (1986) BERG and BERG (1987) DE Vos et al. (1986) R. SIMON (1984) FURUICHI et al. (1985)
Abbreviations are as follows: Ap, ampicillin; Cm, Chloramphenicol; Gm, gentamycin; Km, kanamycin; Nm, neomycin; Ot, oxytetracycline; Sm, streptomycin; Sp, spectinomycin; Tc, tetracycline; QM12 transducing lysates are indicated by Q preceeding the strain number. Details regarding the Pho- mutation in Rm8002 are described in the text under Bacterial Strains. For strain constructions, an arrow indicates transduction from the indicated QM12 lysate into the recipient strain. Example: Q(RmF338)
-
Rm1021, Nm', means a QM12 lysate grown on RmF338 was used to transduce Nm' into Rm1021.1236 I. J. Oresnik, T. C. Charles and T. M. Finan
C ' I A I A A A I A
6 %
-* 20%
8% b
4%
4
32% 58%
48% b 4 20%
COTRANSDUCTION FREQUENCY ( % ) 4 48%
7 0 % 21%
L A 23% ___* 64%
r -
'
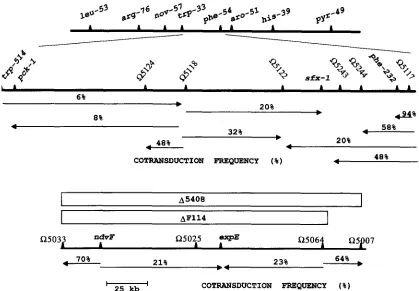
2 5 J c b ' COTRANSDUCTION FREQUENCY (%)FIGURE 1.-Genetic map showing the location of s f x - l on the R. meliloti chromosome. The top diagram shows the chromosomal map of R . meliloti and is based on the map of MEADE and SIGNER 1977. The middle diagram shows the location of sfx-1 together with the linked transposon insertions. The isolation of insertions and the derivation of the map is described in the text and Figure 2. The bottom diagram shows the ndvF gene region together with the linked insertions which were used in this work. The map is taken from CHARLES et al. (1991).
solate bacteria from root-nodules, the nodules were first sur- face sterilized by treating the nodule with 1% sodium hypochlorite for 1 min. Following a thorough washing in ster- ile distilled water the nodules were then crushed in a drop of LB medium containing 0.3 M sucrose. The nodule contents were then plated onto LB medium and following incubation for 3-4 days, desired colony isolates were single colony purified three times prior to subsequent use. Acetylene reduction assays were done in triplicate on the root systems of 9 or 10 plants as previously described (CHARLES et al. 1991).
RESULTS
Suppressors of ndvR R. meliloti strains Rm5408 and RmFll4 carry the deletion mutations A5408 and AF114, respectively, which remove the ndvFregion of megaplas- mid pRmeSU47b. Loss of the ndvF locus from these strains was previously shown to result in the induction of small white nodules on alfalfa which failed to fix N,
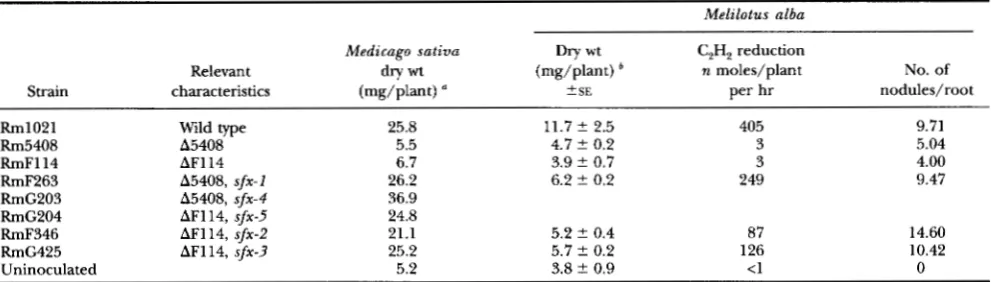
(Fix-). We previously reported that pink Fix+ nodules occasionally arose on plants inoculated with these strains (CHARLES et al. 1991). In the present work we isolated bacteria from five such pink nodules (see MA- TERIALS AND METHODS) obtained from five independent nodulation experiments. These isolates were designated RmF263, RmF346, RmG203, RmG204 and RmG425. Upon reinoculation of alfalfa, all formed Fix' nodules as determined by dry weight analysis of the inoculated plants (Table
2).
We also examined representativestrains for their symbiotic phenotype on sweet clover ( M . alba cv. Polara) which is another symbiotic host of
R. meliloti (Table
2).
Quantitation of the cumulativeN,-fixation phenotype as determined by measurement of the plant dry weights, clearly showed that the Fix phe- notype was not completely restored on M . alba in the three isolates examined. Measurements of nitrogenase activity, via the reduction of acetylene to ethylene, re- vealed that plants inoculated with isolates RmF263, RmF346 and RmG425 had 61%, 21%, and 31%, respec- tively, of wild-type activity. Quantitation of the number of nodules formed on the root systems of inoculated plants showed that whereas the nodule number of the
two ndvF deletion mutants was reduced compared to Rm1021, the nodule number was restored on roots in- oculated with RmF263 and RmG425. The slightly greater number of nodules on the roots of plants in- oculated with RmF346 may reflect the low amount of N,
fixed by this strain.
R. meliloti ndvF Suppressor Alleles 1237 TABLE 2
Symbiotic phenotype of Fiu'second site revertants of ndvF deletion mutants
Melalotus alba
Medicago sativa Dry wt C,H, reduction
Relevant dry wt (mg/plant) n moles/plant No. of
Strain characteristics (mg/plant) 2 SE per hr nodules/root
Rm1021 Rm5408 RmFl14 RmF263 RmC203 RmG204 RmF346 RmG425 Uninoculated
Wild type A5408 AF114 115408, sfx-I 155408, sfx-4
AF114, sfx-2 AF114, sfx-3 AF114, sfx-5
25.8 11.7 ir 2.5 405
5.5 4.7 ir 0.2 3
6.7 3.9 ir 0.7 3
26.2 6.2 ir 0.2 249
36.9 24.8
21.1 5.2 ? 0.4 87
25.2 5.7 2 0.2 126
5.2 3.8 ir 0.9 <1
9.71 5.04 4.00 9.47
14.60 10.42
0
Shoot dry weights were determined on 23-day-old alfalfa ( M . s a t i v a ) plants. Values represent the mean of 17-19 plants.
*
Sweet clover ( M . a l b a ) plants were hamested 28 days after inoculation. Dry weight and nodule numbers were determined from analysis of at least 15 plants per treatment.the ndvF locus (CHARLES and FINAN 1991). Moreover, when Gm'Sp' marking the A5408 deletion was trans- duced from RmF263 into Rm1021, all 20 transductants examined were Fix- when inoculated on alfalfa plants. The above data suggested that RmF263, RmF346, RmG203, RmG204 and RmG425 carried second site mu- tations which suppressed the Fix- phenotype of the ndvF deletion mutations. The suppressor mutations in strains RmF263, RmF346, RmG425, RmG203 and RmG204 were designated sfx-1 through sfx-5, respectively.
Identification of sfx-1: To manipulate the sfx muta- tions we first isolated Tn5 insertions linked to the sfx-1 mutation in RmF263. That was accomplished by employ- ing a two-step transduction strategy. First, several thou- sand random Tn5 insertions were transduced from the wild-type Rm1021 background into RmF263 by selecting for Nm' (FINAN et al. 1984). Among these transductants a low percentage ( c a . 0.7%) should have Tn5 linked in transduction to the suppressor locus. A second lysate was prepared from the pooled Nm' RmF263 transductants. Nm' was transduced from this pool into the Fix- strain Rm5408. Transductants which acquired the suppressor locus (and would therefore be Fix+) via a linked Tn5 insertion were isolated from pink Fix' nodules obtained three weeks following inoculation with approximately 2,000 pooled Rm5408 Nm' transductants. The Tn5 in- sertions in two independent nodule isolates, strains RmF338 and RmF339, were designated 05117 and 0 5 1 18 respectively. To determine if these two insertions were linked to each other, both Tn5 insertions were re- placed with Tn5-233 (Gm'Sp'). In subsequent experi- ments, 6% of colonies obtained after transduction of Nm' from strain RmF378 carrying 05117::Tn5 into RmF994 carrying a5118::Tn5-233 resulted in loss of Gm'Sp'. Similarly 2% of the colonies obtained after transduction of Nm' from strain RmF379 carrying a 5 1 18::Tn5 into RmF993 carrying 05117::Tn5-233 re- sulted in loss of Gm'Sp'. Thus a5117 and 05118 were
ca. 4% linked in transduction.
Tn5-132 insertions linked to the sfx-1 region were ob-
tained by transducing a bank of random Tn5-132 inser- tions from Rm1021 into strains carrying 05117::Tn5 and 05118::Tn5 and screening for Nms (see MATERIALS AND METHODS). Similarly Tn5-233 insertions 05243 and 05244 were isolated on the basis of their linkage to 05122::Tn5-132. The positions of 05122, 05243 and 05244 relative to insertions 05117 and 05118 were de- termined following analysis of data from three factor crosses. The deduced order of these insertions is shown in Figure 1.
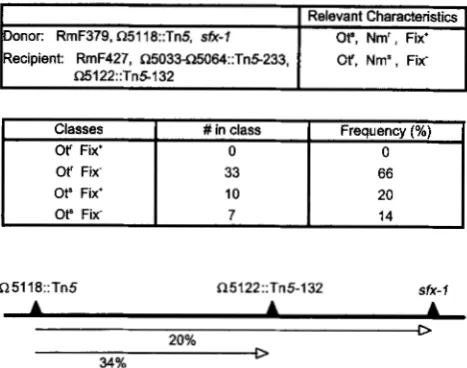
To map the sfx-1 allele in relation to 05118::Tn5 and 05122::Tn5-132 we transduced Nm' from RmF379 carrying 05118::Tn5 sfx-1 into RmF427 carrying R5122::Tn5-132 and A5408 (see Materials and Methods for construction of strains). Fifty Nm' transductants were purified and screened for loss of OtR and cotransduction ofFix'. The results are consistentwith the order 0 5 1 18- 05122-sfx-1 with sfx-1 20% linked to 05118 (Figure 2).
1238 I. J. Oresnik, T. C. Charles and T. M. Finan
I
Relevant Characteristics onor: RmF379, Q5118::Tn5, sfx-II
Ot", Nm', Fix+ Recipient: RmF427, Q503345064::Tn5233, Or, Nrn', Fix'Q5122::Tn5132
I
ClassesI
# in class1
Frequency (%)Of Fix' 0
I
0Ot' Fix- Ot" Fix' Ot" Fix-
33
10 7
66
20
14
Q5118::Tn5 Q5122::Tn5-132 sfk- I
20% 0
34% D
FIGURE 2.-Three-factor cross to determine the relative po- sition of sfx-l. Fifty neomycin resistant (Nm') transductants were purified and screened for loss of oxytetracycline sensi- tivity (Ot') and symbiotic N,-fixation (Fix) phenotype. The lat- ter was scored 3 weeks following inoculation of alfalfa seed- lings. Fix' strains formed large pink nodules on green vigorously growing plants. Fix- strains formed small white nod- ules on small chlorotic plants. The deduced order of 05188, 05122, and sfx-1 is shown.
was weakly linked to pck-1::TnV (17/200 colonies screened had lost TnV::pck-I ).
The p h e - 5 4 marker was shown to be close and clock- wise from trp-33 on the chromosomal map of Rm1021
(MEADE and SIGNER 1977). Results from a three factor cross employing a recipient carrying phe-232:Tn5- 233 and a donor strain carrying 051 17:Tn5 and 05122::Tn5-132 showed that p h e - 2 3 2 was tightly linked to 05117 and lay between 05117 and 05122 (Figure 1).
sfx alleles map to two different loci: To determine whether other nduF suppressor mutations mapped to the sfx-1 locus we transduced the wild-type sfx-1 locus flanked by the insertions 05117::Tn5 and 05122::Tn5- 132 into the Fix' pseudorevertant strains RmF263, RmF346, RmG203 and RmG204. The symbiotic pheno- type of the resulting Nm' Ot' transductants (Figure 3)
clearly indicated that sfx-1, sfx-4 and sfx-5 map to the same locus flanked by the inserts 05122 and 05117. In addition the data also suggested that the suppressor mu- tation, sfx-2, in RmF346 mapped to a separate locus not tightly linked to the Kl5122-lR5117 region.
As we had previously obtained evidence that the nduF locus encodes membrane-associated proteins (CHARLES
et al. 1991), we examined strains carrying nduFsuppres-
sor mutations for cell surface alterations as indicated by altered sensitivity to bacteriophage or detergents. All of the strains showed the same sensitivity to the eight bac- teriophage examined. However, unlike Rm1021, Rm5408, RmFl14, RmF263, RmG203 and RmG204, strains RmF346 and RmG425 were found to be sensitive to the detergents deoxycholate, sodium dodecyl sulfate and Sarkosyl and to the antibiotic bacitracin. The al-
tered chemical sensitivities of strains RmF346 and RmG425 suggested that the sfx-2 and sfx-3 mutations resulted in an alteration in the permeability of the cell envelope. We have isolated a Tn5 insertion, 05258, which was 70% linked in transduction to the deoxy- cholate sensitivity mutation in RmF346. Transduction of 05258::Tn5 into RmG425 also revealed a 70% linkage to the deoxycholate sensitivity mutation in that strain
(ORESNIK 1990). Thus on the basis of the detergent phe- notype, and genetic linkage, the suppressor mutations were divided into two classes; class I consisting of sfx-I, sfx-4 and sfx-5 alleles in strains RmF263, RmG203 and RmG204, respectively, and class I1 consisting of the
sfx-2 and sfx-3 alleles in strains RmF346 and RmG425, respectively.
Cloning of the sfx-1 locus: Two independent pRK7813 cosmid libraries were constructed with partial BamHI
digested DNA from RmF263 which carries the sfx-1 al- lele and is deleted of the nduF locus. Each bank con- sisted of approximately 2300 clones. The cosmid banks were mated into the Fix- nduF deletion strains RmFll4 and Rm5408 and between 14,000 and 31,000 Sm'Tc' transconjugants were pooled and inoculated onto alfalfa seedlings planted in Leonard pot assemblies.
After
20
days of growth, nodules from green Fix' plants were surface sterilized either individually or in a group of 10 and spread on LB-tetracycline (10 pg/ml) agar. Resultant colonies were streak purified once and plasmids were mated into E . coli MT607. Restriction analysis of 36 isolates yielded three distinct cosmid types. Representatives of two classes were designated pTH56 and pTH57. The third cosmid class did not contain any insert and their origin is unclear. Analysis of plasmids pTH56 and pTH57 revealed that while these plasmids differed from each other, each appeared to share an 18-kb BamHI and a 2-kb Hind111 fragment. When the 18-kb BamHI fragment from pTH56 was gel purified and labeled it hybridized to an l&kb fragment on a Southern blot of BamHI-restricted pTH57 DNA. This result confirmed that these two cosmids carried a com- mon BamHI fragment.The ability of pTH56 and pTH57 to suppress the Fix- phenotype of the nduF mutants was quantitated following the transfer of these plasmids from E. coli into Rm5408 and RmFll4. Three independent transconjugant cultures of RmFl14 or Rm5408 containing either pTH56 or pTH57 were inoculated onto alfalfa seedlings. After 23 days the nitrogenase activity of root-nodules was determined by the acetylene reduction technique. In addition, the dry weights of the shoots of these plants were measured to assay
cumulative nitrogen fixation (Table 3). The results indi- cate that the cosmids restore a Fix' phenotype to the Fix-
deletion strains. The variation in the restoration of N2-
R . meliloti nduF Suppressor Alleles 1239
R5122::Tn5-132 (Ot') Q5117::Tn5 (Nm')
A
A
Donor: Fix+Recipient: A ndvf
SfX? Fix+ pseudorevertant
Strain
+
wild type Rm1021
Fix Ot' Nm' Genotype
+
+
+
1
-
1
-
1
-k m F l 1 4 lAFl14
1
-
1
-
1
-
1
I I
I I
.
mF263 (A5408, SI%-7
I
-
I
-
I
+
FIGURE 3.-Mapping of the nduF suppressor alleles, sfx-1, sfx-2, sfx-4 and sfx-5 by trans- duction. The Fix' donor strain RmG256, car- ried R5122::Tn5-132 (Ot') and R5177::Tn5
(Nm') flanking the wild-type sfx-l locus Le., did not suppress nduF. Ten Nm', Ot' transduc-
tants of Rm1021, RmF263, RmG203, RmG204 and RmF346 were selected and screened for symbiotic N,-fixation three weeks following in- oculation of alfalfa seedlings as described for Figure 2.
Recombinants
I
+
+
RmG203 1~5408, sfX-4
I
-
I
-
I
+
I I I
+
+
AF114, sfX-5
I
-
I
-
I
+
Recombinants
+
+
+
Recombinants
+
AFI 14, SfX-2
RmF346
+
+
TABLE 3
Symbiotic phenotype of ndvF deletion mutants carrying the cosmid clones pTH56 and pTH57
ARA" Percent Percent nM/plant/hr wild-type Dry wt wild-type
Strain t S E ARA mg/plantc dry wt
Rm1021 818.7 t 22.2 100 15.4 100
RmFll4 29.3 ? 3.3 4 5.9 38 RM5408 97.0 ? 7.7 12 6.4 42 Rm5408
(pTH56) 994.7 ? 6.1 121 14.4 94 Rm5408
(pTH57) 595.8 ? 68.6 73 11.1 72 RmFl14
(pTH56) 622.4 t 43.5 76 10.5 68
RmFl14
(pTH57) 329.7 5 51.7 40 10.2 66 RmF263 952.0 t 66.7 116 17.7 115
Uninoculated 16.0 ? 0.0 2 5.7 37
a Acetylene reduction activity.
In all cases the root systems of at least nine alfalfa plants were
For dry weight analysis the shoots of at least 26 alfalfa plants were assayed. Values are the mean t standard error.
weighed.
Direct evidence that the cloned insert DNA in pTH56 was from the mapped sfx-I region was obtained by Southern blot analysis of total DNA isolated from strains carrying transposon insertions which mapped to the
sfx-1 region. Thus blots of BamHI-restricted DNA probed with pTH56 revealed that Tn5-233 insertions
R5243 and a5244 (Figure 1) lay within 18- and 7-kb BamHI fragments, respectively, both of which were present in pTH56. Conversely, hybridization of labeled pTH56 to a Southern blot of BamHI-restricted total DNA from a strain carrying R5122::Tn5-132 (Figure 1)
revealed that this insertion lay outside of the region car- ried in plasmid pTH56.
nduF mutants exhibit a low osmolarity mucoid
phe-
notype: Mutants of R. meliloti strain 102F34 defective in the production of periplasmic
El
,2-glucan are impaired in the ability to grow on a low osmolarity GYM medium1240 I. J. Oresnik, T. C. Charles and T. M. Finan
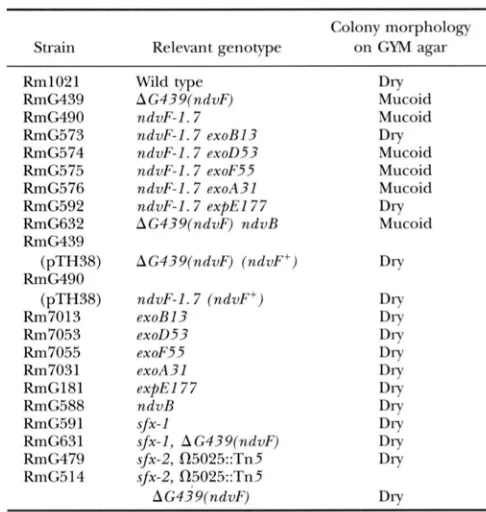
while Fix+ insertions adjacent to ndvF were non- mucoid (see CHARLES et al. 1991; Figure 4). Interest- ingly we found that in a Rm8002 background, nine ndvE:TnphoA insertion mutant strains had a dry wild- type phenotype on GYM. When transduced into a wild-type Rm1021 background all of these insertion mutations resulted in a mucoid colony phenotype on GYM medium (see DISCUSSION). Moreover, the mucoid
phenotype was reversed to the dry wild-type pheno- type following introduction of the wild-type ndvFlocus on plasmid pTH38 (Table 4). Thus there was a cor- relation between the Fix- and low osmolarity mucoid phenotypes.
To investigate whether /3-1,2-glucan or either of the two known exopolysaccharides of R. meliloti were in- volved in the mucoid colony phenotype, double mutants were constructed by transduction of Nm' from various Tn5 marked polysaccharide mutations into the Sp', ndvF interposon mutant strains RmG490 (=Rm1021 ndvF-I. 7::RSp') and RmG491 (=Rm1021 ndvF- 5.3:RSp'). Testing of these double mutants (Table 4) revealed that low osmolarity mucoidy was dependent on the polysaccharide synthetic genes exoB and expE, but did not require exoA, exoF, exoD or ndvR. In
R.
meliloti, exoA, exoF, and exoD are required for succinoglycan pro- duction (LEIGH and LEE 1988), expE is required for ex- opolysaccharide I1 production, ndvB is required for pl,2-glucan production ( GLAZERROOK and WALKER 1989), and exoR is required for all of these (GLAZERROOK and WALKER 1989;ZHAN
et al. 1989; LEIGH and LEE 1988). These results therefore suggested that neither exopo- lysaccharideI
(succinoglycan) nor p112-glucan synthe- ses are involved in generating the mucoid phenotype, but that exopolysaccharideI1
synthesis is required. We note that the Fix- mutants Rm5408 and RmF114 carry1
FIGURE 4,"Colony morphology of
R. meliloti wild-type strain 1021 and
the ndvF mutant RmG439 GYM me-
dium with and without 100 mM NaCI.
TABLE 4
Genetic requirement for low osmolarity-linked mucoidy of ndvF mutants
Colony morphology
Strain Relevant genotype on GYM agar
Rm1021 RmG439
RmG490 RmG573 RmG574 RmG575 RmG576 RmC592 RmG632 RmG439
RmG490
Rm7013 Rm7053 Rm7055 Rm7031 RmG181 KmG588 KmC591 RmG63I RmG479 RmG514
(pTH38)
(pTH38)
Wild type
AG439(ndvF) ndvF-I. 7 ndvF-I. 7 exoB13 ndvF-I. 7 exon53
ndvF-I. 7 exoF55
ndvF-1.7 exoA31 ndvF-I. 7 expEl77 AG439(ndvF) ndvR
AG439(ndvF) (ndvF+)
ndvF-I. 7 (ndvF+) exoR I3
exon53
exoF55
exoA31
ndvR
sfx-I, AG439(ndvF)
sfx-2, R5025::Tn5
sfx-2, R5025::Tn5
AG439/ndvF) e x f E I 77
sfx-1
Dry
Mucoid Mucoid
Mucoid Mucoid Mucoid
Mucoid
Dry
Dry
Dry
Dry Dry
Dry Dry Dry Dry Dry Dry Dry Dry
DIT
R . meliloti nduF Suppressor Alleles 1241
Suppression of the ndvF-mucoid phenotype by sfx-1
and sfx-2 To explore the relationship between the ndvF dependent low osmolarity phenotype and the Fix- phenotype of ndvF mutants we examined whether the
sfx mutations which were isolated via their ability to sup- press the Fix- phenotype of nduF mutations could also suppress the mucoid phenotype of the ndvF mutations.
As
all five suppressor strains were derived from the ndvFexp mutants Rm5408 or RmFll4, and the mucoid phe- notype is dependent on ex$ (above), it was necessary to construct strains which were ndvF+ and exp+ but carried
sfx mutations. We used the sfx-1 and sfx-2 alleles to rep- resent the two classes of suppressor mutations. The nduF deletion AG439 and other ndvF insertion mutations were transduced into the sfx-1 and sfx-2 backgrounds and all of the resulting transductants were found to be dry on GYM agar ( e . g . , RmG631 and RmG514; Table 4). Thus sfx-1 and sfx-2 reversed both the free-living and symbiotic phenotypes associated with ndvF mutations.
To determine if the sfx-1 locus cloned in plasmid pTH56 would suppress the ndvFlow osmolarity mucoid phenotype, pTH56 was mated into the 12-kb ndvF de- letion strain RmG439, and into strains carrying single ndvFxTn4 insertion mutants. All of the transconjugants formed dry colonies on GYM agar showing that pTH56 suppressed the mucoid phenotype of the ndvF mutants examined.
sfx-1 and sfx-2 mutations fail to suppress the Fix- phe- notype of Exo-
and
NdvB- mutants ofR.
meliloti: As second-site mutations which suppress the Fix- pheno- type of exo and nduB mutants of R. meliloti have been described, it was of interest to determine if either thesfx-1 or sfx-2 mutations described in this report could also suppress the Fix- phenotype of exo and ndvB mu- tants. To eliminate any possible influence resulting from an ndvF mutation, we first constructed strains which carried the sfx-1 (RmF379 and RmG591) and sfx-2
(RmG47 and RmG497) alleles in a wild-type n d v p back- ground (for strain constructions, see Materials and Methods). Various exo and nduB mutations were then transduced into the sfx-1 and sfx-2 backgrounds and the symbiotic phenotype of the resulting double mutants were determined (Table 5). The dry weights of plants inoculated with the sfx exo and sfx nduB double mutants were similar to the dry weights of uninoculated plants and plants inoculated with the exo or ndvB mutants. These data clearly show that no suppression of the Fix- phenotype of the exo or ndvB mutations occurred.
DISCUSSION
Our results show that the Fix- phenotype of R. meliloti ndvF mutants on alfalfa can be suppressed by mutations of least two distinct loci represented by sfx-1 and sfx-2.
Neither of these alleles suppressed the Fix- phenotype of ndvB or exo mutations, indicating that sfx-1 and sfx-2
are distinct from previously reported ndvB or ex0 s u p
TABLE 5
Symbiotic phenotype of sfx ex0 and sfx nduF double mutants of R. rneliloti
Plant dry wt
Strain Relevant genotype (mg)
Experiment 1 Rm1021 RmG59 1
Wild type 53.7
RmG720 RmG721
sfx-I exoF:Tn5 4.8
RmG717
sfx-I exoA::Tn5 6.7
RmG631
sfx-l exoB::Tn5 6.2
sfx-1 A G 4 3 9 (ndvF) 68 RmG514 sfx-2 A G 4 3 9 (ndvF)Cl5025::Tn5 60 RmG497 sfx-2 Cl5033::Tn5-233 57 RmG725 sfx-2 exoF:TnS Cl5033::Tn5-233 5.7 RmG726 sfx-2 exoA::Tn5 R5033::Tn5-233
RmG722
5.6 sfx-2 exoB::Tn5 05033::Tn5-233
Rm7031
5.5 exoA3I ::Tn5
Rm7013
7.2
exoB::Tn5 6.2
Uninoculated 5.9
Experiment I1 Rm1021 RmF379
Wild type 13.1
sfx-1 Cl5118::Tn5 RmG479
13.1 sfx-2 Cl5025::Tn5
RmG588
13.9
ndvB::Tn5-233 5.3
RmG635 sfx-1 ndvB::Tn5-233 4.5
RmG637 sfx-2 ndvB::Tn5-233 5.9
Uninoculated 4.4
In experiment I, alfalfa plants were harvested 34 days post inocu- lation; in experiment 11, alfalfa plants were harvested 24 days post inoculation. Values are the means from triplicate samples.
sfx-l 63
pressor loci in R. meliloti ( D m et al. 1990b; GLAZE- BROOK and WALKER 1989; PUTNOKY et al. 1990; WILLIAMS et al. 1990; ZHAN et al. 1989). Moreover the chromo- somal map position of sfx-1 between trp-33 and p h e is clearly different from the megaplasmid located alleles which suppress ndvA and ndvB or exo mutants (GLAZE-
BROOK and WALKER 1989; NAGPAL et al. 1992; PUTNOKY et
al. 1990; WILLIAMS et al. 1990). The sfx alleles showed considerable variability in their ability to suppress the ndvFdependent Fix- phenotype with M . alba as the host plant (Table
2).
This could indicate a direct interaction(recognition) of the sfx determinants with the host plant. Alternatively, the result may reflect slight differ- ences in the physiological status of the M . sativa us. the M . alba root systems.
1242 I. J. Oresnik, T. C. Charles and T. M. Finan
sistent with our earlier results suggesting the the mu- coidy results from increased production of EPS 11. This result also suggests that the nduF mucoid phenotype is mediated through the phosphate regulatory system. It is difficult to reconcile the latter conclusion with the find- ing that the nduF dependent mucoid phenotype is re- versed upon addition of 100 mM NaCl to the GYM me- dium. We note however that there are several reports of interactions between phosphate and osmotic regulatory effects in other microorganisms (MEWR et al. 1990;
SHORTRIDGE et al. 1992; VILLAREJO et al. 1983).
The Fix- phenotype is not dependent on the nduF induced mucoidy as the deletion mutants Rm5408 and RmFll4 are Fix-, both lack the n d v F and e x p gene clus- ters and form dry colonies on GYM medium. The Fix- phenotype of these mutants is restored upon transfer of the nduF gene region (CHARLES et al. 1991). Moreover suppression of the nduF Fix- phenotype is not depend- ent on the presence of exp genes as all five suppressor alleles were originally isolated in the Rm5408 and RmF114 backgrounds. However our data do show that there is an association between the mucoid phenotype and the Fix- phenotype of nduF mutants. First, eighteen out of nineteen nduF insertion mutants generate a mu- coid phenotype and second, the sfx-1 and sfx-2 alleles suppress both the Fix- and mucoid phenotype of nduF mutants.
It would seem likely that the mucoid colony morphol- ogy is a pleiotropic phenotype resulting from the nduF mutation. Thus even when the biochemical function of the nduF locus is known, the question of whether the Fix- symbiotic phenotype is a direct or indirect conse- quence of nduFmutations is likely to remain unresolved. Our ability to manipulate the sfx loci as described in this report should greatly facilitate the resolution of this issue.
It is perhaps worth noting that the mucoid phenotype generated by nduF insertion mutants was observed after much of the genetic analysis of the sfx-1 locus reported in this paper was carried out. Thus cosmids pTH56 and pTH57 were isolated by their ability to restore a Fix' phenotype to nduF mutants. Upon subsequent testing, both clones were found to suppress the nduF mucoid colony phenotype. We are confident that these cosmids carry the sfx-1 locus, as both share common BamHI and Hind111 restriction enzyme fragments and the cloned DNA is contiguous with the genetically identified sfx-1
locus. Employing the suppression of the mucoid phe- notype as a screen we are now localizing the sfx-1 locus on pTH56 and are carrying out a detailed molecular analysis of this region.
We thank BRIAN DRISCOLL., MAGNE OSTERAS and SKWE BARDIN for comments on this manuscript. This work was supported by operating grants fron the Natural Sciences and Engineering Council of Canada to T. M. F.
LITERATURE CITED
BERG, C. M., and D. E. BERG, 1987 Uses of transposable elements and maps of known insertions, pp. 1071-1107 in Escherichia coli and Salmonella typhimurium Cellular and Molecular Biology, edited by F. C. NIEDHARDT, J. L. INGRAHAM, K. B. Low, B. MAGASANIK et al.
American Society for Microbiology, Washington, D.C.
CHARLES, T. C., and T. M. FINAN, 1990 Genetic map of Rhizobium meliloti pRmeSU47b. J. Bacteriol. 172: 2469-2476.
CHARLES, T. C., and T. M. FINAN, 1991 Analysis of a 1600 kilobase
Rhizobium meliloti megaplasmid using defined deletions gener- ated in uivo. Genetics. 127: 5-20.
CHARLES, T. C., NEWCOMB, W. and T. M. FINAN, 1991 nduF, a novel locus located on megaplasmid pRmeSU47b (pEXO) of Rhizo- bium meliloti, is required for normal nodule development. J. Bac- teriol. 173: 3981-3992.
DE VOS, G. F., G. WALKER and E. R. SIGNER, 1986 Genetic manipula- tions in Rhizobium meliloti utilizing two new transposon T n 5 de- rivatives. Mol. Gen. Genet. 204: 485-491.
D m , T., L. IELPI, S. STANFIELD, L. KASHYAP, C. DOUGLAS et al.,
1986 Rhizobium meliloti genes required for nodule develop ment are related to chromosomal virulence genes in Agrobacte- rium tumefaciens. Proc. Natl. Acad. Sci. USA 83: 4403-4407. DXAN, T., D. R. HELINSKI and G. R. D ~ A , 1990a Hypoosmotic ad-
aptation in Rhizobium meliloti requires p(1-2) glucan. J. Bacte- riol. 172: 1400-1408.
D m , T., P. NAGPUL, D. R. HELINSKI and G. R. D I A , 1990b Symbiotic pseudorevertants of Rhizobium meliloti ndu mutants. J. Bacteriol.
FIGURSKI, D. H., and D. R. HELINSKI, 1979 Replication of an origin- containing derivative plasmid of RK2 dependant on a function provided in trans. Proc. Natl. Acad. Sci. USA 76: 1648-1652. FINAN, T. M., E. HARMEG, K. LEMIEUX, K. BERGMAN, G. C. WALKER et al.,
1984 General transduction in Rhizobium meliloti. J . Bacteriol.
159: 120-124.
FINAN, T. M., A. M. HIRSCH, J. A. LEIGH, E. JOHANSEN, G. A. KULDAU et al., 1985 Symbiotic mutants of Rhizobium meliloti that un- couple plant from bacterial differentiation. Cell 4 0 869-877. FINAN, T. M., B. KUNKEL, G. F. DEVOS and E. R. SIGNER, 1986 Second
symbiotic megaplasmid in Rhizobium meliloti carrying exopc- lysaccharide and thiamine biosynthetic genes. J. Bacteriol. 167:
FINAN, T. M., I. J. ORESNIK and A. BOITACIN, 1988 Mutants of Rhizo-
3396-3403.
bium meliloti defective in succinate metabolism. J. Bacteriol. 170:
FURUICHI, T., M. INOUYE and S. INOWE, 1985 Novel one-step cloning vectorwith a transposable element: application to the Myxococcus xanthus genome. J. Bacteriol. 164: 270-275.
GLAZEBROOK, J., and G. C. WALKER, 1989 A novel exopolysaccharide can function in place of the calcofluor binding exopolysaccharide of Rhizobium meliloti. Cell 56: 661-672.
HYNES, M. F., R. SIMON, P. MULLER, K. NIEHAUS, M. LABES et al.,
1986 The two megaplasmids of Rhizobium meliloti are involved in the effective nodulation of alfalfa. Mol. Gen. Genet. 202:
IELPI, L., T. D m , G. S. DITTA, D. R. HELINSKI and S. STANFIELD, 1990 The n d v B locus of Rhizobium meliloti encodes a 319-kDa protein involved in the production of p(1-2) glucan. J. Biol. Chem. 265: 2843-2851.
JONES, J. D. G., and N. GUTTERSON, 1987 An effecient mobilizable cosmid vector, pRK7813, and its use in a method for rapid marker exchange in Pseudomonas flourescens strain HV37a. Gene 61:
299-306.
KLEIN, S., K. LOHMAN, R. CLOVER, G. C. WALKER and E. R. SIGNER, 1992 A directional, high-frequency chromosomal mobilization system for genetic mapping of Rhizobium meliloti. J. Bacteriol.
174 324-326.
LEIGH, J. A,, and C. C. LEE, 1988 Characterization of polysaccharide of Rhizobium meliloti exo mutants that form ineffective nodules. J. Bacteriol. 170: 3327-3332.
LEIGH, J. A,, E. R. SIGNER and G. C. WALKER, 1985 Exopolysaccharide deficient mutants of Rhizobium meliloti that form ineffective nod- ules, Proc. Natl. Acad. Sci. USA 82: 6231-6234.
LEROUCF,, P., P. ROCHE, C. FAUCHER, F. MAILLET, G. TRUCHET et al., 1 7 2 1409-1417.
66-72.
R. meliloti nduF Suppressor Alleles 1243
1990 Symbiotic host specificity of Rhizobium meliloti is deter- mined by a sulphated and acylated glucosamine oligo saccharide signal. Nature 344: 781-784.
LONG, S. R., 1989 Rhizobiumgenetics. Annu. Rev. Genet. 23: 483-506
LONG, S. S., S. MCCUNE and G. C. WALKER, 1988 Symbiotic loci of
Rhizobium meliloti identified by random TnphoA mutagenesis. J. Bacteriol. 170: 4257-4265.
MEADE, H. M., and E. R. SIGNER, 1977 Genetic mapping of Rhizobium meliloti. Proc. Natl. Acad. Sci. USA 74: 2076-2078.
MEADE, H., S. LONG, G. RUVKIN, S. BROWN and F. AUSUBEL, 1982 Physi- cal and genetic characterization of symbiotic and auxotrophic mutants of Rhizobium meliloti. J. Bacteriol. 149: 114-122.
MEYER, S. E., S. GRANETT, J. U . JUNG and M. R. VILLARIJO, 1990 Osmotic regulation o f p h o E porin synthesis in Escherichia coli. J. Bacteriol.
NAGPAL, P., S. P. S. KHANUJA and S. W. STANFIELD, 1992 Suppression of the n d u mutant phenotype of Rhizobium meliloti by cloned exo
genes. Mol. Microbiol. 6: 479-488.
ORESNIK, I. J., 1990 Genetic analysis of second site revertants of Jix- I14 ( n d u F ) in Rhizobium meliloti. M.Sc. Thesis, McMaster Uni- versity, Hamilton, Ontario.
PUTNOKY, P., G. PETROVICS, A. KERESZT, E. GROSSKOPF, D. C. HA et al., 1990 Rhizobium meliloti lipopolysaccharide and exopolysaccha- ride can have the same function in the plant-bacterium interac- tion. J. Bacteriol. 172: 5450-5458.
REED, J. W., and G . C. WALKER, 1991 Acidic conditions permit effec- tive nodulation of alfalfa by invasiondeficient Rhizobium meliloti exoD mutants. Genes Dev. 5 2274-2287.
SHORTRIDGE, V. D., A. LAZDUNSKI and M. L. VASIL, 1992 Osmopro- tectants and phosphate regulate expression of phospholipase C
in Pseudomonas aerugznosa. Mol. Microbiol. 6: 863-871.
SIMON, R., 1984 High frequency mobilization of gram-negative bac-
172: 5501-5502.
terial replicons by in vitro constructed Tn5-mob transposon. Mol. Gen. Genet. 196: 413-420.
SPAINK, H, P., D. M. SHEELEY, A. A. N. VAN BRUSSEL, J. GLUSHKA, W. S. YORK, T. TAK et al., 1991 A novel highly unsaturated, fatty acid moiety of lipo-oligosaccharide signal molecules determines host- specificity of Rhizobium leguminosarum. Nature 356: 125-130.
phated lipo-oligosaccharide signals of Rhizobium meliloti ellicit root nodule organogenesis in alfalfa. Nature 351: 670-673.
VAN BRUSSEL, A. A. N., R. BAKHUIZEN, P. C. VAN SPRONSEN, H. P. SPAINK, T. TAK et al., 1992 Induction of pre-infection thread structures in the legimonous host plant by mitogenic lipc-oligosaccharides of Rhizobium. Science 257: 70-72
VILWJO, M., J. L. DAVIS and S. GRANETT, 1983 Osmoregulation of alkaline phosphatase synthesis in Escherichia coli K-12. J. Bacte- rial. 156 975-978.
VINCENT, J. M., 1970 A Manual for the Practical Study oJRoot Nodule Bacteria (IBM Handbook No. 15). Blackwell Scientific Publica- tions, Oxford.
WILLIAMS, M. N. V., R. I. HOLLINGSWORTH, S. KLEIN and E. R. SIGNER,
1990 The symbiotic defect of Rhizobium meliloti exopolysaccha- ride mutant is suppressed by IpsZ', a gene involved in lipopo- lysaccharide biosynthesis. J. Bacteriol. 172: 2622-2632.
ZAHN, H., S. B. LEVERY, C. C. LEE and J. A. LEIGH, 1989 A second exopolysaccharide of Rhizobium meliloti strain SU47 that can function in root nodule invasion. Proc. Natl. Acad. Sci. USA 86: 3055-3059.
ZHAN, H., C. C. LEE and J. A. LEIGH, 1991 Induction of the second exopolysaccharide (EPSb) in Rhizobium meliloti SU47 by low phosphate concentrations. J. Bacteriol. 173: 7319-7394.
TRUCHET, G., P. ROCHE, P. LEROUGE, J. VASSE,